
Gut Microbiome and Degradation Product Formation during Biodegradation of Expanded Polystyrene by Mealworm Larvae under Different Feeding Strategies

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Biodegradation of PS and Next-Generation Sequencing of Gut Microbiome under Different Feeding Protocols
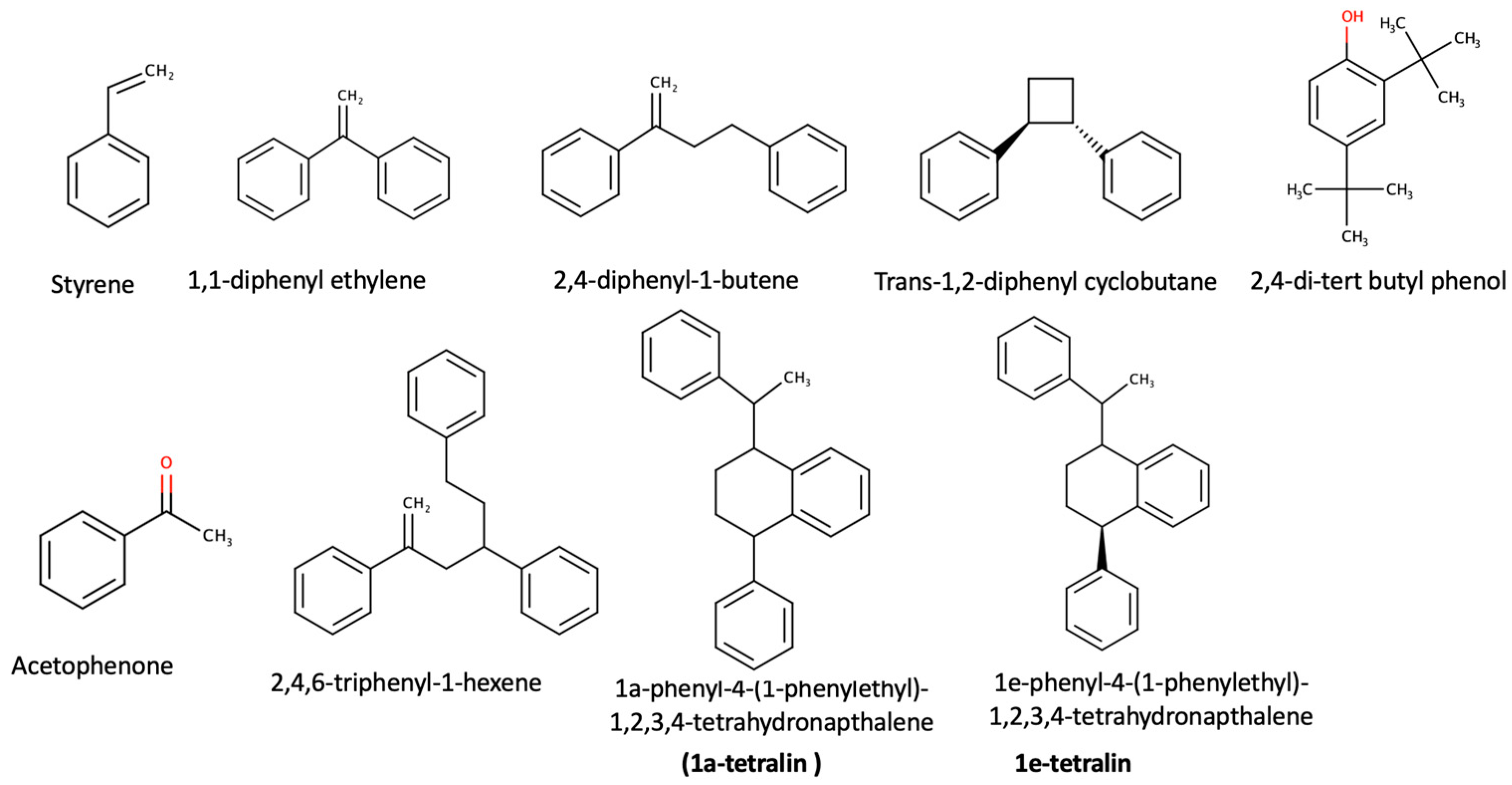
2.2. Chemical and Statistical Analysis
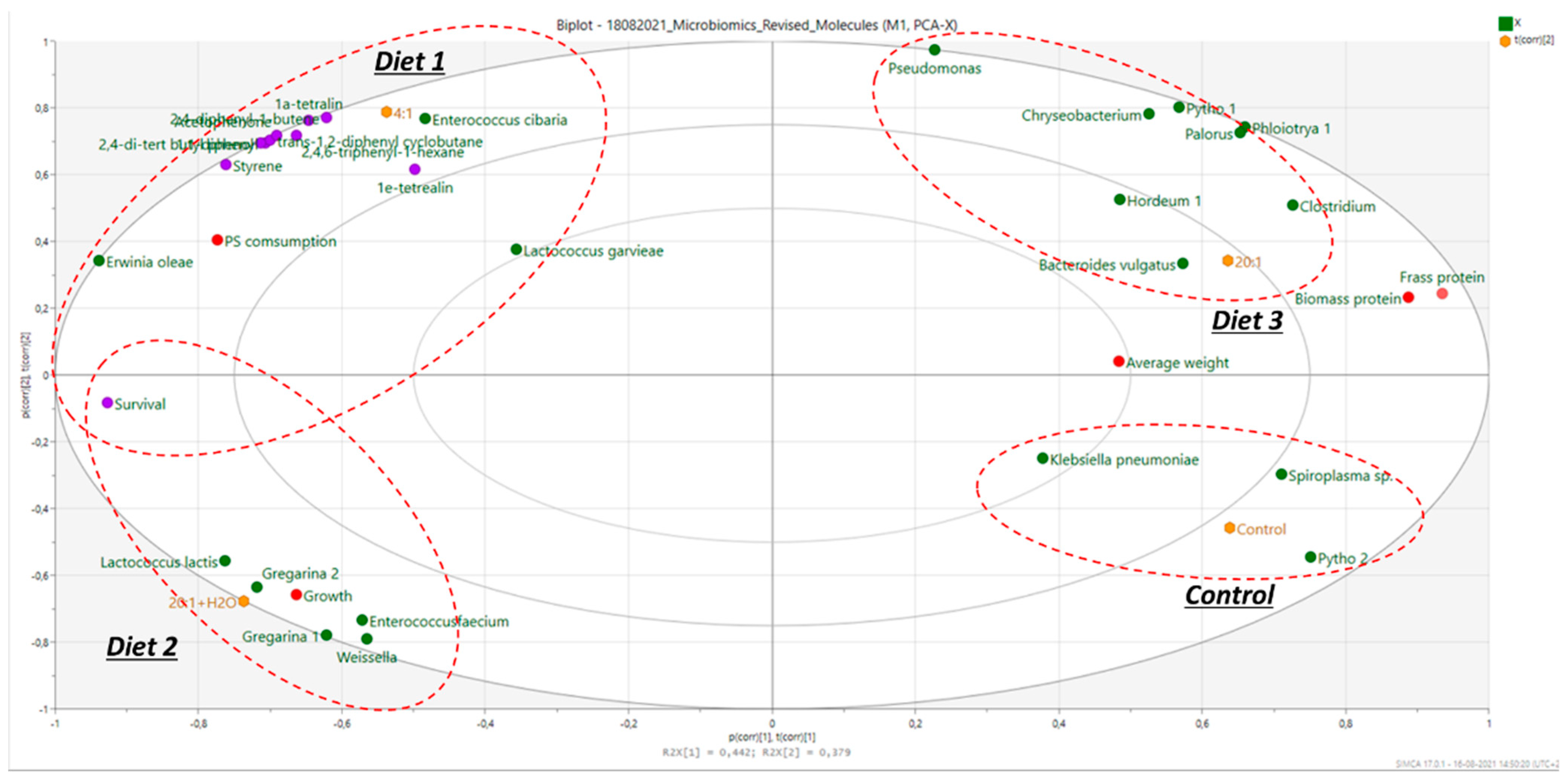
2.3. Multivariate Statistical Analysis
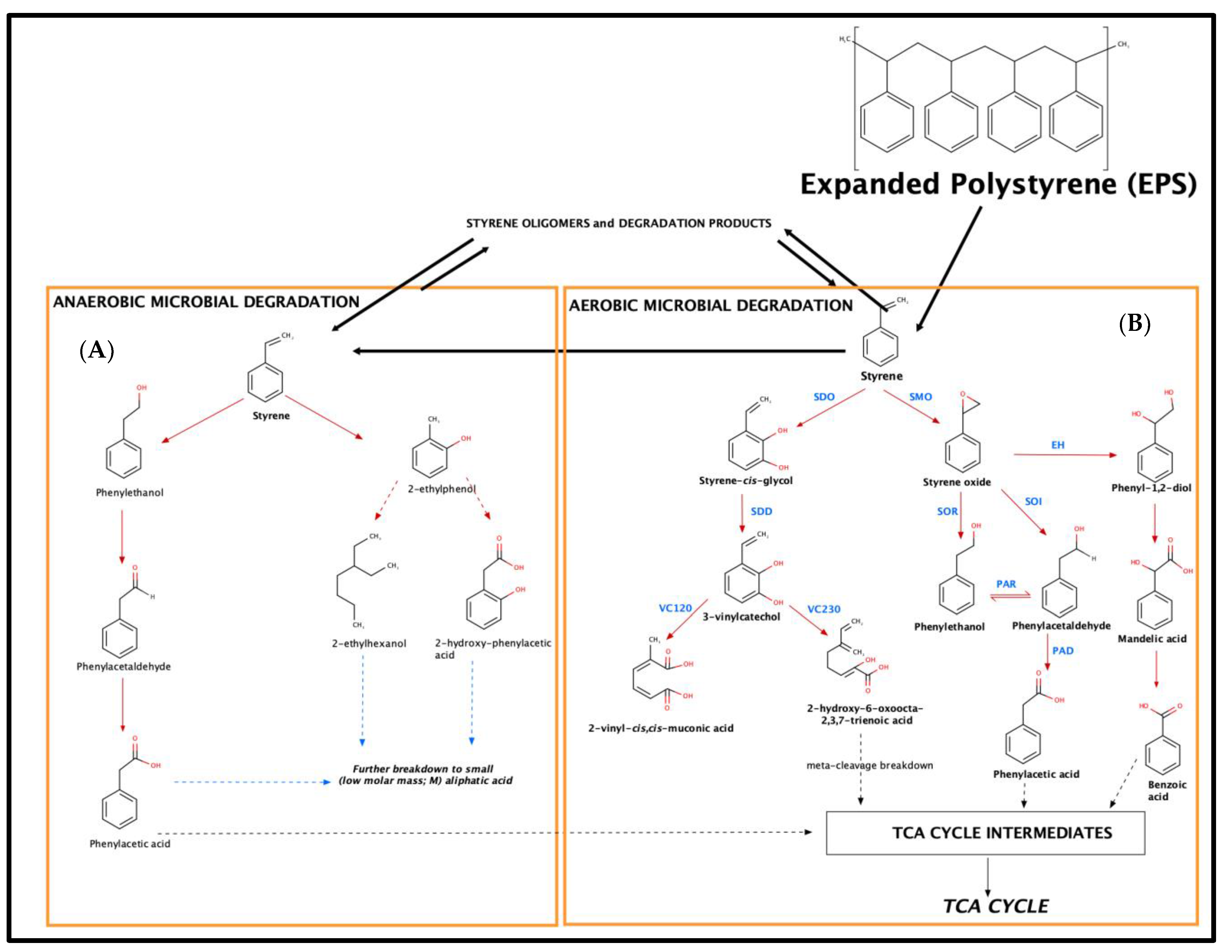
2.4. Metabolic Pathways for Styrene Biodegradation
3. Materials and Methods
3.1. Test Materials and Tenebrio molitor Larvae
3.2. Biodegradation of PS
3.3. Collection of Microflora
3.4. DNA Extraction
3.5. 16S-18S Assay: NGS-Based Detection and Differentiation of Nuclear Ribosomal Genes
3.6. Nitrogen/Protein Content in Frass and Biomass
3.7. Targeted Analysis of Monomers and Oligomers
3.8. Data Analysis and Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
Disclaimer
References
- Tsochatzis, E.D.; Gika, H.; Theodoridis, G. Development and Validation of a Fast Gas Chromatography Mass Spectrometry Method for the Quantification of Selected Non-Intentionally Added Substances and Polystyrene/Polyurethane Oligomers in Liquid Food Simulants. Anal. Chim. Acta 2020, 1130, 49–59. [Google Scholar] [CrossRef] [PubMed]
- International Life Sciences Institute (ILSI). Packaging Materials Vol. 2, Polystyrene for Food Packaging Applications: Report Prepared under the Responsibility of the ILSI Europe Packaging Material Task Force; ILSI Europe: Brussels, Belgium, 2002; ISBN 9781578811274. [Google Scholar]
