
Myorelaxant Effect of the Dysphania ambrosioides Essential Oil on Sus scrofa domesticus Coronary Artery and Its Toxicity in the Drosophila melanogaster Model

 , , , ,
, , , ,
Abstract
1. Introduction
2. Results
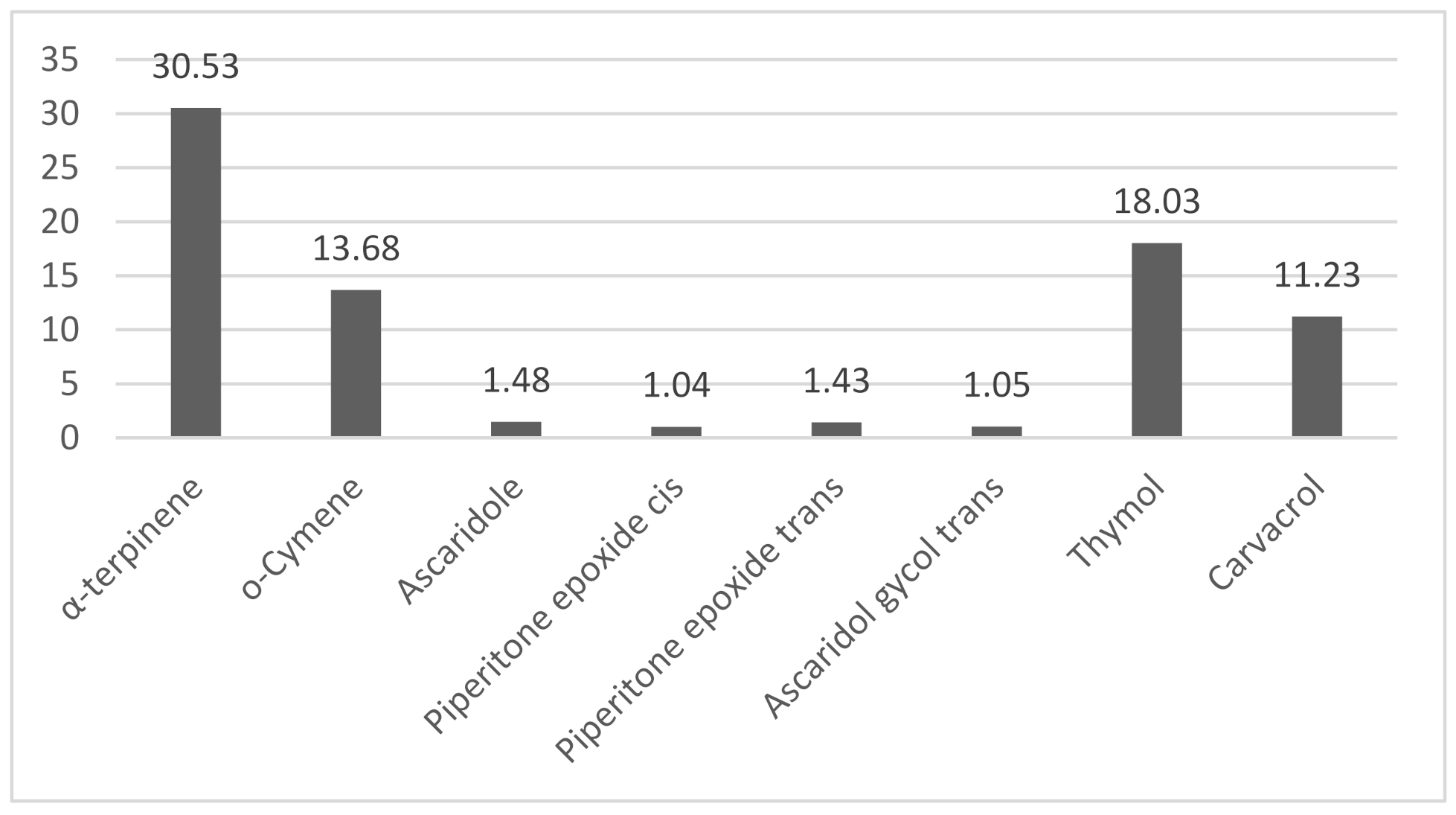
2.1. Phytochemical Analysis
2.2. Ex Vivo Smooth Muscle Contractility Assay
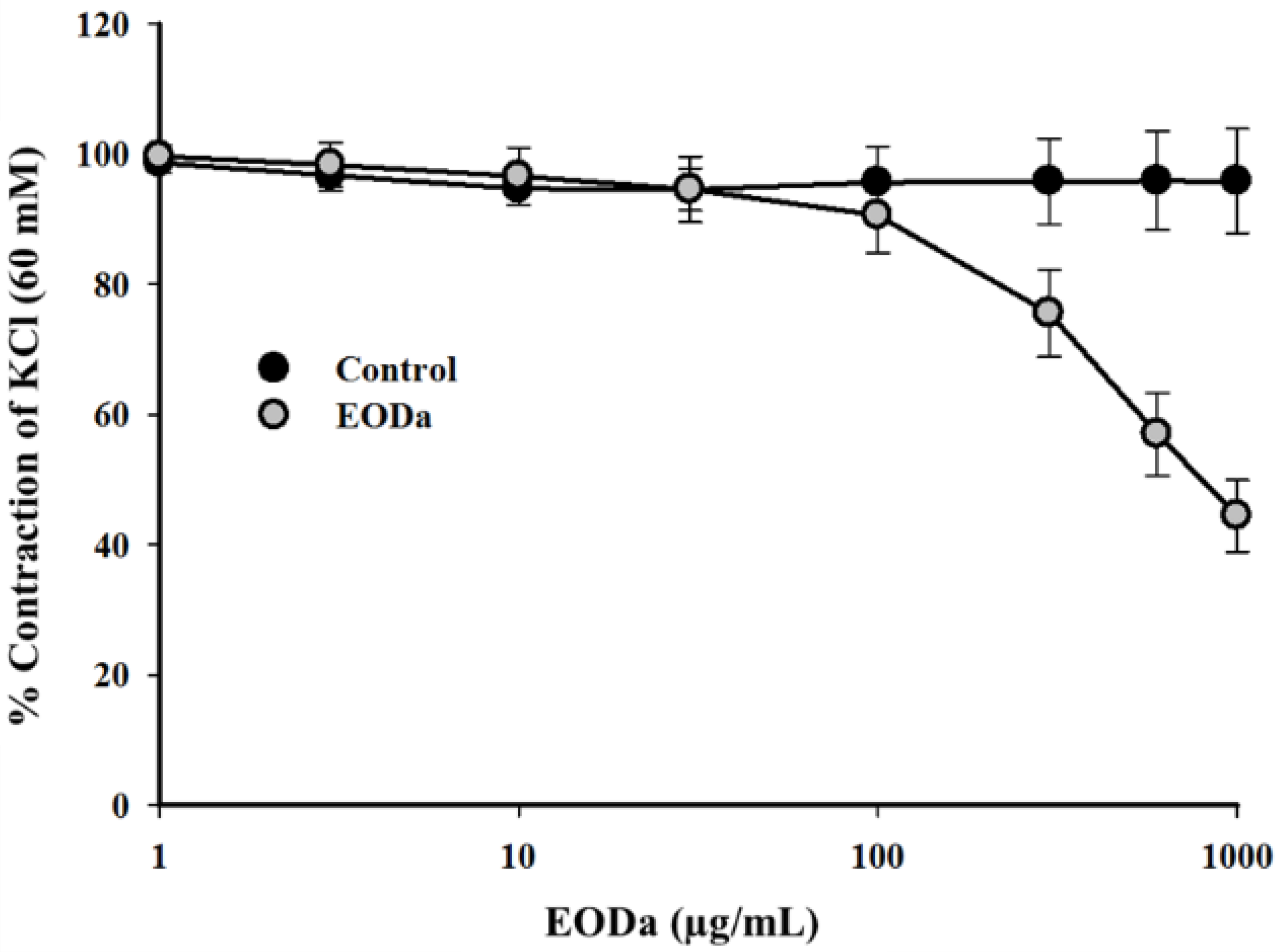
2.2.1. Effect of the Dysphania ambrosioides Essential Oil on K+-Induced Contractions Using a Sus scrofa domesticus Coronary Artery Model
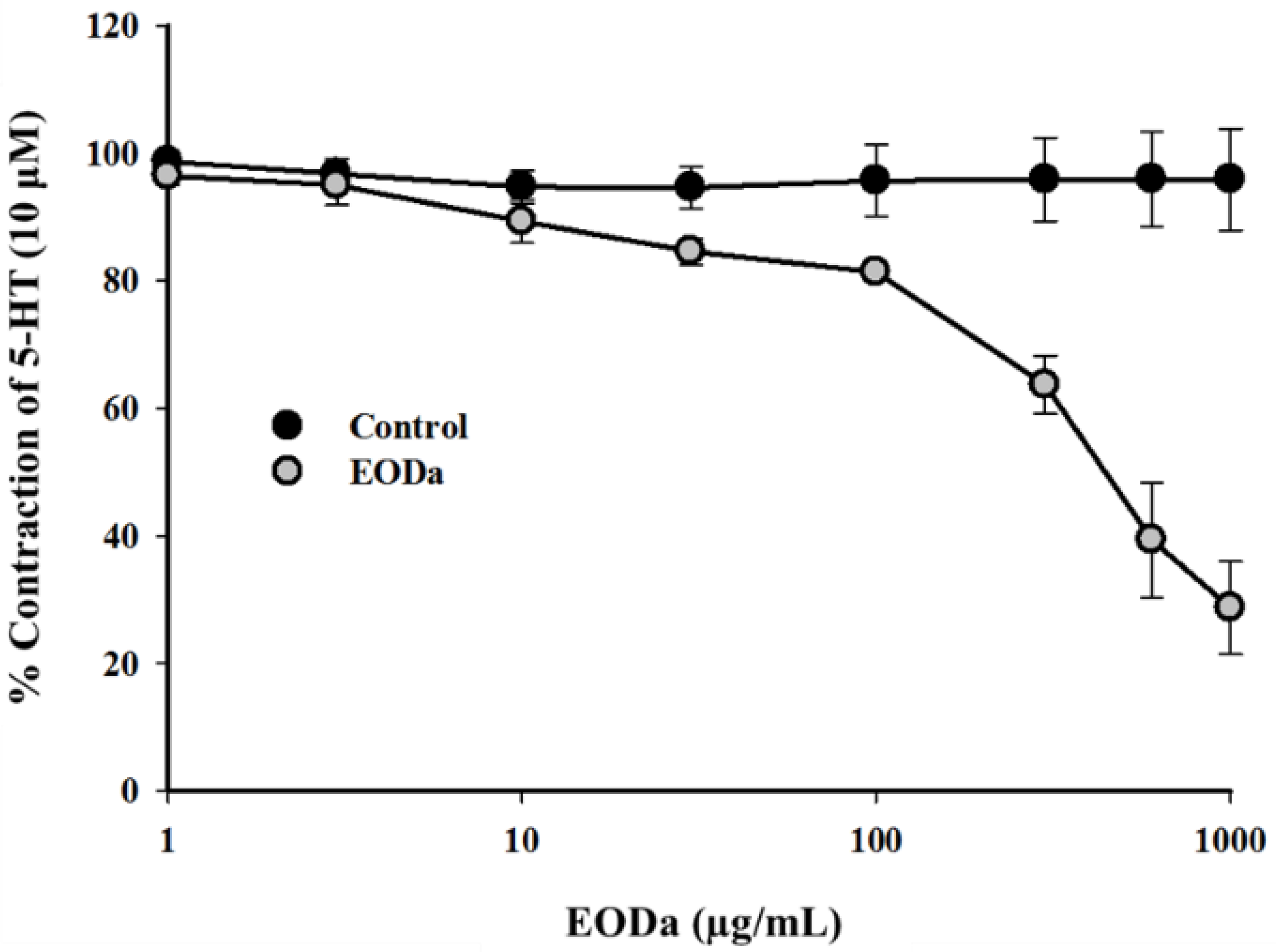
2.2.2. Effect of the Dysphania ambrosioides Essential Oil on Contractions Induced by Serotonin (5-HT) Using a Sus scrofa domesticus Coronary Artery Model
2.3. D. melanogaster-Alternative Model
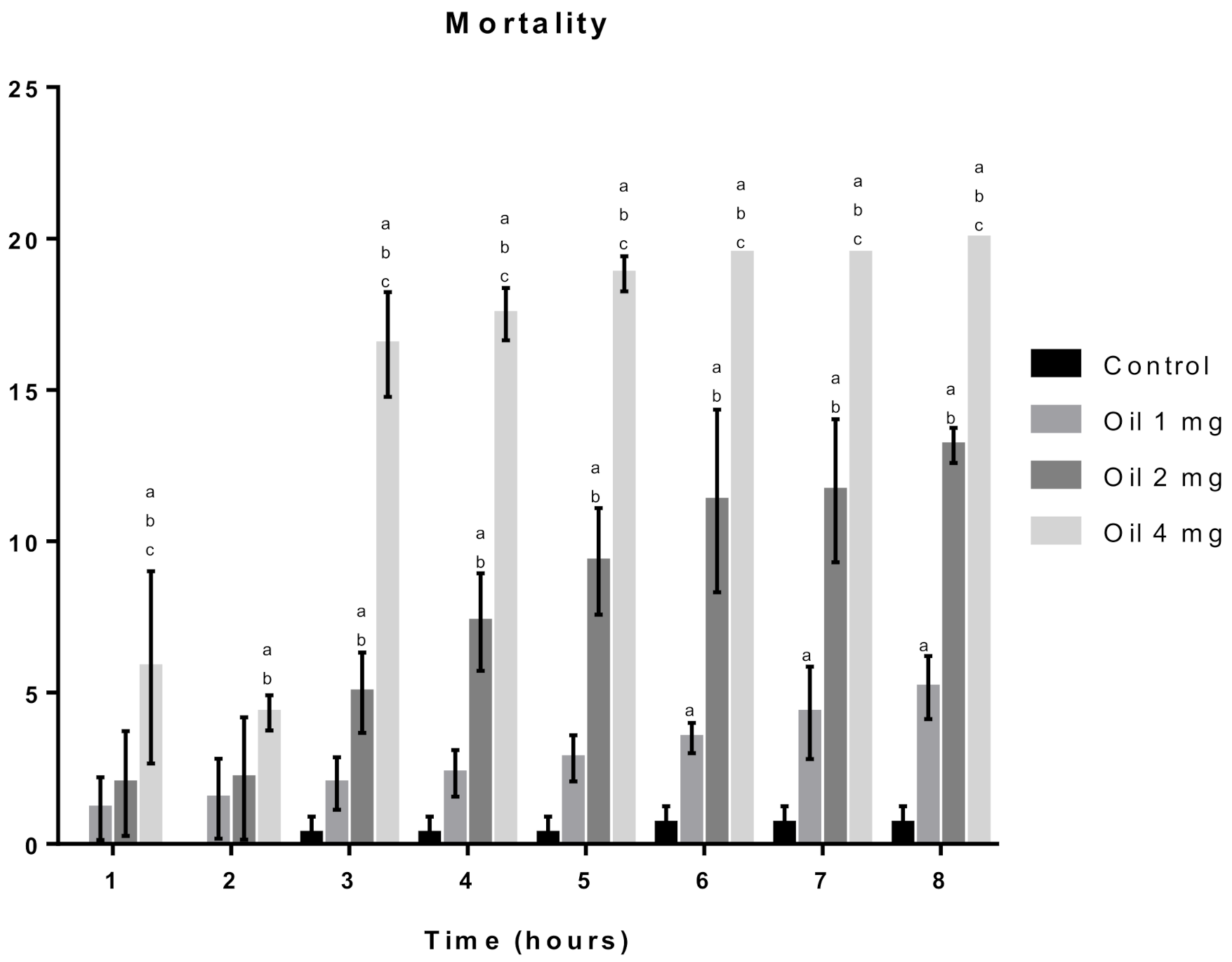
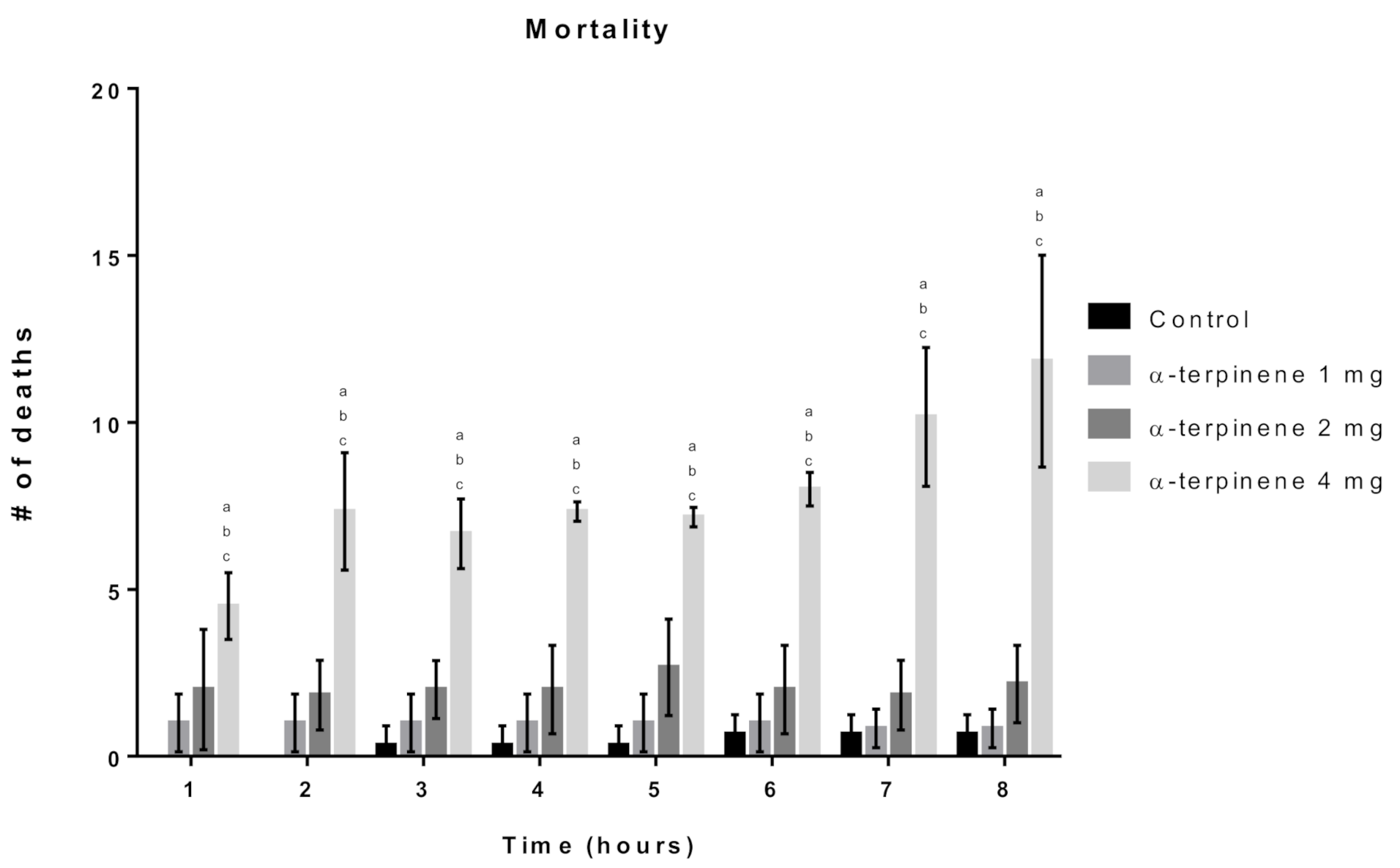
2.3.1. Toxicity Assay
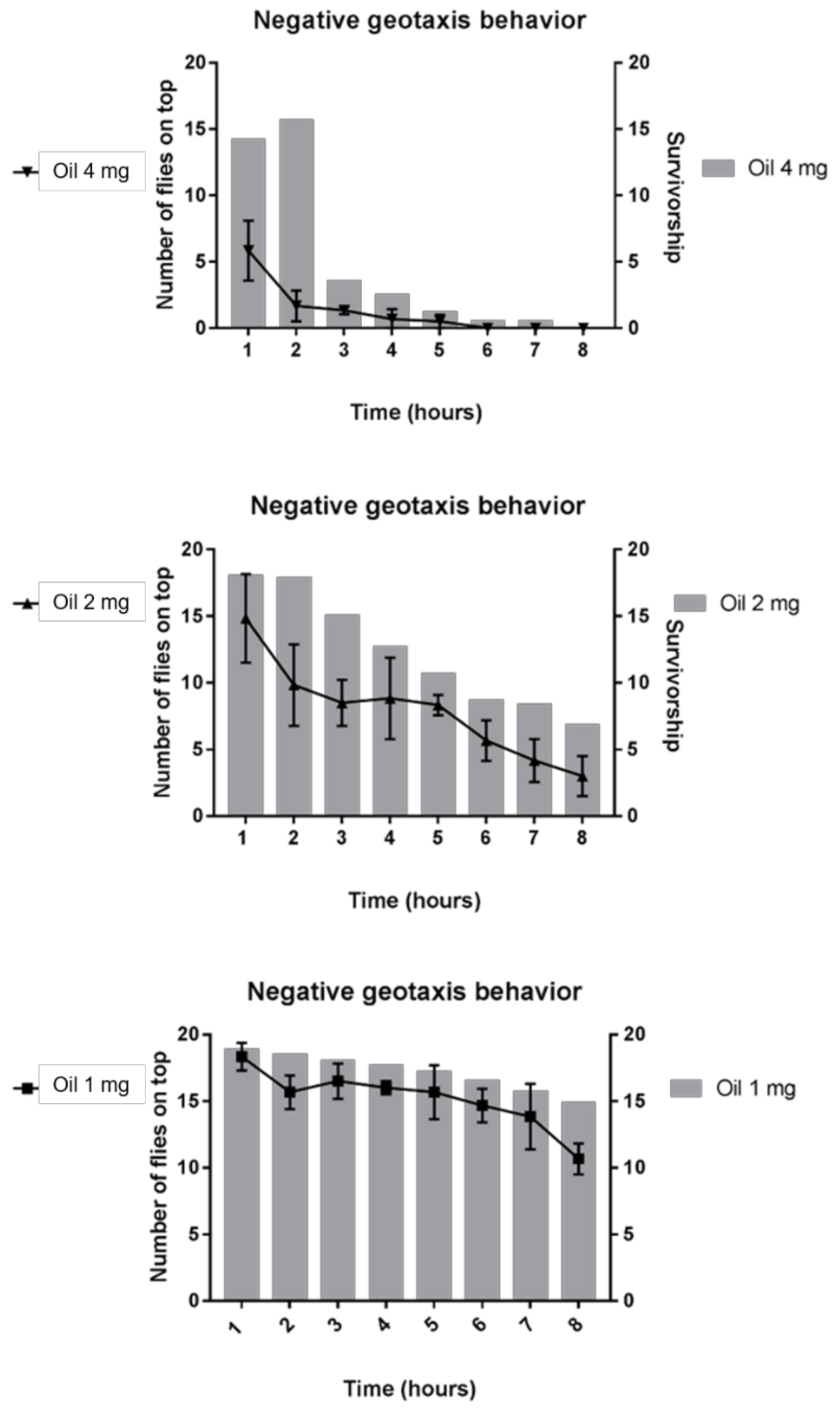
2.3.2. Geotaxis
3. Discussion
4. Materials and Methods
4.1. Plant Collection and Identification
4.2. Essential Oil Extraction
4.3. Phytochemical Analysis
4.4. Ex Vivo Assays and Pharmacological Tests
4.5. Toxicity Test with the Drosophila melanogaster Alternative Model
4.5.1. Drosophila Stock and Culture
4.5.2. D. melanogaster Assays and Essential Oil Exposure
4.5.3. Locomotor Assay
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Gruber, F.P.; Hartung, T. Alternatives to animal experimentation in basic research. ALTEX 2004, 21 (Suppl. 1), 3–31. [Google Scholar] [PubMed]
- Doke, S.K.; Dhawale, S.C. Alternatives to animal testing: A review. Saudi Pharm. J. 2015, 23, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Hamm, J.; Sullivan, K.; Clippinger, A.J.; Strickland, J.; Bell, S.; Bhhatarai, B.; Blaauboer, B.; Casey, W.; Dorman, D.; Forsby, A.; et al. Alternative approaches for identifying acute systemic toxicity: Moving from research to regulatory testing. Toxicol. Vitr. 2017, 41, 245–259. [Google Scholar] [CrossRef] [PubMed]
- Mwanauta, R.W.; Mtei, K.A.; Ndakidemi, P.A. Prospective Bioactive Compounds from Vernonia amygdalina, Lippia javanica, Dysphania ambrosioides and Tithonia diversifolia in Controlling Legume Insect Pests. Agric. Sci. 2014, 5, 1129–1139. [Google Scholar] [CrossRef][Green Version]
- Aumeeruddy-Elalfi, Z.; Mahomoodally, F. Essential oils and nanoemulsions alternative tool to biofilm eradication. In Essential Oils and Nanotechnology for Treatment of Microbial Diseases; Rai, M., Zacchino, S., Derita, M.G., Eds.; CRC Press: Boca Raton, FL, USA; CRC Press/Taylor & Francis Group: Milton Park, UK, 2017; pp. 159–174. ISBN 9781351806077. [Google Scholar]
