
The Effects of Andrographolide on the Enhancement of Chondrogenesis and Osteogenesis in Human Suprapatellar Fat Pad Derived Mesenchymal Stem Cells

 ,
,  , , and
, , and 
Abstract
1. Introduction
2. Results
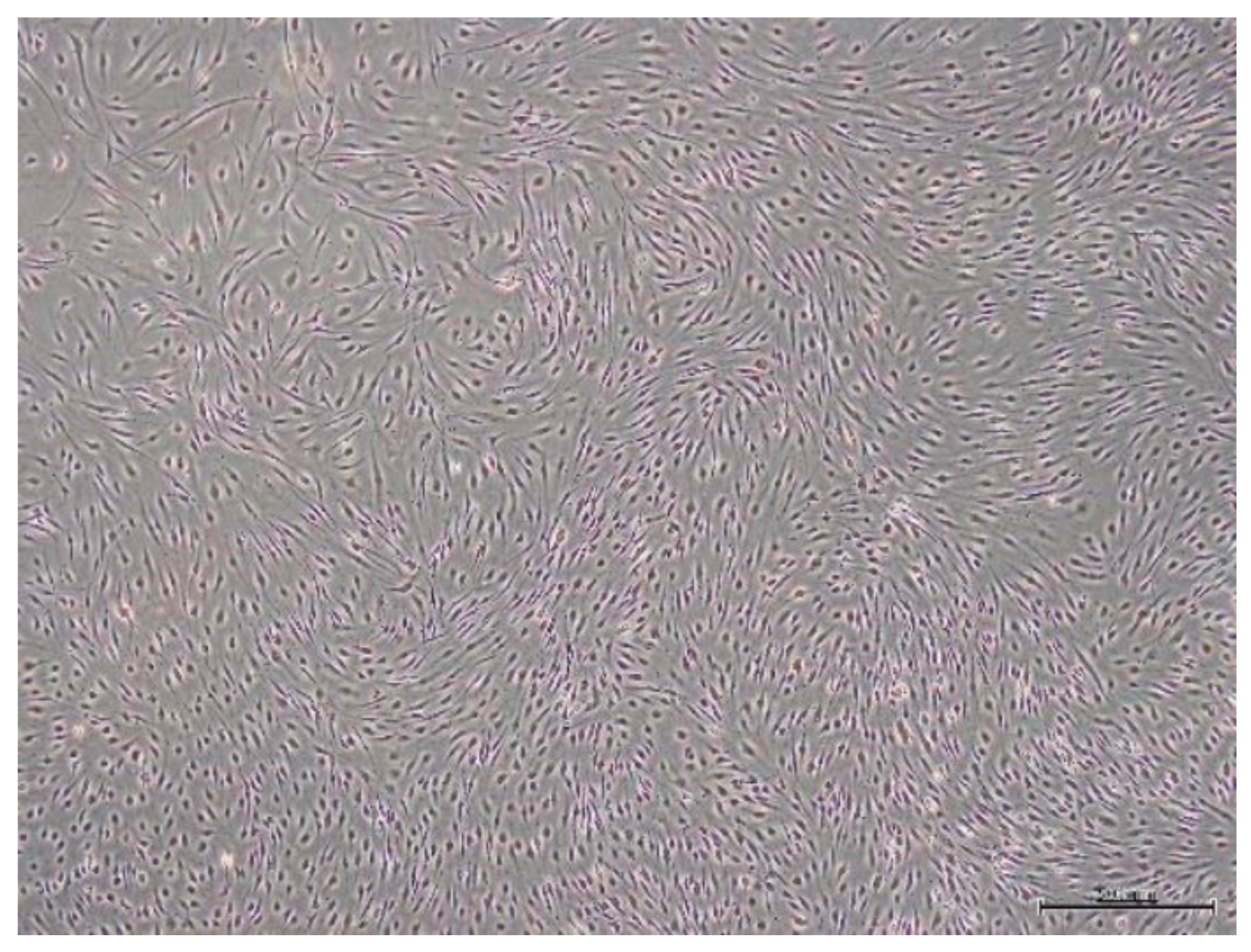
2.1. Isolation and Culture Expansion of SPFP-MSCs
2.2. Cell Surface Marker Profile
2.3. Effect of AG on the Cell Viability of Differentiating SPFP-MSCs
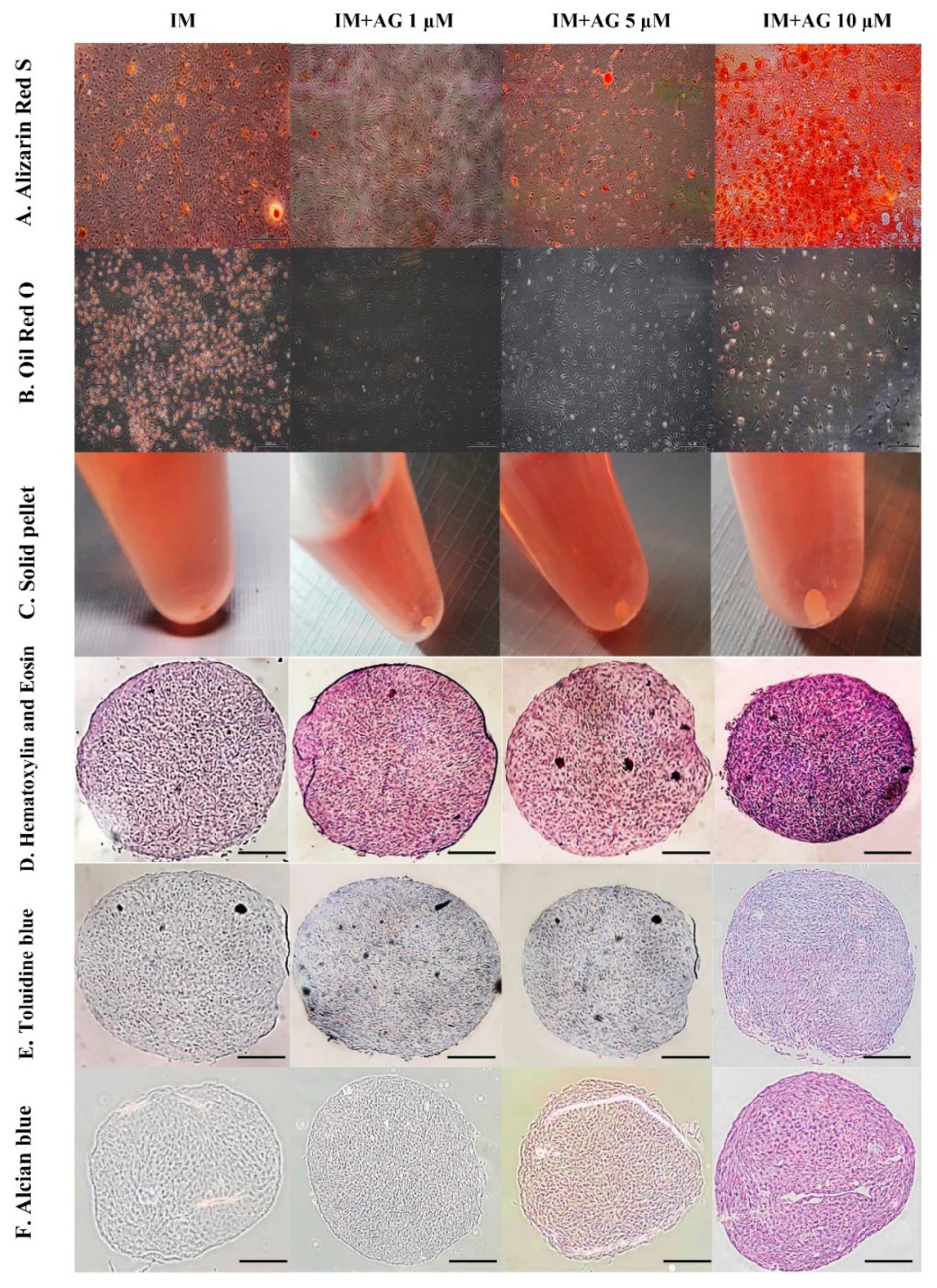
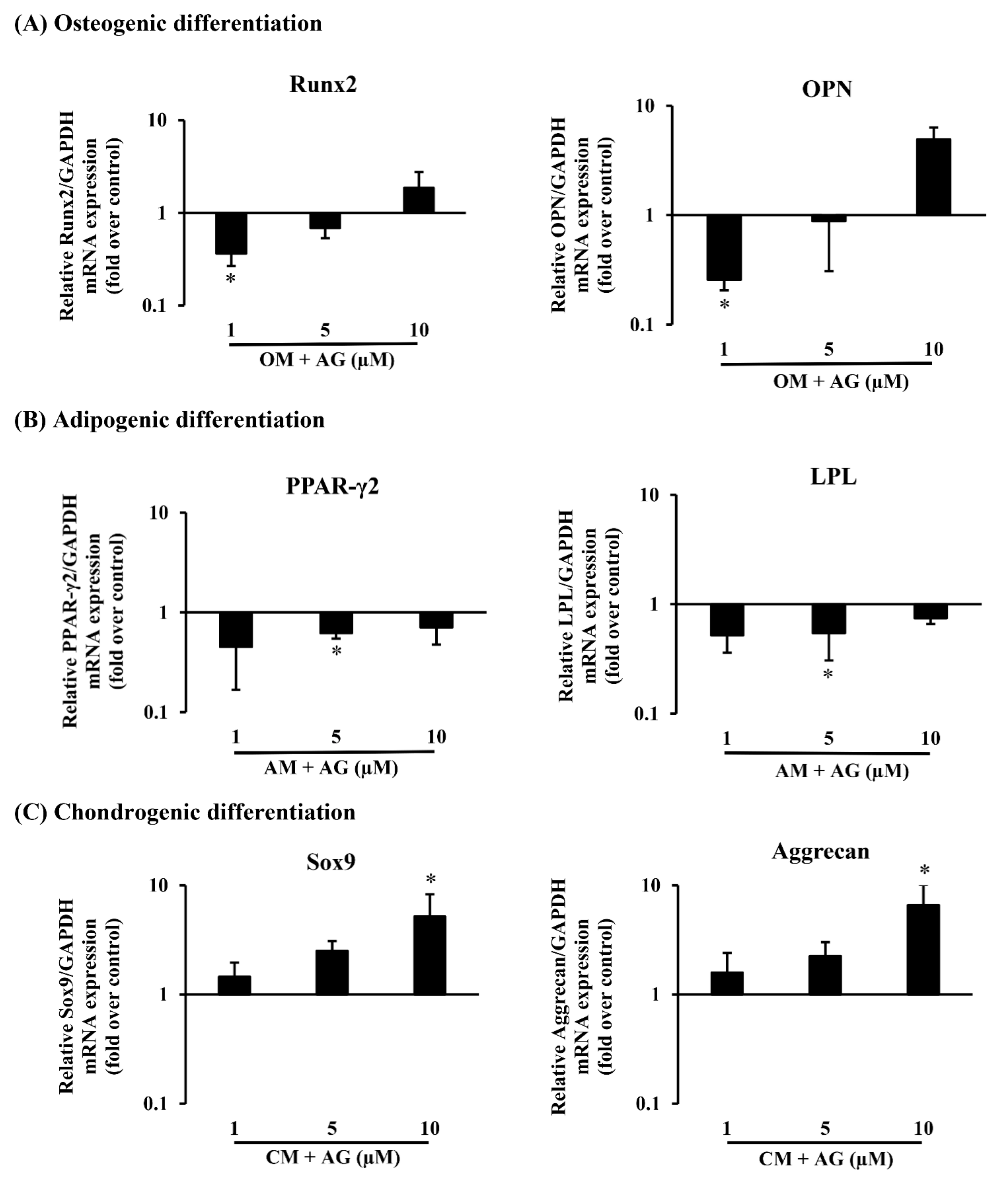
2.4. In Vitro Tri-Lineage Differentiation and Histological Analysis
3. Discussion
4. Materials and Methods
4.1. MSCs Isolation, Expansion and Culture
4.2. Flow Cytometric Analysis
4.3. Cell Viability Assay
4.4. In Vitro Tri-Lineage Differentiation and Histological Analysis
4.5. RNA Isolation and Real-Time PCR Analysis
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Lawrence, R.C.; Felson, D.T.; Helmick, C.G.; Arnold, L.M.; Choi, H.; Deyo, R.A.; Gabriel, S.; Hirsch, R.; Hochberg, M.C.; Hunder, G.G.; et al. Estimates of the prevalence of arthritis and other rheumatic conditions in the United States. Part II. Arthritis Rheum. 2008, 58, 26–35. [Google Scholar] [CrossRef]
- Kean, W.F.; Kean, R.; Buchanan, W.W. Osteoarthritis: Symptoms, signs and source of pain. Inflammopharmacology 2004, 12, 3–31. [Google Scholar] [CrossRef]
- Gerwin, N.; Hops, C.; Lucke, A. Intraarticular drug delivery in osteoarthritis. Adv. Drug Deliv. Rev. 2006, 58, 226–242. [Google Scholar] [CrossRef]
- Buckland-Wright, C. Subchondral bone changes in hand and knee osteoarthritis detected by radiography. Osteoarthr. Cartil. 2004, 12 (Suppl. A), S10–S19. [Google Scholar] [CrossRef] [PubMed]
- Haringman, J.J.; Ludikhuize, J.; Tak, P.P. Chemokines in joint disease: The key to inflammation? Ann. Rheum. Dis. 2004, 63, 1186–1194. [Google Scholar] [CrossRef]
