Abstract
In developing countries, crop deterioration is mainly caused by inappropriate storage conditions that promote insect infestation. Synthetic pesticides are associated with serious adverse effects on humans and the environment. Thus, finding alternative “green” insecticides is a very pressing need. Calotropis procera (Aiton) Dryand (Apocynaceae) growing in Saudi Arabia was selected for this purpose. LC-MS/MS analysis was applied to investigate the metabolic composition of different C. procera extracts. Particularly, C. procera latex and leaves showed a high presence of cardenolides including calactin, uscharidin, 15β-hydroxy-calactin, 16β-hydroxy-calactin, and 12β-hydroxy-calactin. The ovicidal activity of the extracts from different plant organs (flowers, leaves, branches, roots), and of the latex, against Cadra cautella (Walker) (Lepidoptera, Pyralidae) was assessed. Extracts of C. procera roots displayed the most potent activity with 50% of C. cautella eggs not hatching at 10.000 ppm (1%).
1. Introduction
One third of the crops produced worldwide are estimated to be lost in the postharvest period. In developing countries, this loss is mainly caused by inappropriate storage conditions that promote insect infestation, and insect pests are responsible for losses as great as 40% in grain crops [1,2]. For this reason, chemical insecticides have been widely used to protect grains from insect attacks, but the serious adverse effects of synthetic pesticides on humans are a major health concern. Acute exposure to chemical insecticides can lead to respiratory tract issues, skin and eye irritation, allergy, extreme weakness, and seizures, while chronic exposure increases the incidence of Parkinson’s disease and cancer [3,4]. Consequently, in recent years, many synthetic insecticides—such as chlorine, some organophosphorus compounds, and some carbamate derivatives—have been banned in various countries [5].
Conversely, botanical insecticides containing active components derived from plant extracts are considered a safe and environmentally friendly option for integrated pest management (IPM) [6]. The rationale for using botanical insecticides for pest control is that plants are able to defend themselves against their insect enemies by producing bioactive substances [7,8], and these compounds can be extracted and utilized on crops. The main advantage of botanical insecticides is that they can control insect pests while minimizing the side effects on non-target organisms and maintaining an ecological balance. In addition, botanical compounds decompose more quickly and easily than their synthetic counterparts.
One source of natural insecticides is C. procera (Aiton), a plant belonging to the Apocynaceae family. It is native to northern and tropical Africa, western and southern Asia, Indochina, and the Middle East [9]. It is characterized by a wide range of pharmacological properties [10,11] and is used in traditional medicine for the treatment of various diseases ranging from ulcers to leprosy, as well as spleen and liver diseases [12]. Its extracts have anti-bacterial, anti-inflammatory, and analgesic effects [13]. Moreover, C. procera also shows larvicidal activity against disease-carrying mosquitoes, such as Culex quinquefasciatus, Anopheles stephensi Liston, Culex tritaeniorhynchus Giles, and Culex gelidus (Theobald) (Diptera: Culicidae) [14,15,16,17,18], as well as repellent and oviposition deterrent effects against Anopheles arabiensis and Culex quinquefasciatus [19]. C. procera latex and its flavonoids were also shown to negatively affect the feeding behavior of the lesser grain borer, Rhyzopertha dominica (Fabricius) (Coleoptera: Bostrichidae) [20]. Furthermore, C. procera has insecticidal effects against foodstuff insects that cause food deterioration, such as Tribolium confusum and T. castanum [21,22].
C. procera is a desert plant that is native to and available in large quantities in Saudi Arabia [23]. Therefore, the aim of this work was to study the phytochemical profile of extracts from the flowers, leaves, branches, roots, and latex of C. procera. Extracts prepared from the different plant parts were analyzed and tested for their ovicidal activity against the almond moth, C. cautella (Walker) (Lepidoptera: Pyralidae), a stored-product pest that infests grains and dried fruits. To the best of our knowledge, no previous work had investigated the effects of C. procera extracts on this pest.
Plants like C. procera produce many secondary metabolites with various physiological and biological activities that can include deterrent and antifeedant activity [24]. Many plants with antifeedant properties have been evaluated as crop protectors and a source of green pesticides. These botanical insecticides are now proposed as attractive alternatives to synthetic chemical insecticides for pest management [25,26,27,28]. Botanical insecticides notably do not impose any threats to the environment or human health, unlike their synthetic chemical counterparts. However, the use of plants as botanical insecticides requires an availability of those plants on an industrial scale. One important example of a green pesticide plant species is the neem tree, Azadirachta indica A. Juss., which is extracted on a large industrial scale in India and around the world [29]. C. procera is also a good candidate as a green insecticide based on its deterrent effect against several insects.