- De Lavor, É.M.; Fernandes, A.W.C.; de Andrade Teles, R.B.; Leal, A.E.B.P.; de Oliveira Júnior, R.G.; Gama e Silva, M.; De Oliveira, A.P.; Silva, J.C.; de Moura Fontes Araújo, M.T.; Coutinho, H.D.M.; et al. Essential Oils and Their Major Compounds in the Treatment of Chronic Inflammation: A Review of Antioxidant Potential in Preclinical Studies and Molecular Mechanisms. Oxid. Med. Cell. Longev. 2018, 2018, 6468593. [Google Scholar] [CrossRef]
- Simões, R.R.; dos Santos Coelho, I.; Junqueira, S.C.; Pigatto, G.R.; Salvador, M.J.; Santos, A.R.S.; de Faria, F.M. Oral treatment with essential oil of Hyptis spicigera Lam. (Lamiaceae) reduces acute pain and inflammation in mice: Potential interactions with transient receptor potential (TRP) ion channels. J. Ethnopharmacol. 2017, 200, 8–15. [Google Scholar] [CrossRef]
- Lorenzi, H.; Matos, F.J. Plantas Medicinais No Brasil: Nativas e Exóticas, 2nd ed.; Instituto Plantarum: Nova Odessa, Brazil, 2002. [Google Scholar]
- Pavela, R.; Maggi, F.; Lupidi, G.; Mbuntcha, H.; Woguem, V.; Womeni, H.M.; Barboni, L.; Tapondjou, L.A.; Benelli, G. Clausena anisata and Dysphania ambrosioides essential oils: From ethno-medicine to modern uses as effective insecticides. Environ. Sci. Pollut. Res. 2018, 25, 10493–10503. [Google Scholar] [CrossRef]
- Zohra, T.; Ovais, M.; Khalil, A.T.; Qasim, M.; Ayaz, M.; Shinwari, Z.K. Extraction optimization, total phenolic, flavonoid contents, HPLC-DAD analysis and diverse pharmacological evaluations of Dysphania ambrosioides (L.) Mosyakin & Clemants. Nat. Prod. Res. 2019, 33, 136–142. [Google Scholar] [CrossRef]
- Araujo, I.B.; Souza, C.A.M.; De-Carvalho, R.R.; Kuriyama, S.N.; Rodrigues, R.P.; Vollmer, R.S.; Alves, E.N.; Paumgartten, F.J.R. Study of the embryofoetotoxicity of α-terpinene in the rat. Food Chem. Toxicol. 1996, 34, 477–482. [Google Scholar] [CrossRef]
- Baldissera, M.D.; Souza, C.F.; Dolci, G.S.; Grando, T.H.; Sagrillo, M.R.; Vaucher, R.A.; da Luz, S.C.A.; Silveira, S.O.; Duarte, M.M.M.F.; Duarte, T.; et al. Monoterpene alpha-terpinene induced hepatic oxidative, cytotoxic and genotoxic damage is associated to caspase activation in rats. J. Appl. Biomed 2017, 15, 187–195. [Google Scholar] [CrossRef]
- Baldissera, M.D.; Souza, C.F.; Grando, T.H.; Sagrillo, M.R.; De Brum, G.F.; Nascimento, K.; Peres, D.S.; Maciel, M.F.; Silveira, S.O.; Da Luz, S.C.A.; et al. Memory deficit, toxic effects and activity of Na+, K+-ATPase and NTPDase in brain of Wistar rats submitted to orally treatment with alpha-terpinene. Environ. Toxicol. Pharmacol. 2016, 46, 1–8. [Google Scholar] [CrossRef]