- Steinberg, J.; Zeggini, E. Functional genomics in osteoarthritis: Past, present, and future. J. Orthop. Res. 2016, 34, 1105–1110. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Chubinskaya, S.; Rueger, D.C.; Bau, B.; Haag, J.; Aigner, T. Regulation of anabolic and catabolic gene expression in normal and osteoarthritic adult human articular chondrocytes by osteogenic protein-1. Clin. Exp. Rheumatol. 2004, 22, 103–106. [Google Scholar] [PubMed]
- Lajeunesse, D. The role of bone in the treatment of osteoarthritis11Supported by Procter & Gamble Pharmaceuticals, Mason, OH. Osteoarthr. Cartil. 2004, 12, 34–38. [Google Scholar] [CrossRef]
- Foster, M.; Hunter, D.; Samman, S. Evaluation of the Nutritional and Metabolic Effects of Aloe vera. In Herbal Medicine: Biomolecular and Clinical Aspects, 2nd ed.; Benzie, I.F.F., Wachtel-Galor, S., Eds.; CRC Press: Boca Raton, FL, USA; Taylor & Francis: Boca Raton, FL, USA, 2011; ISBN 978-1-4398-0713-2. [Google Scholar]
- Kabir, M.H.; Hasan, N.; Rahman, M.M.; Rahman, M.A.; Khan, J.A.; Hoque, N.T.; Bhuiyan, M.R.Q.; Mou, S.M.; Jahan, R.; Rahmatullah, M. A survey of medicinal plants used by the Deb barma clan of the Tripura tribe of Moulvibazar district, Bangladesh. J. Ethnobiol. Ethnomed. 2014, 10, 19. [Google Scholar] [CrossRef] [PubMed]
- Akbar, S. Andrographis paniculata: A review of pharmacological activities and clinical effects. Altern. Med. Rev. 2011, 16, 66–77. [Google Scholar]
- Jarukamjorn, K.; Nemoto, N. Pharmacological Aspects of Andrographis paniculata on Health and Its Major Diterpenoid Constituent Andrographolide. J. Health Sci. 2008, 54, 370–381. [Google Scholar] [CrossRef]
- Jayakumar, T.; Hsieh, C.-Y.; Lee, J.-J.; Sheu, J.-R. Experimental and Clinical Pharmacology of Andrographis paniculata and Its Major Bioactive Phytoconstituent Andrographolide. Evid. Based Complement. Altern. Med. 2013, 2013, 846740. [Google Scholar] [CrossRef]