In the present study, we hypothesized that C. procera is a potential commercial source of biodegradable insecticide because of its extensive distribution and plentiful biomass in temperate and tropical regions. Mass production on an industrial scale is feasible and could satisfy the increased demand for greener insecticides. Therefore, the aim of this study was to extract natural insecticide products to control insect and pest populations without affecting the ecological balance [30,31,32,33]. Here, liquid chromatography-mass spectrometry (LC-MS) analysis of the chemical contents from different organs and latex of C. procera revealed the presence of 37 constituents belonging to the cardenolide and flavonoid classes. The evaluation of the ovicidal activity on C. cautella eggs revealed that extracts of C. procera roots had the most potent activity and prevented the hatching of 50% of C. cautella eggs when administered at 10,000 ppm.
2. Results and Discussion
2.1. Chemical Characterization of Extracts
Previously published data suggest that the anti-microbial activity of C. procera extracts is related to their content in cardenolides [18]. In the present work, liquid chromatography coupled with high resolution mass spectrometry was used to study the metabolic composition of different organs of C. procera. By using liquid chromatography coupled with electrospray ionization to Orbitrap mass spectrometry technique (LC-ESI-Orbitrap-MS/MS), 37 compounds were identified or tentatively identified on the bases of their retention time, MS spectra, and MS fragmentation patterns. Thus, we carried out a preliminary nuclear magnetic resonance (NMR)-based investigation of the extracts to confirm the presence of such compounds (Figures S1–S5). The n-hexane extracts were found to contain mainly waxes and, therefore, they were not analyzed further. The methanol extracts showed the presence of cardenolides with aldehydic functions and flavonoid derivatives (Figures S1–S5). The methanol extract from the leaves, which was the most abundant, were subjected to phytochemical investigation to isolate pure compounds. The cardenolides that were isolated and characterized are reported in Figure 1.
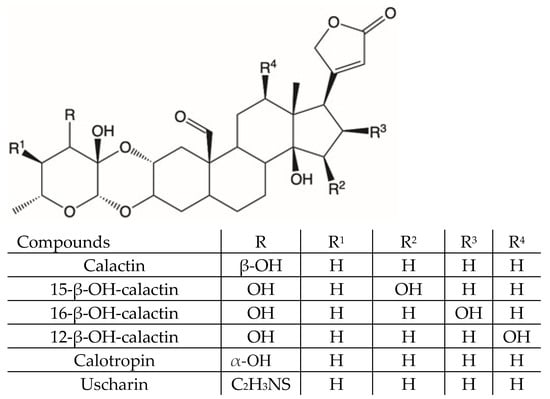
Figure 1.
Cardenolides isolated from Calotropis procera leaves.
2.2. LC-MS/MS Analysis
The methanolic extract of different plant organs (e.g., flowers, leaves, branches, roots), and the ethylacetate and butanolic extracts of latex, were then subjected to LC-MS/MS analysis in order to obtain a complete profile of the specialized metabolites and to compare their level in the extract. Resulting chromatograms, acquired in positive ion mode, are reported in Figure 2.
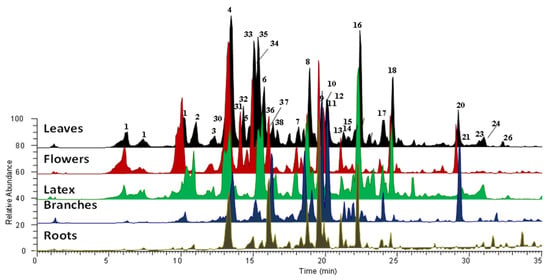
Figure 2.
Extracted ion chromatograms, registered in positive ion mode, of methanolic extracts of different organs and latex of C. procera.
The identification of all compounds was based on the accurate mass value, mass fragmentation spectra, literature data comparisons, and, in some cases, by the use of pure compounds. Metabolites belonging to the flavonoid and cardenolide classes of chemicals were recognized (Table 1 and Figure 2). All of the organs and latex for C. procera from Saudi Arabia were found to be rich in cardenolides compounds, particularly calactin and its derivatives.

Table 1.
MS Data of Compounds 1–37 detected in leaves, latex, flowers, branches and roots extract of C. procera.
The flavonoid derivatives isorhamnetin, quercetin, and kaempferol were identified. Peaks 31, 33, 35, and 36 in the MS/MS spectra showed the same fragmentation ion at an m/z value of 317, which corresponded to isorhamnetin aglycone. Thus, based on the MS, MS/MS fragmentation, and literature data [42], these compounds were identified as isorhamnetin-hexoside-pentoside, isorhamnetin rutinoside, isorhamnetin robinoside, and two isorhamnetin hexoside isomers, respectively. Peak 30 was identified as rutin by a direct comparison to a reference compound. Finally, peaks 32, 34, and 37 corresponded to kaempferol derivatives, as their MS/MS spectra showed the same fragment ion at a m/z value of 287. These compounds were supposed to be kaempferol-rutinoside, kaempferol-robinoside, and kaempferol-glucoside; a subsequent injection of standard compounds led to a confirmation of these structures.