- Lou, Y.; Ekaterina, P.; Yang, S.-S.; Lu, B.; Liu, B.; Ren, N.; Corvini, P.F.-X.; Xing, D. Biodegradation of Polyethylene and Polystyrene by Greater Wax Moth Larvae (Galleria Mellonella L.) and the Effect of Co-Diet Supplementation on the Core Gut Microbiome. Environ. Sci. Technol. 2020, 54, 2821–2831. [Google Scholar] [CrossRef] [PubMed]
- Tsochatzis, E.D.; Berggreen, I.E.; Nørgaard, J.V.; Theodoridis, G.; Dalsgaard, T.K. Biodegradation of Expanded Polystyrene by Mealworm Larvae under Different Feeding Strategies Evaluated by Metabolic Profiling Using GC-TOF-MS. Chemosphere 2021, 281, 130840. [Google Scholar] [CrossRef] [PubMed]
- Brandon, A.M.; Garcia, A.M.; Khlystov, N.A.; Wu, W.-M.; Criddle, C.S. Enhanced Bioavailability and Microbial Biodegradation of Polystyrene in an Enrichment Derived from the Gut Microbiome of Tenebrio Molitor (Mealworm Larvae). Environ. Sci. Technol. 2021, 55, 2027–2036. [Google Scholar] [CrossRef]
- Brandon, A.M.; Gao, S.-H.; Tian, R.; Ning, D.; Yang, S.-S.; Zhou, J.; Wu, W.-M.; Criddle, C.S. Biodegradation of Polyethylene and Plastic Mixtures in Mealworms (Larvae of Tenebrio Molitor ) and Effects on the Gut Microbiome. Environ. Sci. Technol. 2018, 52, 6526–6533. [Google Scholar] [CrossRef]
- Tsochatzis, E.; Alberto Lopes, J.F. Polystyrene Biodegradation by Tenebrio Molitor Larvae: Identification and Investigation by GC-MS Based Untargeted Screening. Polymers 2020, 13, 17. [Google Scholar] [CrossRef]
- Yang, S.-S.; Wu, W.-M.; Brandon, A.M.; Fan, H.-Q.; Receveur, J.P.; Li, Y.; Wang, Z.-Y.; Fan, R.; McClellan, R.L.; Gao, S.-H.; et al. Ubiquity of Polystyrene Digestion and Biodegradation within Yellow Mealworms, Larvae of Tenebrio Molitor Linnaeus (Coleoptera: Tenebrionidae). Chemosphere 2018, 212, 262–271. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Yang, J.; Wu, W.