- Hu, X.-Y.; Wu, R.-H.; Logue, M.; Blondel, C.; Lai, L.Y.W.; Stuart, B.; Flower, A.; Fei, Y.-T.; Moore, M.; Shepherd, J.; et al. Andrographis paniculata (Chuān Xīn Lián) for symptomatic relief of acute respiratory tract infections in adults and children: A systematic review and meta-analysis. PLoS ONE 2017, 12, e0181780. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.M.; But, P.P.-H. Pharmacology and Applications Chinese Materia Medica; World Scientific Publishing Co. Pte. Ltd.: Singapore, 1987; Volume 2, p. 716. [Google Scholar]
- Chantrapromma, S.; Boonnak, N.; Pitakpornpreecha, T.; Yordthong, T.; Chidan Kumar, C.S.; Fun, H.K. Absolute Configuration of Andrographolide and Its Proliferation of Osteoblast Cell Lines. Crystallogr. Rep. 2018, 63, 412–417. [Google Scholar] [CrossRef]
- Khan, I.; Khan, F.; Farooqui, A.; Ansari, I.A. Andrographolide Exhibits Anticancer Potential Against Human Colon Cancer Cells by Inducing Cell Cycle Arrest and Programmed Cell Death via Augmentation of Intracellular Reactive Oxygen Species Level. Nutr. Cancer 2018, 70, 787–803. [Google Scholar] [CrossRef]
- Liao, H.-C.; Chou, Y.-J.; Lin, C.-C.; Liu, S.-H.; Oswita, A.; Huang, Y.-L.; Wang, Y.-L.; Syu, J.-L.; Sun, C.-M.; Leu, C.-M.; et al. Andrographolide and its potent derivative exhibit anticancer effects against imatinib-resistant chronic myeloid leukemia cells by downregulating the Bcr-Abl oncoprotein. Biochem. Pharmacol. 2019, 163, 308–320. [Google Scholar] [CrossRef]
- Chua, L.S. Review on liver inflammation and antiinflammatory activity of Andrographis paniculata for hepatoprotection. Phytother. Res. 2014, 28, 1589–1598. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.S.D.; Liao, W.; Zhou, S.; Wong, W.S.F. Is there a future for andrographolide to be an anti-inflammatory drug? Deciphering its major mechanisms of action. Biochem. Pharmacol. 2017, 139, 71–81. [Google Scholar] [CrossRef]