- Rodrigues, M.S.A.C.; Silva, A.C.; Aguas, A.P.; Grande, N.R. The coronary circulation of the pig heart: Comparison with the human heart. Eur. J. Anat. 2021, 9, 67–87. [Google Scholar]
- Goodwill, A.G.; Noblet, J.N.; Sassoon, D.; Fu, L.; Kassab, G.S.; Schepers, L.; Herring, B.P.; Rottgen, T.S.; Tune, J.D.; Dick, G.M. Critical contribution of KV1 channels to the regulation of coronary blood flow. Basic Res. Cardiol. 2016, 111, 1–13. [Google Scholar] [CrossRef]
- Grover, A.K. Sodium–Calcium Exchanger in Pig Coronary Artery. In Advances in Pharmacology; Academic Press Inc.: Cambridge, MA, USA, 2017; Volume 78, pp. 145–170. [Google Scholar]
- Lawandy, I.; Liu, Y.; Shi, G.; Zhang, Z.; Scrimgeour, L.A.; Pavlov, V.; Jaworski, R.; Sellke, F.W.; Feng, J. Increased coronary arteriolar contraction to serotonin in juvenile pigs with metabolic syndrome. Mol. Cell. Biochem. 2019, 461, 57–64. [Google Scholar] [CrossRef]
- Bell, D.R.; Gochenaur, K. Direct vasoactive and vasoprotective properties of anthocyanin-rich extracts. J. Appl. Physiol. 2006, 100, 1164–1170. [Google Scholar] [CrossRef]
- Auger, C.; Pollet, B.; Arnold, C.; Marx, C.; Schini-Kerth, V.B. Great heterogeneity of commercial fruit juices to induce endothelium-dependent relaxations in Isolated porcine coronary Arteries: Role of the phenolic content and composition. J. Med. Food 2015, 18, 128–136. [Google Scholar] [CrossRef]
- Elle, R.E.; Defraigne, J.-O.; Schini-kerth, V.; Pollet, B.; Augier, C.; Kevers, C.; Tabart, J.; Pincemail, J. Ex-vivo vasorelaxation induced by polyphenol-rich extracts from plant, fruits and berries on porcine coronary artery rings. Free Radic. Biol. Med. 2018, 120, S80. [Google Scholar] [CrossRef]
- Tabart, J.; Auger, C.; Kevers, C.; Dommes, J.; Pollet, B.; Defraigne, J.O.; Schini-Kerth, V.B.; Pincemail, J. The potency of commercial blackcurrant juices to induce relaxation in porcine coronary artery rings is not correlated to their antioxidant capacity but to their anthocyanin content. Nutrition 2018, 51–52, 53–59. [Google Scholar] [CrossRef]
- Wu, H.C.; Horng, C.T.; Tsai, S.C.; Lee, Y.L.; Hsu, S.C.; Tsai, Y.J.; Tsai, F.J.; Chiang, J.H.; Kuo, D.H.; Yang, J.S. Relaxant and vasoprotective effects of ginger extracts on porcine coronary arteries. Int. J. Mol. Med. 2018, 41, 2420–2428. [Google Scholar] [CrossRef]
