
A Rationale for Drug Design Provided by Co-Crystal Structure of IC261 in Complex with Tubulin


Abstract
1. Introduction
2. Results
2.1. Visualizing Binding Modes of Tubulin-IC261 Complex
2.2. Comparison of the Binding Modes between IC261 and Colchicine
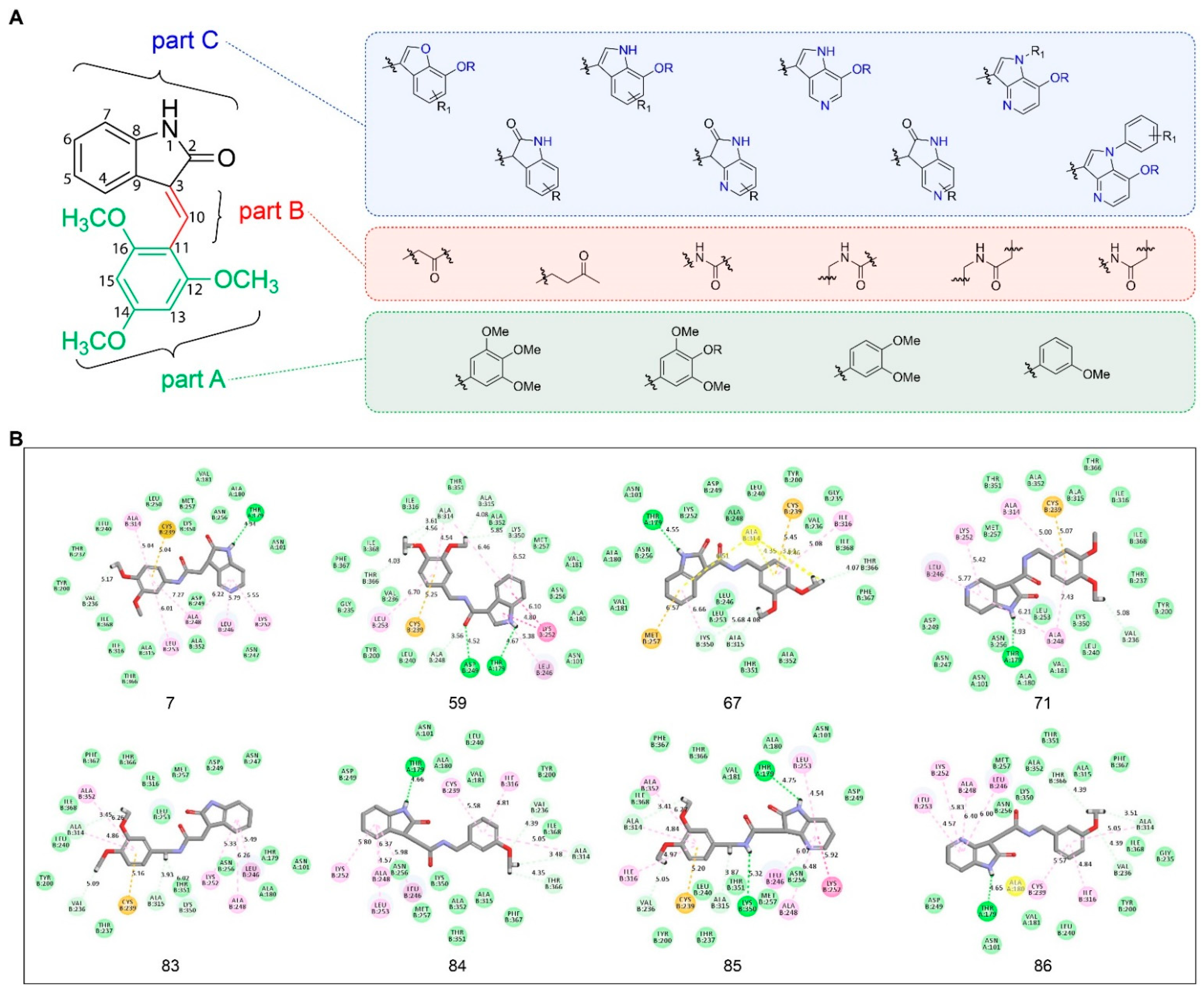
2.3. Insights into Drug Design and Molecular Docking Studies
3. Materials and Methods
3.1. Special Reagents
3.2. Protein Expression and Purification
3.3. Crystallization and Crystals Soaking
3.4. Data Collection and Structure Determination
3.5. Molecular Docking
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Brouhard, G.J.; Rice, L.M. Microtubule dynamics: An interplay of biochemistry and mechanics. Nat. Rev. Mol. Cell Biol. 2018, 19, 451–463. [Google Scholar] [CrossRef]
- Goodson, H.V.; Jonasson, E.M. Microtubules and Microtubule-Associated Proteins. Cold Spring Harb. Perspect. Biol. 2018, 10, a022608. [Google Scholar] [CrossRef] [PubMed]
- Borisy, G.; Heald, R.; Howard, J.; Janke, C.; Musacchio, A.; Nogales, E. Microtubules: 50 years on from the discovery of tubulin. Nat. Rev. Mol. Cell Biol. 2016, 17, 322–328. [Google Scholar] [CrossRef]
- Janke, C.; Magiera, M.M. The tubulin code and its role in controlling microtubule properties and functions. Nat. Rev. Mol. Cell Biol. 2020, 21, 307–326. [Google Scholar] [CrossRef]
- Stanton, R.A.; Gernert, K.M.; Nettles, J.H.; Aneja, R. Drugs that target dynamic microtubules: A new molecular perspective. Med. Res. Rev. 2011, 31, 443–481. [Google Scholar] [CrossRef] [PubMed]
- McLoughlin, E.C.; O’Boyle, N.M. Colchicine-Binding Site Inhibitors from Chemistry to Clinic: A Review. Pharmaceuticals 2020, 13, 8. [Google Scholar] [CrossRef] [PubMed]
- Gigant, B.; Wang, C.; Ravelli, R.B.; Roussi, F.; Steinmetz, M.O.; Curmi, P.A.; Sobel, A.; Knossow, M. Structural basis for the regulation of tubulin by vinblastine. Nature 2005, 435, 519–522. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Sun, X. Regulation of paclitaxel activity by microtubule-associated proteins in cancer chemotherapy. Cancer Chemother. Pharmacol. 2017, 80, 909–917. [Google Scholar] [CrossRef] [PubMed]
- Dorléans, A.; Gigant, B.; Ravelli, R.B.; Mailliet, P.; Mikol, V.; Knossow, M. Variations in the colchicine-binding domain provide insight into the structural switch of tubulin. Proc. Natl. Acad. Sci. USA 2009, 106, 13775–13779. [Google Scholar] [CrossRef]
- Yusuf, R.Z.; Duan, Z.; Lamendola, D.E.; Penson, R.T.; Seiden, M.V. Paclitaxel resistance: Molecular mechanisms and pharmacologic manipulation. Curr. Cancer Drug Targets 2003, 3, 1–19. [Google Scholar] [CrossRef]