- Yongyun, C.; Jingwei, Z.; Zhiqing, L.; Wenxiang, C.; Huiwu, L. Andrographolide stimulates osteoblastogenesis and bone formation by inhibiting nuclear factor kappa-Β signaling both in vivo and in vitro. J. Orthop. Transl. 2019, 19, 47–57. [Google Scholar] [CrossRef]
- Zhai, Z.J.; Li, H.W.; Liu, G.W.; Qu, X.H.; Tian, B.; Yan, W.; Lin, Z.; Tang, T.T.; Qin, A.; Dai, K.R. Andrographolide suppresses RANKL-induced osteoclastogenesis in vitro and prevents inflammatory bone loss in vivo. Br. J. Pharmacol. 2014, 171, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Liu, Q.; Zhou, L.; Yuan, J.B.; Lin, X.; Zeng, R.; Liang, X.; Zhao, J.; Xu, J. Andrographolide Inhibits Ovariectomy-Induced Bone Loss via the Suppression of RANKL Signaling Pathways. Int. J. Mol. Sci. 2015, 16, 27470–27481. [Google Scholar] [CrossRef]
- Tangyuenyong, S.; Viriyakhasem, N.; Peansukmanee, S.; Kongtawelert, P.; Ongchai, S. Andrographolide Exerts Chondroprotective Activity in Equine Cartilage Explant and Suppresses Interleukin-1β-Induced MMP-2 Expression in Equine Chondrocyte Culture. Int. Sch. Res. Not. 2014, 2014, 464136. [Google Scholar] [CrossRef]
- Li, B.; Jiang, T.; Liu, H.; Miao, Z.; Fang, D.; Zheng, L.; Zhao, J. Andrographolide protects chondrocytes from oxidative stress injury by activation of the Keap1-Nrf2-Are signaling pathway. J. Cell. Physiol. 2018, 234, 561–571. [Google Scholar] [CrossRef]
- Ambili, R.; Janam, P.; Saneesh Babu, P.S.; Prasad, M.; Vinod, D.; Anil Kumar, P.R.; Kumary, T.V.; Asha Nair, S.; Radhakrishna Pillai, M. An ex vivo evaluation of the efficacy of andrographolide in modulating differential expression of transcription factors and target genes in periodontal cells and its potential role in treating periodontal diseases. J. Ethnopharmacol. 2017, 196, 160–167. [Google Scholar] [CrossRef]