- Pereira-de-Morais, L.; de Alencar Silva, A.; da Silva, R.E.R.; Navarro, D.M.D.A.F.; Coutinho, H.D.M.; de Menezes, I.R.A.; Kerntopf, M.R.; da Cunha, F.A.B.; Leal-Cardoso, J.H.; Barbosa, R. Myorelaxant action of the Dysphania ambrosioides (L.) Mosyakin & Clemants essential oil and its major constituent α-terpinene in isolated rat trachea. Food Chem. 2020, 325, 126923. [Google Scholar] [CrossRef]
- Boutkhil, S.; El Idrissi, M.; Amechrouq, A.; Chbicheb, A.; Chakir, S.; El Badaoui, K. Chemical composition and antimicrobial activity of crude, aqueous, ethanol extracts and essential oils of Dysphania ambrosioides (L.) Mosyakin & Clemants. Acta Bot. Gall. 2009, 156, 201–209. [Google Scholar] [CrossRef]
- Chu, S.S.; Feng Hu, J.; Liu, Z.L. Composition of essential oil of Chinese Chenopodium ambrosioides and insecticidal activity against maize weevil, Sitophilus zeamais. Pest Manag. Sci. 2011, 67, 714–718. [Google Scholar] [CrossRef]
- Johnson, M.A.; Croteau, R. Biosynthesis of ascaridole: Iodide peroxidase-catalyzed synthesis of a monoterpene endoperoxide in soluble extracts of Chenopodium ambrosioides fruit. Arch. Biochem. Biophys. 1984, 235, 254–266. [Google Scholar] [CrossRef]
- Rudbäck, J.; Bergström, M.A.; Börje, A.; Nilsson, U.; Karlberg, A.-T. α-Terpinene, an antioxidant in tea tree oil, autoxidizes rapidly to skin allergens on air exposure. Chem. Res. Toxicol. 2012, 25, 713–721. [Google Scholar] [CrossRef]
- Bolton, T.B.; Prestwich, S.A.; Zholos, A.V.; Gordienko, D. V Excitation-contraction coupling in gastrointestinal and other smooth muscles. Annu. Rev. Physiol. 1999, 61, 85–115. [Google Scholar] [CrossRef]
- Catterall, W.A. Voltage-gated calcium channels. Cold Spring Harb. Perspect. Biol. 2011, 3, a003947. [Google Scholar] [CrossRef]
- Clelland, L.J.; Browne, B.M.; Alvarez, S.M.; Miner, A.S.; Ratz, P.H. Rho-kinase inhibition attenuates calcium-induced contraction in β-escin but not Triton X-100 permeabilized rabbit femoral artery. J. Muscle Res. Cell Motil. 2011, 32, 77–88. [Google Scholar] [CrossRef]
- Menezes, P.M.N.; Brito, M.C.; de Paiva, G.O.; dos Santos, C.O.; de Oliveira, L.M.; de Araújo Ribeiro, L.A.; de Lima, J.T.; Lucchese, A.M.; Silva, F.S. Relaxant effect of Lippia origanoides essential oil in guinea-pig trachea smooth muscle involves potassium channels and soluble guanylyl cyclase. J. Ethnopharmacol. 2018, 220, 16–25. [Google Scholar] [CrossRef]
- Hille, B. Ion Channels of Excitable Membranes, 3rd ed.; Sinauer: Sunderland, MA, USA, 2001; Volume 507. [Google Scholar]