- Jiang, J. CK1 in Developmental Signaling: Hedgehog and Wnt. Curr. Top. Dev. Biol. 2017, 123, 303–329. [Google Scholar] [PubMed]
- Narasimamurthy, R.; Hunt, S.R.; Lu, Y.; Fustin, J.M.; Okamura, H.; Partch, C.L.; Forger, D.B.; Kim, J.K.; Virshup, D.M. CK1δ/ε protein kinase primes the PER2 circadian phosphoswitch. Proc. Natl. Acad. Sci. USA 2018, 115, 5986–5991. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Qin, S.; Atangan, L.I.; Molina, Y.; Okawa, Y.; Arpawong, H.T.; Ghosn, C.; Xiao, J.H.; Vuligonda, V.; Brown, G.; et al. Casein kinase 1alpha interacts with retinoid X receptor and interferes with agonist-induced apoptosis. J. Biol. Chem. 2004, 279, 30844–30849. [Google Scholar] [CrossRef]
- Lee, S.Y.; Kim, H.; Li, C.M.; Kang, J.; Najafov, A.; Jung, M.; Kang, S.; Wang, S.; Yuan, J.; Jung, Y.K. Casein kinase-1γ1 and 3 stimulate tumor necrosis factor-induced necroptosis through RIPK3. Cell Death Dis. 2019, 10, 923. [Google Scholar] [CrossRef] [PubMed]
- Mashhoon, N.; DeMaggio, A.J.; Tereshko, V.; Bergmeier, S.C.; Egli, M.; Hoekstra, M.F.; Kuret, J. Crystal structure of a conformation-selective casein kinase-1 inhibitor. J. Biol. Chem. 2000, 275, 20052–20060. [Google Scholar] [CrossRef]
- Cheong, J.K.; Nguyen, T.H.; Wang, H.; Tan, P.; Voorhoeve, P.M.; Lee, S.H.; Virshup, D.M. IC261 induces cell cycle arrest and apoptosis of human cancer cells via CK1δ/ɛ and Wnt/β-catenin independent inhibition of mitotic spindle formation. Oncogene 2011, 30, 2558–2569. [Google Scholar] [CrossRef]
- Stöter, M.; Krüger, M.; Banting, G.; Henne-Bruns, D.; Knippschild, U. Microtubules depolymerization caused by the CK1 inhibitor IC261 may be not mediated by CK1 blockage. PLoS ONE 2014, 9, e100090. [Google Scholar] [CrossRef] [PubMed]
- Kremmidiotis, G.; Leske, A.F.; Lavranos, T.C.; Beaumont, D.; Gasic, J.; Hall, A.; O’Callaghan, M.; Matthews, C.A.; Flynn, B. BNC105: A novel tubulin polymerization inhibitor that selectively disrupts tumor vasculature and displays single-agent antitumor efficacy. Mol. Cancer Ther. 2010, 9, 1562–1573. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Wu, C.; Wang, C.; Zhang, G.; Arnst, K.E.; Yao, Y.; Zhang, Z.; Wang, Y.; Pu, D.; Li, W. Unraveling the molecular mechanism of BNC105, a phase II clinical trial vascular disrupting agent, provides insights into drug design. Biochem. Biophys. Res. Commun. 2020, 525, 148–154. [Google Scholar] [CrossRef]