Compounds 1–29 were tentatively identified as cardenolides by comparing their HPLC elution order, HR-MS data, and HR-MS/MS data with previously reported data (Figure 2, Table 1) [34,35,36,37,38,39,40,41,42,43,44,45,46,47,48]. The cardenolide compounds exhibited very similar MS/MS spectra, i.e., several losses of H2O and CO, and for the glycoside compound, loss of the sugar unit [50]. Compounds 2, 3, 6, 8, and 10 showed a pseudo molecular ion at an m/z of 549.2622, which corresponded to the molecular formula of C29H40O10. Due to the MS/MS experiments, it was possible to assign different isobar compounds. Peaks 2, 3, and 6 exhibited the same fragmentation pathways. By comparison with reference samples, we were able to assign these peaks to isomers of hydroxy-calactin carrying the hydroxyl group in different positions (12, 15, and 16-hydroxycalactin). The fragment ion at m/z 405 in the MS/MS spectrum of the compound in peak 10, corresponding to a loss of the acidic residue, confirmed the presence of calactinic acid in the structure of this molecule. Finally, peak 8 was identified as calotoxin, and the MS/MS spectrum showed a loss of a hexose. Compounds 16 and 18 corresponded to calactin and calotropin, respectively. The fragment ion 387 detected in their MS/MS spectra was diagnostic of the presence of the aglycone calotropagenin (peak 4), as already observed in the MS/MS spectra of the previously described compounds (8 and 10). The presence of these analytes was confirmed by comparison with the isolated compounds. The same approach was used to identify the compounds having calotropagenin as aglycone, specifically compounds 1, 14, 17, 19, 21–24, and 26–29, which differed from each other only in the glycone unit [35]. Compounds 5 and 9 had the same aglycone, coroglaucigenin (peak 11). According to the MS/MS spectra, we could assign these two metabolites to hydroxy-coroglaucigenin and frugoside (coroglaucigenin-rhamnoside) [35]. Peaks 7, 12, and 15 corresponded to the uzarigenin (peak 20) derivatives uzarin, desglucouzarin, and C. procera saponin I, respectively. Finally, peaks 13 and 16, showed a similar MS/MS spectra, suggesting the presence of the aglycone afrogenin, and were assigned to afroside and labriformine, respectively.
2.3. Quantitative Analysis
In order to evaluate the amount of metabolites in the different plant parts, some of the most representative isolated compounds, belonging to cardenolide and flavonoid classes, were used to perform quantitative analysis in the extracts from organs and latex of C. procera. For this purpose, a fast and efficient LC-MS/MS analytical method was developed, optimized, and validated. Due to the use of the multiple-reaction monitoring (MRM) detection mode, it was possible to reach satisfactory lower limits of quantization for all compounds (see Material and Methods).
Quantitative analyses were carried out on different C. procera methanol extracts (latex, leaves, roots, branches, flowers). The mean values (±SD) calculated on the basis of the results obtained in at least three experiments showing similar results are reported (Figure 3). This analysis revealed that flavonoids were very abundant in the leaves (particularly isorhamnetin derivatives), while higher concentrations of cardenolide compounds were measured in the latex extract.
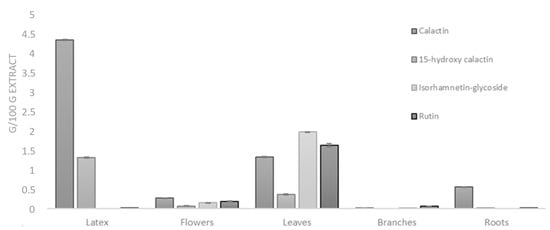
Figure 3.
Amounts (g/100 g extract) of calactin, 15-hydroxy calactin, isorhamnetin glycoside, and rutin measured in different organs and latex of C. procera via multiple-reaction monitoring (MRM)-based LC-MS/MS analyses.
2.4. C. procera Extracts Ovicidal Activity
The ovicidal activity of C. procera latex extracts ranged from 0% to 20% for methanolic and ethyl acetate extracts, respectively (Figure 4), with no significant differences among the extractants (F2,8 = 1.75; P = 0.252). This data was probably linked, according to the LC-MS analyses, to the presence of some cardenolides present, mainly in the ethyl acetate extract of latex such as compounds 19, 21, 25, and 27 (Table 1).
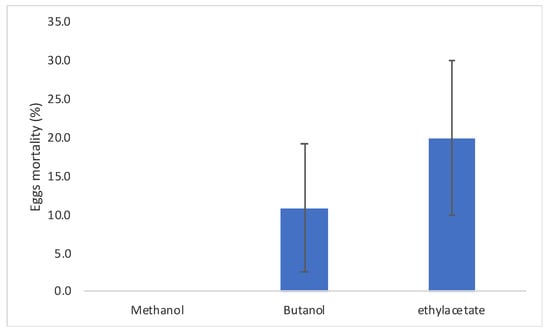
Figure 4.
Cadra cautella eggs mortality when treated with the C. procera latex extracts. Bars represent standard error.
As for the different plant organ extracts, the methanolic extracts of C. procera roots showed an ovicidal activity ranging from 36% to 52% of the treated eggs for leaves and roots extracts, respectively (Figure 5), with no significant differences among the plant organ extracts (F3,11 = 1.17; P = 0.378).