- Jackson, W.F.; Boerman, E.M. Voltage-gated Ca2+ channel activity modulates smooth muscle cell calcium waves in hamster cremaster arterioles. Am. J. Physiol. Circ. Physiol. 2018, 315, H871–H878. [Google Scholar] [CrossRef]
- Morgado, M.; Cairrão, E.; Santos-Silva, A.J.; Verde, I. Cyclic nucleotide-dependent relaxation pathways in vascular smooth muscle. Cell. Mol. Life Sci. 2012, 69, 247–266. [Google Scholar] [CrossRef]
- Pelaia, G.; Renda, T.; Gallelli, L.; Vatrella, A.; Busceti, M.T.; Agati, S.; Caputi, M.; Cazzola, M.; Maselli, R.; Marsico, S.A. Molecular mechanisms underlying airway smooth muscle contraction and proliferation: Implications for asthma. Respir. Med. 2008, 102, 1173–1181. [Google Scholar] [CrossRef] [PubMed]
- Sadraei, H.; Ghannadi, A.; Malekshahi, K. Relaxant effect of essential oil of Melissa officinalis and citral on rat ileum contractions. Fitoterapia 2003, 74, 445–452. [Google Scholar] [CrossRef]
- Ribeiro, T.F.; Da Silva, R.E.R.; de Morais, L.P.; Carvalho, P.M.M.; Macêdo, C.A.F.; Silva, A.A.; Menezes, I.R.A.; Barbosa, R.; Kerntopf, M.R. Effect of the Lippia alba (Mill.) N.E. Brown essential oil and its main constituents, citral and limonene, on the tracheal smooth muscle of rats. Biotechnol. Rep. 2017, 17, 31–34. [Google Scholar] [CrossRef]
- Jarvis, G.E.; Barbosa, R.; Thompson, A.J. Noncompetitive inhibition of 5-HT3 receptors by citral, linalool, and eucalyptol revealed by nonlinear mixed-effects modeling. J. Pharmacol. Exp. Ther. 2016, 356, 549–562. [Google Scholar] [CrossRef]
- Assaidi, A.; Legssyer, A.; Berrichi, A.; Aziz, M.; Mekhfi, H.; Bnouham, M.; Ziyyat, A. Hypotensive property of Chenopodium ambrosioides in anesthetized normotensive rats. J. Complement. Integr. Med. 2014, 11, 1–7. [Google Scholar] [CrossRef]
- Pinho, A.I.; Wallau, G.L.; Nunes, M.E.M.; Leite, N.F.; Tintino, S.R.; da Cruz, L.C.; da Cunha, F.A.B.; da Costa, J.G.M.; Douglas Melo Coutinho, H.; Posser, T. Fumigant activity of the Psidium guajava var. pomifera (Myrtaceae) essential oil in Drosophila melanogaster by means of oxidative stress. Oxid. Med. Cell. Longev. 2014, 2014, 696785. [Google Scholar] [CrossRef]
- Soares, M.H.; Dias, H.J.; Vieira, T.M.; de Souza, M.G.M.; Cruz, A.F.F.; Badoco, F.R.; Nicolella, H.D.; Cunha, W.R.; Groppo, M.; Martins, C.H.G.; et al. Chemical Composition, Antibacterial, Schistosomicidal, and Cytotoxic Activities of the Essential Oil of Dysphania ambrosioides (L.) Mosyakin & Clemants (Chenopodiaceae). Chem. Biodivers. 2017, 14, e1700149. [Google Scholar] [CrossRef]