- Hsieh, Y.-L.; Shibu, M.A.; Lii, C.-K.; Viswanadha, V.P.; Lin, Y.-L.; Lai, C.-H.; Chen, Y.-F.; Lin, K.-H.; Kuo, W.-W.; Huang, C.-Y. Andrographis paniculata extract attenuates pathological cardiac hypertrophy and apoptosis in high-fat diet fed mice. J. Ethnopharmacol. 2016, 192, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, C.; Berman, S.H.; Babish, J.G.; Ma, X.; Shinto, L.; Dorr, M.; Wells, K.; Wenner, C.A.; Standish, L.J. A phase I trial of andrographolide in HIV positive patients and normal volunteers. Phytother. Res. 2000, 14, 333–338. [Google Scholar] [CrossRef]
- Uttekar, M.M.; Das, T.; Pawar, R.S.; Bhandari, B.; Menon, V.; Nutan; Gupta, S.K.; Bhat, S.V. Anti-HIV activity of semisynthetic derivatives of andrographolide and computational study of HIV-1 gp120 protein binding. Eur. J. Med. Chem. 2012, 56, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Dey, D.; Jingar, P.; Agrawal, S.; Shrivastava, V.; Bhattacharya, A.; Manhas, J.; Garg, B.; Ansari, M.T.; Mridha, A.R.; Sreenivas, V.; et al. Symphytum officinale augments osteogenesis in human bone marrow-derived mesenchymal stem cells in vitro as they differentiate into osteoblasts. J. Ethnopharmacol. 2020, 248, 112329. [Google Scholar] [CrossRef]
- Liu, Z.; Lin, Y.; Fang, X.; Yang, J.; Chen, Z. Epigallocatechin-3-Gallate Promotes Osteo-/Odontogenic Differentiation of Stem Cells from the Apical Papilla through Activating the BMP–Smad Signaling Pathway. Molecules 2021, 26, 1580. [Google Scholar] [CrossRef]
- Ganguly, K.; Jeong, M.-S.; Dutta, S.D.; Patel, D.K.; Cho, S.-J.; Lim, K.-T. Protaetia brevitarsis seulensis Derived Protein Isolate with Enhanced Osteomodulatory and Antioxidative Property. Molecules 2020, 25, 6056. [Google Scholar] [CrossRef]
- Pholphana, N.; Rangkadilok, N.; Thongnest, S.; Ruchirawat, S.; Ruchirawat, M.; Satayavivad, J. Determination and variation of three active diterpenoids in Andrographis paniculata (Burm.f.) Nees. Phytochem. Anal. 2004, 15, 365–371. [Google Scholar] [CrossRef]