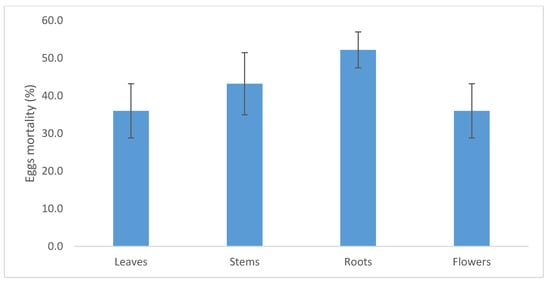
Figure 5.
Mortality of eggs of Cadra Cautella treated with the C. procera methanolic extracts of leaves, branches, roots, and flowers. Bars represent standard error.
In this research, C. procera root extracts showed clear ovicidal activity (50%). This finding is in line with the ovicidal activity of extracts from Acorus calamus (Araceae) (64%) roots. Further, it was higher than the Lamiaceae Vitex negundo (39%), Adhatoda vasica (15%) extracts, and Dioscorea deltoidea (Dioscoreaceae) (14%) extracts at a similar concentration (1.25%) against Plutella xylostella (Lepidoptera:Plutellidae) [51]. Similarly, Plantago lanceolata (Plantaginaceae) and Momordica charantia (Cucurbitaceae) methanolic extracts at 0.9% against Leucoptera coffeella (Lepidoptera: Lyonetiidae) eggs showed an ovicidal activity of 27% and 22%, respectively [52].
3. Materials and Methods
3.1. Plant Material
In 2017, C. procera plant material was collected in Makkah Al-Mukarramah between Arafat and Muzdalefah, Saudi Arabia (GPS coordinates 21°12′53.1′′ N, 40°17′05.3′′ E, altitude 286 m above sea level). Latex was collected by cutting the young and green stems. Leaves, branches, and flowers were collected from 20 plants. Roots were obtained from three middle sized plants. Plant organs were separately dried in shadows. The dried plant material was ground finely before extraction. All solvents used in the experiment were purchased from Sigma-Aldrich.
3.2. Extraction Methods
In this study, 50 g of each type of plant material—powdered leaves, flowers, stems, and roots—was extracted with 300 mL MeOH, followed by sonication for 30 min and centrifugation for 10 min at 2000 rpm. The supernatant was collected and evaporated under a vacuum. The latex was fractioned using a separatory funnel. Ethyl acetate was added and the mixture was vigorously shaken. After complete separation, the ethyl acetate layer was collected and filtered through filter paper to get rid of any latex traces. Then, the ethyl acetate extract evaporated under a vacuum using a rotary evaporator. The remaining latex was extracted with n-butanol by vigorous shaking. After complete separation of the two layers, the n-butanol fraction was evaporated under a vacuum using the rotary evaporator. Finally, methanol was added to the latex. After shaking, the methanolic layer was separated and evaporated as described above. The extract was separated on a silica gel column, the obtained fractions were further purified by reverse phase high performance liquid chromatography (RP-HPLC), and the isolated compounds were characterized by NMR experiments.
3.3. General Experimental Procedures
Optical rotations were measured by an Atago AP-300 digital polarimeter with a 1 dm microcell and sodium lamp (589 nm). NMR experiments were recorded on a Bruker Digital Receiver uniX 600 spectrometer at 300 K (Bruker BioSpin, Germany), revealing the spectra for methanol-d4 and CDCl3. HRESIMS data were obtained by using the positive ion modes on an Linear trap quadrupole LTQ Orbitrap XL mass spectrometer (Thermo Fisher Scientific). Quantitative data were acquired using an API6500 Q-Trap (ABSciex Foster City, CA, USA) apparatus.
3.4. Qualitative Analysis, LC-ESI-OrbitrapMS
The LC-MS method was applied to analyze specialized metabolites from different parts of C. procera. The adopted instrument configuration included an Accela (Thermo Fisher Scientific, Waltham, MA, USA) HPLC interfaced to a linear ion trap coupled with a high-resolution mass analyzer (LTQ-Orbitrap XL Thermo Fisher Scientific) through an Electrospray Ionization (EI) source. Separation was performed on a C18 column (Luna C18, Phenomenex, 100 × 2.0 mm, 2.5 µm) using a binary mobile phase composed of eluent A (ultrapure water–formic acid 0.1% v/v) and eluent B (ultrapure acetonitrile–formic acid 0.1% v/v). The separation conditions are 10% to 50% B within 40 min, followed by a second, faster gradient from 50% to 95% B within 10 min. The flow rate was 0.200 mL/min and the injection volume was 10.0 µL.
MS data were acquired in positive ion mode. At first, the full-mass and data-dependent scan mode was applied, and then tandem MS experiments were performed to identify the characteristic metabolites. The capillary temperature was set at 300 °C K, and the flow rate of sheath gas and auxiliary gas were set at 30.0 and 10.0 arbitrary units. The capillary voltage was 35.0 V, the source voltage was 3.5 kV, and the tube lens was 110 V. The mass resolution was set at 60,000.