-M.; Zhao, J.; Song, Y.; Gao, L.; Yang, R.; Jiang, L. Biodegradation and Mineralization of Polystyrene by Plastic-Eating Mealworms: Part 1. Chemical and Physical Characterization and Isotopic Tests. Environ. Sci. Technol. 2015, 49, 12080–12086. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, J.; Wu, W.-M.; Zhao, J.; Song, Y.; Gao, L.; Yang, R.; Jiang, L. Biodegradation and Mineralization of Polystyrene by Plastic-Eating Mealworms: Part 2. Role of Gut Microorganisms. Environ. Sci. Technol. 2015, 49, 12087–12093. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, J.; Xia, M. Biodegradation and Mineralization of Polystyrene by Plastic-Eating Superworms Zophobas Atratus. Sci. Total Environ. 2020, 708, 135233. [Google Scholar] [CrossRef] [PubMed]
- Bombelli, P.; Howe, C.J.; Bertocchini, F. Polyethylene Bio-Degradation by Caterpillars of the Wax Moth Galleria Mellonella. Curr. Biol. 2017, 27, R292–R293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kundungal, H.; Gangarapu, M.; Sarangapani, S.; Patchaiyappan, A.; Devipriya, S.P. Efficient Biodegradation of Polyethylene (HDPE) Waste by the Plastic-Eating Lesser Waxworm (Achroia Grisella). Env. Sci Pollut Res 2019, 26, 18509–18519. [Google Scholar] [CrossRef]
- Kong, H.G.; Kim, H.H.; Chung, J.; Jun, J.; Lee, S.; Kim, H.-M.; Jeon, S.; Park, S.G.; Bhak, J.; Ryu, C.-M. The Galleria Mellonella Hologenome Supports Microbiota-Independent Metabolism of Long-Chain Hydrocarbon Beeswax. Cell Rep. 2019, 26, 2451–2464.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara, R.; Hirai, K.; Suzuki, S.; Kino, K. A Chemoenzymatic Process for Amide Bond Formation by an Adenylating Enzyme-Mediated Mechanism. Sci. Rep. 2018, 8, 2950. [Google Scholar] [CrossRef] [Green Version]