- Arena, J.S.; Omarini, A.B.; Zunino, M.P.; Peschiutta, M.L.; Defagó, M.T.; Zygadlo, J.A. Essential oils from Dysphania ambrosioides and Tagetes minuta enhance the toxicity of a conventional insecticide against Alphitobius diaperinus. Ind. Crops Prod. 2018, 122, 190–194. [Google Scholar] [CrossRef]
- Da Cunha, F.A.B.; Wallau, G.L.; Pinho, A.I.; Nunes, M.E.M.; Leite, N.F.; Tintino, S.R.; Da Costa, G.M.; Athayde, M.L.; Boligon, A.A.; Coutinho, H.D.M.; et al. Eugenia uniflora leaves essential oil induces toxicity in Drosophila melanogaster: Involvement of oxidative stress mechanisms. Toxicol. Res. 2015, 4, 634–644. [Google Scholar] [CrossRef]
- Adedara, I.A.; Abolaji, A.O.; Rocha, J.B.T.; Farombi, E.O. Diphenyl Diselenide Protects Against Mortality, Locomotor Deficits and Oxidative Stress in Drosophila melanogaster Model of Manganese-Induced Neurotoxicity. Neurochem. Res. 2016, 41, 1430–1438. [Google Scholar] [CrossRef]
- Jiang, Z.; Akhtar, Y.; Bradbury, R.; Zhang, X.; Isman, M.B. Comparative toxicity of essential oils of Litsea pungens and Litsea cubeba and blends of their major constituents against the cabbage looper, Trichoplusia ni. J. Agric. Food Chem. 2009, 57, 4833–4837. [Google Scholar] [CrossRef]
- Paes, J.P.P.; Rondelli, V.M.; Costa, A.V.; Vianna, U.R.; De Queiroz, V.T. Chemical characterization and effect of the essential oil from santa-maria herb on the strawberry mite. Rev. Bras. Frutic. 2015, 37, 346–354. [Google Scholar] [CrossRef][Green Version]
- Zhang, Z.; Yang, T.; Zhang, Y.; Wang, L.; Xie, Y. Fumigant toxicity of monoterpenes against fruitfly, Drosophila melanogaster. Ind. Crops Prod. 2016, 81, 147–151. [Google Scholar] [CrossRef]
- Omolo, M.O.; Okinyo, D.; Ndiege, I.O.; Lwande, W.; Hassanali, A. Fumigant toxicity of the essential oils of some African plants against Anopheles gambiae sensu stricto. Phytomedicine 2005, 12, 241–246. [Google Scholar] [CrossRef]
- Lacerda-Neto, L.J.; Barbosa, A.G.; Quintans-Junior, L.J.; Coutinho, H.D.; da Cunha, F.A. The complex pharmacology of natural products. Future Med. Chem. 2019, 11, 797–799. [Google Scholar] [CrossRef]
- Abolaji, A.O.; Kamdem, J.P.; Lugokenski, T.H.; Farombi, E.O.; Souza, D.O.; da Silva Loreto, É.L.; Rocha, J.B.T. Ovotoxicants 4-vinylcyclohexene 1, 2-monoepoxide and 4-vinylcyclohexene diepoxide disrupt redox status and modify different electrophile sensitive target enzymes and genes in Drosophila melanogaster. Redox Biol. 2015, 5, 328–339. [Google Scholar] [CrossRef]