- Li, Y.; Yang, J.; Niu, L.; Hu, D.; Li, H.; Chen, L.; Yu, Y.; Chen, Q. Structural insights into the design of indole derivatives as tubulin polymerization inhibitors. FEBS Lett. 2020, 594, 199–204. [Google Scholar] [CrossRef]
- Ravelli, R.B.; Gigant, B.; Curmi, P.A.; Jourdain, I.; Lachkar, S.; Sobel, A.; Knossow, M. Insight into tubulin regulation from a complex with colchicine and a stathmin-like domain. Nature 2004, 428, 198–202. [Google Scholar] [CrossRef]
- Hartung, E.F. History of the use of colchicum and related medicaments in gout; with suggestions for further research. Ann. Rheum. Dis. 1954, 13, 190–200. [Google Scholar] [CrossRef]
- Li, L.; Jiang, S.; Li, X.; Liu, Y.; Su, J.; Chen, J. Recent advances in trimethoxyphenyl (TMP) based tubulin inhibitors targeting the colchicine binding site. Eur. J. Med. Chem. 2018, 151, 482–494. [Google Scholar] [CrossRef]
- Steinmetz, M.O.; Prota, A.E. Microtubule-Targeting Agents: Strategies To Hijack the Cytoskeleton. Trends Cell Biol. 2018, 28, 776–792. [Google Scholar] [CrossRef]
- Ghawanmeh, A.A.; Chong, K.F.; Sarkar, S.M.; Bakar, M.A.; Othaman, R.; Khalid, R.M. Colchicine prodrugs and codrugs: Chemistry and bioactivities. Eur. J. Med. Chem. 2018, 144, 229–242. [Google Scholar] [CrossRef]
- Prota, A.E.; Bargsten, K.; Zurwerra, D.; Field, J.J.; Díaz, J.F.; Altmann, K.H.; Steinmetz, M.O. Molecular mechanism of action of microtubule-stabilizing anticancer agents. Science 2013, 339, 587–590. [Google Scholar] [CrossRef]
- Charbaut, E.; Curmi, P.A.; Ozon, S.; Lachkar, S.; Redeker, V.; Sobel, A. Stathmin family proteins display specific molecular and tubulin binding properties. J. Biol. Chem. 2001, 276, 16146–16154. [Google Scholar] [CrossRef] [PubMed]
- Otwinowski, Z.; Minor, W. Processing of X-ray diffraction data collected in oscillation mode. Methods Enzymol. 1997, 276, 307–326. [Google Scholar]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. D Biol. Crystallogr. 2004, 60 Pt 12, 2126–2132. [Google Scholar] [CrossRef] [PubMed]
- Adams, P.D.; Afonine, P.V.; Bunkóczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D Biol. Crystallogr. 2010, 66 Pt 2, 213–221. [Google Scholar] [CrossRef]
- Chen, V.B.; Arendall, W.B., 3rd; Headd, J.J.; Keedy, D.A.; Immormino, R.M.; Kapral, G.J.; Murray, L.W.; Richardson, J.S.; Richardson, D.C. MolProbity: All-atom structure validation for macromolecular crystallography. Acta Crystallogr D Biol Crystallogr. 2010, 66 Pt 1, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Höttecke, N.; Liebeck, M.; Baumann, K.; Schubenel, R.; Winkler, E.; Steiner, H.; Schmidt, B. Inhibition of gamma-secretase by the CK1 inhibitor IC261 does not depend on CK1delta. Bioorg. Med. Chem. Lett. 2010, 20, 2958–2963. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data | Tubulin/IC261 |