- Persson, M.; Lehenkari, P.P.; Berglin, L.; Turunen, S.; Finnilä, M.A.J.; Risteli, J.; Skrifvars, M.; Tuukkanen, J. Osteogenic Differentiation of Human Mesenchymal Stem cells in a 3D Woven Scaffold. Sci. Rep. 2018, 8, 10457. [Google Scholar] [CrossRef]
- Hanna, H.; Mir, L.M.; Andre, F.M. In vitro osteoblastic differentiation of mesenchymal stem cells generates cell layers with distinct properties. Stem Cell Res. Ther. 2018, 9, 203. [Google Scholar] [CrossRef]
- Aljohani, H.; Senbanjo, L.T.; Chellaiah, M.A. Methylsulfonylmethane increases osteogenesis and regulates the mineralization of the matrix by transglutaminase 2 in SHED cells. PLoS ONE 2019, 14, e0225598. [Google Scholar] [CrossRef]
- Chalise, J.P.; Hashimoto, S.; Parajuli, G.; Kang, S.; Singh, S.K.; Gemechu, Y.; Metwally, H.; Nyati, K.K.; Dubey, P.K.; Zaman, M.M.-U.; et al. Feedback regulation of Arid5a and Ppar-γ2 maintains adipose tissue homeostasis. Proc. Natl. Acad. Sci. USA 2019, 116, 15128–15133. [Google Scholar] [CrossRef]
- Jurek, S.; Sandhu, M.A.; Trappe, S.; Bermúdez-Peña, M.C.; Kolisek, M.; Sponder, G.; Aschenbach, J.R. Optimizing adipogenic transdifferentiation of bovine mesenchymal stem cells: A prominent role of ascorbic acid in FABP4 induction. Adipocyte 2020, 9, 35–50. [Google Scholar] [CrossRef]
- Venkatesan, J.K.; Meng, W.; Rey-Rico, A.; Schmitt, G.; Speicher-Mentges, S.; Falentin-Daudré, C.; Leroux, A.; Madry, H.; Migonney, V.; Cucchiarini, M. Enhanced Chondrogenic Differentiation Activities in Human Bone Marrow Aspirates via sox9 Overexpression Mediated by pNaSS-Grafted PCL Film-Guided rAAV Gene Transfer. Pharmaceutics 2020, 12, 280. [Google Scholar] [CrossRef] [PubMed]
- Bergholt, N.L.; Lysdahl, H.; Lind, M.; Foldager, C.B. A Standardized Method of Applying Toluidine Blue Metachromatic Staining for Assessment of Chondrogenesis. Cartilage 2019, 10, 370–374. [Google Scholar] [CrossRef]