- Przemieniecki, S.W.; Kosewska, A.; Ciesielski, S.; Kosewska, O. Changes in the Gut Microbiome and Enzymatic Profile of Tenebrio Molitor Larvae Biodegrading Cellulose, Polyethylene and Polystyrene Waste. Environ. Pollut. 2020, 256, 113265. [Google Scholar] [CrossRef] [PubMed]
- Stoops, J.; Crauwels, S.; Waud, M.; Claes, J.; Lievens, B.; Van Campenhout, L. Microbial Community Assessment of Mealworm Larvae ( Tenebrio Molitor ) and Grasshoppers (Locusta Migratoria Migratorioides) Sold for Human Consumption. Food Microbiol. 2016, 53, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Urbanek, A.K.; Rybak, J.; Wróbel, M.; Leluk, K.; Mirończuk, A.M. A Comprehensive Assessment of Microbiome Diversity in Tenebrio Molitor Fed with Polystyrene Waste. Environ. Pollut. 2020, 262, 114281. [Google Scholar] [CrossRef]
- Bae, J.; woo Cho, H.; Jung, H.; Park, J.; Yun, S.; Ha, S.; Lee, Y.; Kim, T.-J. Changes in Intestinal Microbiota Due to the Expanded Polystyrene Diet of Mealworms (Tenebrio Molitor). Indian J. Microbiol. 2021, 61, 130–136. [Google Scholar] [CrossRef]
- Singh, S.N.; Kumari, B.; Mishra, S. Microbial Degradation of Xenobiotics; Singh, S.N., Ed.; Environmental Science and Engineering; Springer: Berlin/Heidelberg, Germany, 2012; ISBN 9783642237881. [Google Scholar]
- Tischler, D. Microbial Styrene Degradation; Springer International Publishing: Cham, Switzerland, 2015; ISBN 9783319248608. [Google Scholar]
- Ho, B.T.; Roberts, T.K.; Lucas, S. An Overview on Biodegradation of Polystyrene and Modified Polystyrene: The Microbial Approach. Crit. Rev. Biotechnol. 2018, 38, 308–320. [Google Scholar] [CrossRef]