|---|---|
| X-Ray Source | SSRF-BL18U1 |
| Integration Package | HKL2000 |
| Data Collection | |
| Resolution Range (Å) | 49.87–2.85 |
| Space Group | P 212121 |
| Unit Cell (Å, °) | 105.1 157.7 182.1 90, 90, 90 |
| Total Reflections | 931,632 |
| Unique Reflections | 71,664 |
| Redundancy | 13.3 (13.3) |
| Completeness (%) | 100.0 (100.0) |
| Mean I/sigma (I) | 15.5 (2) |
| Rmerge | 0.177 (0.933) |
| CC1/2 | 0.995 (0.833) |
| Structure Refinement | |
| R-Factor/R-Free | 0.1864/0.1903 |
| Root Mean Square (RMS) (Bonds) | 0.01 |
| RMS (Angles) | 0.17 |
| No. of Atoms | 17,757 |
| Protein | 17,520 |
| Ligands | 237 |
| Waters | 0 |
| Average B-Factor | 50.23 |
| Protein | 50.18 |
| Others | 50.83 |
| Ramachandran Plot Statistics | |
| Most Favored Regions (%) | 96.84 |
| Allowed Regions (%) | 3.16 |
| Disallowed Regions (%) | 0 |
| ID | CDOCKER Energy | CDOCKER Interaction Energy |
|---|---|---|
| IC261-85 | −28.35 | −55.01 |
| IC261-67 | −27.68 | −53.24 |
| IC261-7 | −19.01 | −52.47 |
| IC261-83 | −29.90 | −52.16 |
| IC261-59 | −33.53 | −51.44 |
| IC261-71 | −24.69 | −51.38 |
| IC261-86 | −29.65 | −51.33 |
| IC261-84 | −28.82 | −51.13 |
| IC261 | −5.65 | −50.76 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xian, J.; Bu, F.; Wang, Y.; Long, F.; Zhang, Z.; Wu, C.; Tao, Y.; Wang, T.; Wang, G. A Rationale for Drug Design Provided by Co-Crystal Structure of IC261 in Complex with Tubulin. Molecules 2021, 26, 946. https://doi.org/10.3390/molecules26040946
Xian J, Bu F, Wang Y, Long F, Zhang Z, Wu C, Tao Y, Wang T, Wang G. A Rationale for Drug Design Provided by Co-Crystal Structure of IC261 in Complex with Tubulin. Molecules. 2021; 26(4):946. https://doi.org/10.3390/molecules26040946
Chicago/Turabian StyleXian, Jinghong, Faqian Bu, Yuxi Wang, Fangyi Long, Zhixiong Zhang, Chengyong Wu, Yiran Tao, Ting Wang, and Guan Wang. 2021. "A Rationale for Drug Design Provided by Co-Crystal Structure of IC261 in Complex with Tubulin" Molecules 26, no. 4: 946. https://doi.org/10.3390/molecules26040946
APA StyleXian, J., Bu, F., Wang, Y., Long, F., Zhang, Z., Wu, C., Tao, Y., Wang, T., & Wang, G. (2021). A Rationale for Drug Design Provided by Co-Crystal Structure of IC261 in Complex with Tubulin. Molecules, 26(4), 946. https://doi.org/10.3390/molecules26040946