- Katagiri, K.; Matsukura, Y.; Muneta, T.; Ozeki, N.; Mizuno, M.; Katano, H.; Sekiya, I. Fibrous Synovium Releases Higher Numbers of Mesenchymal Stem Cells Than Adipose Synovium in a Suspended Synovium Culture Model. Arthrosc. J. Arthrosc. Relat. Surg. 2017, 33, 800–810. [Google Scholar] [CrossRef] [PubMed]
- Bailey, A.M.; Kapur, S.; Katz, A.J. Characterization of adipose-derived stem cells: An update. Curr. Stem Cell Res. Ther. 2010, 5, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Hatakeyama, A.; Uchida, S.; Utsunomiya, H.; Tsukamoto, M.; Nakashima, H.; Nakamura, E.; Pascual-Garrido, C.; Sekiya, I.; Sakai, A. Isolation and Characterization of Synovial Mesenchymal Stem Cell Derived from Hip Joints: A Comparative Analysis with a Matched Control Knee Group. Stem Cells Int. 2017, 2017, 8–10. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Criado, I.; Meseguer-Ripolles, J.; Mellado-López, M.; Alastrue-Agudo, A.; Griffeth, R.J.; Forteza-Vila, J.; Cugat, R.; García, M.; Moreno-Manzano, V. Human Suprapatellar Fat Pad-Derived Mesenchymal Stem Cells Induce Chondrogenesis and Cartilage Repair in a Model of Severe Osteoarthritis. Stem Cells Int. 2017, 2017. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient No. | Age | Sex | Diagnosis |
|---|---|---|---|
| 1 | 81 | Female | Osteoarthritis |
| 2 | 75 | Male | Osteoarthritis |
| 3 | 61 | Female | Osteoarthritis |
| 4 | 78 | Female | Osteoarthritis |
| 5 | 76 | Female | Osteoarthritis |
| 6 | 86 | Female | Osteoarthritis |
| 7 | 77 | Female | Osteoarthritis |
| 8 | 83 | Female | Osteoarthritis |
| 9 | 75 | Female | Osteoarthritis |
| 10 | 81 | Female | Osteoarthritis |
| Gene | Forward Primer 5-3 | Reverse Primer 5-3 |
|---|---|---|
| Runx2 | 5′TATGGCACTTCTTCAGGATCC′3 | 5′GCGTCAACACCATCATTCTGG′3 |