- Medeiros, E.; Prigol, M.; Alves, D.; do Sacramento, M.; Musachio, E.A.S.; de Freitas Couto, S. Assessment of toxicity of compound 4-phenylselenyl-7-chlorochinoline and its changes in the fruit fly model Drosophila melanogaster. An. Salão Int. Ensino Pesqui. Extensão 2018, 9, 56–56. [Google Scholar]
- Brunner, F.; Kühberger, E.; Groschner, K.; Poch, G.; Kukovetz, W.R. Characterization of muscarinic receptors mediating endothelium-dependent relaxation of bovine coronary artery. Eur. J. Pharmacol. 1991, 200, 25–33. [Google Scholar] [CrossRef]
- Obi, T.; Kabeyama, A.; Nishio, A. Characterization of muscarinic receptor subtype mediating contraction and relaxation in equine coronary artery in vitro. J. Vet. Pharmacol. Ther. 1994, 17, 226–231. [Google Scholar] [CrossRef]
- Bruder-Nascimento, T.; Campos, D.H.S.; Cicogna, A.C. Chronic stress improves NO-and Ca2+ flux-dependent vascular function: A pharmacological study. Arq. Bras. Cardiol. 2015, 104, 226–233. [Google Scholar] [CrossRef]
- Storch, A.S.; Mattos, J.D.D.; Alves, R.; Galdino, I.D.S.; Rocha, H.N.M. Methods of Endothelial Function Assessment: Description and Applications. Int. J. Cardiovasc. Sci. 2017, 30. [Google Scholar] [CrossRef]
- Hanna, M.E.; Bednářová, A.; Rakshit, K.; Chaudhuri, A.; O’Donnell, J.M.; Krishnan, N. Perturbations in dopamine synthesis lead to discrete physiological effects and impact oxidative stress response in Drosophila. J. Insect Physiol. 2015, 73, 11–19. [Google Scholar] [CrossRef] [PubMed]
- De-Oliveira, A.C.A.X.; Ribeiro-Pinto, L.F.; Paumgartten, F.J.R. In vitro inhibition of CYP2B1 monooxygenase by β-myrcene and other monoterpenoid compounds. Toxicol. Lett. 1997, 92, 39–46. [Google Scholar] [CrossRef]
- Sanchez-Hernandez, J.C.; Sanchez, B.M. Lizard cholinesterases as biomarkers of pesticide exposure: Enzymological characterization. Environ. Toxicol. Chem. 2002, 21, 2319–2325. [Google Scholar] [CrossRef]
- Kavitha, P.; Rao, J.V. Oxidative stress and locomotor behaviour response as biomarkers for assessing recovery status of mosquito fish, Gambusia affinis after lethal effect of an organophosphate pesticide, monocrotophos. Pestic. Biochem. Physiol. 2007, 87, 182–188. [Google Scholar] [CrossRef]
- Sharma, A.; Mishra, M.; Shukla, A.K.; Kumar, R.; Abdin, M.Z.; Chowdhuri, D.K. Organochlorine pesticide, endosulfan induced cellular and organismal response in Drosophila melanogaster. J. Hazard. Mater. 2012, 221, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Ramin, M.; Azizi, P.; Motamedi, F.; Khodagholi, F.; Haghparast, A.; Fatahi, Z.; Alamdary, S.Z. Inhibition of JNK phosphorylation reverses memory deficit induced by β-amyloid (1–42) associated with decrease of apoptotic factors. Behav. Brain Res. 2014, 266, 222. [Google Scholar] [CrossRef]