3.5. Quantitative Analysis
The quantization of some isolated compounds, in particular calactin, 15-hydroxy calactin, rutin, and isorhamnetin-glycoside was carried out using an API6500 Q-Trap (ABSciex Foster City, CA, USA) coupled with a NexeraX2 UHPLC apparatus (Shimadzu, Kyoto, Japan). A positive multiple-reaction monitoring (MRM) system was selected for the LC-MS/MS analyses (Table 2). All the instrumental parameters were optimized by directly injecting solutions containing 10 mg/L of all of the pure compounds in methanol and water (50:50 v/v). All analyses of the MS data were performed in the positive ion mode. Samples of 5 μL were loaded onto a Luna Omega column (Phenomenex) (1.6 μm Polar C18 100 A, 50 mm × 2.1 mm), and compounds were separated using a linear gradient from 35% to 55% of acetonitrile (eluent B) in H2O containing 0.1% formic acid (eluent A) over 5 min. The flow rate was 0.4 mL min−1, and the injection volume was 5 μL for the standard compounds and latex extract samples. The total run time was 6 min. To perform accurate quantitative analyses, eight-point (0, 50, 150, 450, 1350, 4000, 12,000, and 24,000 ng/mL) calibration curves were built for the four purified compounds.
Table 2.
Technical and validation parameters of the LC/MS-based method used for the quantization of calactin, 15-hydroxy calactin, rutin, and isorhamnetin glycoside.
The linearity of the instrumental response as a function of sample concentration was confirmed for a wide range of concentrations (from 50 ng/mL to 24 μg/mL), and accuracy was fully satisfied (Table 2) for the investigated molecules.
3.6. Ovicidal Activity of C. procera Extracts on Cadra Cautella
The methanolic, butanolic, and ethyl acetate latex extracts were dissolved in methanol, n-butanol, and ethyl-acetate, respectively. Flower, leaf, root, and branch extracts were dissolved in DMSO 0.1% (aqueous solution). All solutions were tested at 10,000 ppm (1% concentration). Filter paper discs (5 cm diameter) treated with 100 μL of each extract solution, or solvent only for the controls, were placed inside a Petri dish (5 cm diameter). After the evaporation of the solvent, 50 eggs of C. cautella (0–24 h) were individually transferred onto the filter paper. Filter papers were kept wet with 50 μL of tap water. The test was replicated five times. The number of eggs unhatched was assessed after 24 h and calculated as a percentage of the total number of eggs. Experiments were conducted at 25 ± 1 °C, RH 65 ± 2%, and a photoperiod of 16:8 (L/D). C. cautella eggs mortalities were reported as a mean ± standard error (ES). Mortality was adjusted for the natural mortality of eggs [49] according to the following formula: Adjusted mortality (%) = 100 × (X−Y)/(100−Y), where X is the percentage mortality of the treated sample, and Y is the percentage mortality of the untreated control sample. Means were compared via one-way ANOVA with the extractant (latex extracts) or plant organ (plant extracts) as fixed factors. Equality of variances was checked before the analyses using the Levene’s test. Statistics were performed using SPSS 22.0 software (IBM SPSS Statistics, Armonk, North Castle, NY, USA).
4. Conclusions
In this study, we compared the phyto-constituents of different organs from an important medicinal plant widely distributed in the temperate regions, C. procera. In terms of large-scale extraction, this study could orient scientists when choosing organs rich in phyto-constituents. The title plant could be considered a source of biomolecules with potential anti-parasitic effects. Although the ovicidal activity is not as strong as that excreted by commercial synthetic molecules, our study contributes to the future development of safer and greener insecticides.
Supplementary Materials
The following are available online, Figure S1: 1H NMR spectrum of MeOH Extract of leaves, Figure S2. 1H NMR spectrum of MeOH Extract of flowers; Figure S3. 1H NMR spectrum of MeOH Extract of stems; Figure S4. 1H NMR spectrum of MeOH Extract of roots; Figure S5. 1H NMR spectrum of Latex Extract.
Author Contributions
Writing—original draft, data curation, and resources, A.B. and Z.O.; conceptualization and funding acquisition, A.I.A.-A.; investigation, formal analysis, writing—review and editing, and validation V.S., N.D.T., M.D., and F.D.P.; validation, visualization, and writing—review and editing, B.C. and S.B.; funding acquisition, project administration, and supervision, M.H. All authors have read and agreed to the published version of the manuscript.
Funding
This project was funded by King Abdulaziz City for Science and Technology, Kingdom of Saudi Arabia, Project Number: A R P-Large-123-35.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
Authors gratefully appreciate King Abdullah International Medical Research Center, King Saud Bin Abdulaziz University for Health Sciences for its precious support to conduct this Project.
Conflicts of Interest
The authors declare that there is no conflict of interest.