| OPN | 5′TGAAACGAGTCAGCTGGATG′3 | 5′TGAAATTCATGGCTGTGGAA′3 |
| PPAR-γ2 | 5′GCTGTTATGGGTGAAACTCTG′3 | 5′ATAAGGTGGAGATGCAGGCTC′3 |
| LPL | 5′GAGATTTCTCTGTATGGCACC′3 | 5′CTGCAAATGAGACACTTTCTC′3 |
| Sox9 | 5′ATCTGAAGAAGGAGAGCGAG′3 | 5′TCAGAAGTCTCCAGAGCTTG′3 |
| Aggrecan | 5′TGAGGAGGGCTGGAACAAGTACC′3 | 5′GGAGGTGGTAATTGCAGGGAACA′3 |
| GAPDH | 5′GTGAAGGTCGGAGTCAACGG′3 | 5′TCAATGAAGGGGTCATTGATGG′3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kulsirirat, T.; Honsawek, S.; Takeda-Morishita, M.; Sinchaipanid, N.; Udomsinprasert, W.; Leanpolchareanchai, J.; Sathirakul, K. The Effects of Andrographolide on the Enhancement of Chondrogenesis and Osteogenesis in Human Suprapatellar Fat Pad Derived Mesenchymal Stem Cells. Molecules 2021, 26, 1831. https://doi.org/10.3390/molecules26071831
Kulsirirat T, Honsawek S, Takeda-Morishita M, Sinchaipanid N, Udomsinprasert W, Leanpolchareanchai J, Sathirakul K. The Effects of Andrographolide on the Enhancement of Chondrogenesis and Osteogenesis in Human Suprapatellar Fat Pad Derived Mesenchymal Stem Cells. Molecules. 2021; 26(7):1831. https://doi.org/10.3390/molecules26071831
Chicago/Turabian StyleKulsirirat, Thitianan, Sittisak Honsawek, Mariko Takeda-Morishita, Nuttanan Sinchaipanid, Wanvisa Udomsinprasert, Jiraporn Leanpolchareanchai, and Korbtham Sathirakul. 2021. "The Effects of Andrographolide on the Enhancement of Chondrogenesis and Osteogenesis in Human Suprapatellar Fat Pad Derived Mesenchymal Stem Cells" Molecules 26, no. 7: 1831. https://doi.org/10.3390/molecules26071831
APA StyleKulsirirat, T., Honsawek, S., Takeda-Morishita, M., Sinchaipanid, N., Udomsinprasert, W., Leanpolchareanchai, J., & Sathirakul, K. (2021). The Effects of Andrographolide on the Enhancement of Chondrogenesis and Osteogenesis in Human Suprapatellar Fat Pad Derived Mesenchymal Stem Cells. Molecules, 26(7), 1831. https://doi.org/10.3390/molecules26071831