- Wei, H.; Liu, J.; Li, B.; Zhan, Z.; Chen, Y.; Tian, H.; Lin, S.; Gu, X. The toxicity and physiological effect of essential oil from Chenopodium ambrosioides against the diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae). Crop Prot. 2015, 76, 68–74. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | LC50 (mg L−1 Air) | 95% Fiducial Limits |
|---|---|---|
| EODa | 1.806 | 1.588–2.296 |
| α-Terpinene | 2.282 | 1.933–2.671 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Lacerda Neto, L.J.; Ramos, A.G.B.; da Silva, R.E.R.; Pereira-de-Morais, L.; Silva, F.M.; da Costa, R.H.S.; Rodrigues Dantas, L.B.; da Costa, J.G.M.; Coutinho, H.D.M.; Kowalska, G.; et al. Myorelaxant Effect of the Dysphania ambrosioides Essential Oil on Sus scrofa domesticus Coronary Artery and Its Toxicity in the Drosophila melanogaster Model. Molecules 2021, 26, 2041. https://doi.org/10.3390/molecules26072041
de Lacerda Neto LJ, Ramos AGB, da Silva RER, Pereira-de-Morais L, Silva FM, da Costa RHS, Rodrigues Dantas LB, da Costa JGM, Coutinho HDM, Kowalska G, et al. Myorelaxant Effect of the Dysphania ambrosioides Essential Oil on Sus scrofa domesticus Coronary Artery and Its Toxicity in the Drosophila melanogaster Model. Molecules. 2021; 26(7):2041. https://doi.org/10.3390/molecules26072041
Chicago/Turabian Stylede Lacerda Neto, Luiz Jardelino, Andreza Guedes Barbosa Ramos, Renata Evaristo Rodrigues da Silva, Luís Pereira-de-Morais, Fernanda Maria Silva, Roger Henrique Sousa da Costa, Lindaiane Bezerra Rodrigues Dantas, José Galberto Martins da Costa, Henrique Douglas Melo Coutinho, Grażyna Kowalska, and et al. 2021. "Myorelaxant Effect of the Dysphania ambrosioides Essential Oil on Sus scrofa domesticus Coronary Artery and Its Toxicity in the Drosophila melanogaster Model" Molecules 26, no. 7: 2041. https://doi.org/10.3390/molecules26072041
APA Stylede Lacerda Neto, L. J., Ramos, A. G. B., da Silva, R. E. R., Pereira-de-Morais, L., Silva, F. M., da Costa, R. H. S., Rodrigues Dantas, L. B., da Costa, J. G. M., Coutinho, H. D. M., Kowalska, G., Hawlena, J., Kowalski, R., Barbosa, R., & da Cunha, F. A. B. (2021). Myorelaxant Effect of the Dysphania ambrosioides Essential Oil on Sus scrofa domesticus Coronary Artery and Its Toxicity in the Drosophila melanogaster Model. Molecules, 26(7), 2041. https://doi.org/10.3390/molecules26072041