Sample Availability
Samples of the calactin, rutin, 15-β-OH-calactin, isorhamnetin glycoside are available from the authors.
References
- Kumar, D.; Kalita, P. Reducing Postharvest Losses during Storage of Grain Crops to Strengthen Food Security in Developing Countries. Foods 2017, 6, 8. [Google Scholar] [CrossRef]
- Donatelli, M.; Magarey, R.D.; Bregaglio, S.; Willocquet, L.; Whish, J.P.M.; Savary, S. Modelling the impacts of pests and diseases on agricultural systems. Agric. Syst. 2017, 155, 213–224. [Google Scholar] [CrossRef]
- Jiao, Z.; Wu, Y.; Qu, S. Fenpropathrin induces degeneration of dopaminergic neurons via disruption of the mitochondrial quality control system. Cell Death Discov. 2020, 6, 78. [Google Scholar] [CrossRef] [PubMed]
- Damalas, C.A.; Eleftherohorinos, I.G. Pesticide exposure, safety issues, and risk assessment indicators. Int. J. Environ. Res. Public Health 2011, 8, 1402–1419. [Google Scholar] [CrossRef] [PubMed]
- Garry, V.F. Pesticides and children. Toxicol. Appl. Pharmacol. 2004, 198, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Martin-Guay, M.O.; Paquette, A.; Dupras, J.; Rivest, D. The new Green Revolution: Sustainable intensification of agriculture by intercropping. Sci. Total Environ. 2018, 615, 767–772. [Google Scholar] [CrossRef]
- Hummelbrunner, L.A.; Isman, M.B. Acute, sublethal, antifeedant, and synergistic effects of monoterpenoid essential oil compounds on the tobacco cutworm, Spodoptera litura (Lep., Noctuidae). J. Agric. Food Chem. 2001, 49, 715–720. [Google Scholar] [CrossRef]
- Benelli, G. Plant-borne ovicides in the fight against mosquito vectors of medical and veterinary importance: A systematic review. Parasitol. Res. 2015, 114, 3201–3212. [Google Scholar] [CrossRef]
- Rahman, M.A.; Wilcock, C.C. A taxonomic revision of Calotropis (Asclepiadaceae). Nord. J. Bot. 1991, 11, 301–308. [Google Scholar] [CrossRef]
- Sweidan, N.I.; Abu Zarga, M.H. Two novel cardenolides from Calotropis procera. J. Asian Nat. Prod. Res. 2015, 17, 900–907. [Google Scholar] [CrossRef]
- Dhalendra, G.; Rathore, P.; Satapathy, T.; Roy, A. Pharmacognostical, phytochemical and pharmacological study of Calotropis procera: A review. Res. J. Pharm. Technol. 2014, 7, 346–351. [Google Scholar]
- Chundattu, S.J.; Agrawal, V.K.; Ganesh, N. Phytochemical investigation of Calotropis procera. Arab. J. Chem. 2016, 9, S230–S234. [Google Scholar] [CrossRef]
- Meena, A.K.; Yadav, A.; Rao, M. Ayurvedic uses and pharmacological activities of Calotropis procera Linn. Asian J. Tradit. Med. 2011, 6, 45–53. [Google Scholar]
- Sharma, P.; Sharma, J.D. Evaluation of in vitro schizontocidal activity of plant parts of Calotropis procera—An ethnobotanical approach. J. Ethnopharmacol. 1999, 68, 83–95. [Google Scholar] [CrossRef]
- Rao, K.V.B.; Kumar, G.; Karthik, L.; Kirthi, A.V.; Jayaseelan, C.; Rahuman, A.A. Phytochemical composition, mosquito larvicidal, ovicidal and repellent activity of Calotropis procera against Culex tritaeniorhynchus and Culex gelidus. Bangladesh J. Pharmacol. 2012, 7, 63–69. [Google Scholar]
- Singh, R.; Mittal, P.; Dhiman, R. Laboratory study on larvicidal properties of leaf extract of Calotropis procera (Family-Asclepiadaceae) against mosquito larvae. J. Commun. Dis. 2005, 37, 109. [Google Scholar] [PubMed]
- Shahi, M.; Hanafi-Bojd, A.; Iranshahi, M.; Vatandoost, H.; Hanafi-Bojd, M. Larvicidal efficacy of latex and extract of Calotropis procera (Gentianales: Asclepiadaceae) against Culex quinquefasciatus and Anopheles stephensi (Diptera: Culicidae). J. Vector Borne Dis. 2010, 47, 185–188. [Google Scholar]
- Su, T.; Mulla, M.S. Antifeedancy of neem products containing Azadirachtin against Culex tarsalis and Culex quinquefasciatus (Diptera: Culicidae). J. Soc. Vector Ecol. 1998, 23, 114–122. [Google Scholar]
- Elimam, A.M.; Elmalik, K.H.; Ali, F.S. Efficacy of leaves extract of Calotropis procera Ait. (Asclepiadaceae) in controlling Anopheles arabiensis and Culex quinquefasciatus mosquitoes. Saudi J. Biol. Sci. 2009, 16, 95–100. [Google Scholar] [CrossRef]
- Elamir, E.E.; Almadiy, A.A.; Nenaah, G.E.; Alabas, A.A.; Alsaqri, H.S. Comparing six mathematical link function models of the antifeedant activity of lesser grain borer exposed to sub-lethal concentrations of some extracts from Calotropis procera. Bioengineered 2019, 10, 292–305. [Google Scholar] [CrossRef]
- Farrar, N.; Golestaneh, S.; Farsi, M.; Sadeghi, S.; Askari, H. Effectiveness of Extract of Calotropis procera on Nutritional Indices of Tribolium confusum Duv. In Proceedings of the XXVIII International Horticultural Congress on Science and Horticulture for People (IHC2010): International Symposium on the 940, Lisboa, Portugal, 22–27 August 2010; pp. 669–672. [Google Scholar]
- Khaliq, A.; Ullah, M.I.; Afzal, M.; Ali, S.; Sajjad, A.; Ahmad, A.; Khalid, S. Management of Tribolium castaneum using synergism between conventional fumigant and plant essential oils. Int. J. Trop. Insect Sci. 2020, 40, 781–788. [Google Scholar] [CrossRef]
- Zahran, M.; Younes, H.; Al-Tawil, B. Ecology of four community types: Red Sea coastal desert, Saudi Arabia. J. Coast. Res. 1985, 1, 279–288. [Google Scholar]
- Siciliano, T.; Leo, M.D.; Bader, A.; Tommasi, N.D.; Vrieling, K.; Braca, A.; Morelli, I. Pyrrolizidine alkaloids from Anchusa strigosa and their antifeedant activity. Phytochemistry 2005, 66, 1593–1600. [Google Scholar] [CrossRef] [PubMed]
- Duke, S.O.; Cantrell, C.L.; Meepagala, K.M.; Wedge, D.E.; Tabanca, N.; Schrader, K.K. Natural toxins for use in pest management. Toxins 2010, 2, 1943–1962. [Google Scholar] [CrossRef]
- Begum, N. Calotropis procera and Annona squamosa: Potential Alternatives to Chemical Pesticides. Br. J. Appl. Sci. Technol. 2013, 3, 254–267. [Google Scholar] [CrossRef]
- Nenaah, G. Antimicrobial activity of Calotropis procera Ait. (Asclepiadaceae) and isolation of four flavonoid glycosides as the active constituents. World J. Microbiol. Biotechnol. 2013, 29, 1255–1262. [Google Scholar] [CrossRef]
- Begum, N.; Sharma, B.; Pandey, R. Evaluation of Insecticidal Efficacy of Calotropis Procera and Annona Squamosa Ethanol Extracts against Musca Domestica. J. Biofertil. Biopestic. 2010, 1, 2–6. [Google Scholar] [CrossRef]
- Roy, S.; Handique, G.; Muraleedharan, N.; Dashora, K.; Roy, S.M.; Mukhopadhyay, A.; Babu, A. Use of plant extracts for tea pest management in India. Appl. Microbiol. Biotechnol. 2016, 100, 4831–4844. [Google Scholar] [CrossRef]
- Patil, S.V.; Patil, C.D.; Salunkhe, R.B.; Salunke, B.K. Larvicidal activities of six plants extracts against two mosquito species, Aedes aegypti and Anopheles stephensi. Trop. Biomed. 2010, 27, 360–365. [Google Scholar]
- Bedini, S.; Flamini, G.; Ascrizzi, R.; Venturi, F.; Ferroni, G.; Bader, A.; Girardi, J.; Conti, B. Essential oils sensory quality and their bioactivity against the mosquito Aedes albopictus. Sci. Rep. 2018, 8, 17857. [Google Scholar] [CrossRef]
- Ghosh, A.; Chowdhury, N.; Chandra, G. Plant extracts as potential mosquito larvicides. Indian J. Med. Res. 2012, 135, 581–598. [Google Scholar]
- Tahir, M.; Ishaq, T.; Mukhtar, M.; Khan, S. Potential use of Calotropis procera (Milk Weed) to Control Culex quinquefasciatus (Diptera: Culicidae). Pak. J. Zool. 2013, 45, 615–621. [Google Scholar]
- Ranjit, P.M.; Eswara Rao, G.; Krishnapriya, M.; Nagalakshmi, V.; Silpa, P.; Anjali, M. An overview of phytochemical and pharmacological activities of Calotropis procera. FS J. Pharm. Res. 2012, 1, 18–25. [Google Scholar]
- Pandey, A.; Swarnkar, V.; Pandey, T.; Srivastava, P.; Kanojiya, S.; Mishra, D.K.; Tripathi, V. Transcriptome and Metabolite analysis reveal candidate genes of the cardiac glycoside biosynthetic pathway from Calotropis procera. Sci. Rep. 2016, 6, 34464. [Google Scholar] [CrossRef]
- Hanna, A.G.; Shalaby, N.M.; Morsy, N.A.; Simon, A.; Toth, G.; Malik, S.; Duddeck, H. Structure of a calotropagenin-derived artifact from Calotropis procera. Magn. Reson. Chem. 2002, 40, 599–602. [Google Scholar] [CrossRef]
- Kanojiya, S.; Madhusudanan, K.P. Rapid identification of calotropagenin glycosides using high-performance liquid chromatography electrospray ionisation tandem mass spectrometry. Phytochem. Anal. 2012, 23, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Seiber, J.N.; JN, S.; CJ, N. Cardenolides in the latex and leaves of seven Asclepias species and Calotropis procera. Phytochemistry 1982, 21, 2343–2348. [Google Scholar] [CrossRef]
- Mohamed, N.H.; Liu, M.; Abdel-Mageed, W.M.; Alwahibi, L.H.; Dai, H.; Ismail, M.A.; Badr, G.; Quinn, R.J.; Liu, X.; Zhang, L.; et al. Cytotoxic cardenolides from the latex of Calotropis procera. Bioorg. Med. Chem. Lett. 2015, 25, 4615–4620. [Google Scholar] [CrossRef] [PubMed]
- Parihar, G.; Balekar, N. Calotropis procera: A phytochemical and pharmacological review. Thai J. Pharm. Sci. 2016, 40, 1–17. [Google Scholar]
- Crout, D.; Hassall, C.; Jones, T. 405. Cardenolides. Part VI. Uscharidin, calotropin, and calotoxin. J. Chem. Soc. 1964, 2187–2194. [Google Scholar] [CrossRef]
- Ahmad, V.U.; Basha, A. Spectroscopic Data of Steroid Glycosides: Spirostanes, Bufanolides, Cardenolides; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2010; Volume 3. [Google Scholar]
- Hassall, C.; Reyle, K. 18. Cardenolides. Part III. The constitution of calotropagenin. J. Chem. Soc. 1959, 85–89. [Google Scholar] [CrossRef]
- Elgamal, M.H.A.; Hanna, A.G.; Morsy, N.A.; Duddeck, H.; Simon, A.; Gáti, T.; Tóth, G. Complete 1H and 13C signal assignments of 5α-cardenolides isolated from Calotropis procera R. BR. J. Mol. Struct. 1999, 477, 201–208. [Google Scholar] [CrossRef]
- Ibrahim, S.R.; Mohamed, G.A.; Shaala, L.A.; Moreno, L.; Banuls, Y.; Kiss, R.; Youssef, D.T. Proceraside A, a new cardiac glycoside from the root barks of Calotropis procera with in vitro anticancer effects. Nat. Prod. Res. 2014, 28, 1322–1327. [Google Scholar] [CrossRef] [PubMed]
- Crout, D.; Curtis, R.; Hassall, C. 347. Cardenolides. Part V. The constitution of calactinic acid. J. Chem. Soc. 1963, 1866–1875. [Google Scholar] [CrossRef]
- Van Quaquebeke, E.; Simon, G.; André, A.; Dewelle, J.; El Yazidi, M.; Bruyneel, F.; Tuti, J.; Nacoulma, O.; Guissou, P.; Decaestecker, C.; et al. Identification of a novel cardenolide (2″-oxovoruscharin) from Calotropis procera and the hemisynthesis of novel derivatives displaying potent in vitro antitumor activities and high in vivo tolerance: Structure-activity relationship analyses. J. Med. Chem. 2005, 48, 849–856. [Google Scholar] [CrossRef]
- Shaker, K.H.; Morsy, N.; Zinecker, H.; Imhoff, J.F.; Schneider, B. Secondary metabolites from Calotropis procera (Aiton). Phytochem. Lett. 2010, 3, 212–216. [Google Scholar] [CrossRef]
- Bedini, S.; Farina, P.; Napoli, E.; Flamini, G.; Ascrizzi, R.; Verzera, A.; Conti, B.; Zappalà, L. Bioactivity of Different Chemotypes of Oregano Essential Oil against the Blowfly Calliphora vomitoria Vector of Foodborne Pathogens. Insects 2021, 12, 52. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, M.A.; Hamed, M.M.; Ahmed, W.S.; Abdou, A.M. Antioxidant and cytotoxic flavonols from Calotropis procera. Z. Nat. C 2011, 66, 547–554. [Google Scholar] [CrossRef]
- Matharu, K.; Mehta, P. Ovicidal activity of crude extracts of indigenous plant species against Plutella xylostella (L.) (Lepidoptera: Plutellidae). Environ. Ecol. 2017, 35, 285–289. [Google Scholar]
- Alves, D.S.; Oliveira, D.F.; Carvalho, G.A.; Santos, H.M.; Carvalho, D.A.; Santos, M.A.; Carvalho, H.W. Plant extracts as an alternative to control Leucoptera coffeella (Guérin-Mèneville) (Lepidoptera: Lyonetiidae). Neotrop. Entomol. 2011, 40, 123–128. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).