- Oelschlägel, M.; Zimmerling, J.; Tischler, D. A Review: The Styrene Metabolizing Cascade of Side-Chain Oxygenation as Biotechnological Basis to Gain Various Valuable Compounds. Front. Microbiol. 2018, 9, 490. [Google Scholar] [CrossRef]
- Sowmya, H.V.; Ramalingappa; Krishnappa, M.; Thippeswamy, B. Degradation of Polyethylene by Penicillium Simplicissimum Isolated from Local Dumpsite of Shivamogga District. Env. Dev. Sustain. 2015, 17, 731–745. [Google Scholar] [CrossRef]
- Zhao, J.-H.; Wang, X.-Q.; Zeng, J.; Yang, G.; Shi, F.-H.; Yan, Q. Biodegradation of Poly(Butylene Succinate-Co-Butylene Adipate) by Aspergillus Versicolor. Polym. Degrad. Stab. 2005, 90, 173–179. [Google Scholar] [CrossRef]
- Valeriano, V.D.V.; Balolong, M.P.; Kang, D.-K. Probiotic Roles of Lactobacillus Sp. in Swine: Insights from Gut Microbiota. J. Appl. Microbiol. 2017, 122, 554–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Commission. Commission Regulation (EU) No 10/2011; European Commission: Brussels, Belgium, 2011. [Google Scholar]
- Guo, P.; Zhang, K.; Ma, X.; He, P. Clostridium Species as Probiotics: Potentials and Challenges. J. Anim. Sci. Biotechnol. 2020, 11, 24. [Google Scholar] [CrossRef] [PubMed]
- Sin, O.; Nollen, E.A.A. Regulation of Protein Homeostasis in Neurodegenerative Diseases: The Role of Coding and Non-Coding Genes. Cell. Mol. Life Sci. 2015, 72, 4027–4047. [Google Scholar] [CrossRef] [Green Version]
- Wawrzyńska, A.; Sirko, A. The Role of Selective Protein Degradation in the Regulation of Iron and Sulfur Homeostasis in Plants. IJMS 2020, 21, 2771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallela, A.; Nariya, M.K.; Deeds, E.J. Crosstalk and Ultrasensitivity in Protein Degradation Pathways. PLoS Comput. Biol. 2020, 16, e1008492. [Google Scholar] [CrossRef] [PubMed]
- Urs, K.C.D.; Hopkins, T.L. Effect of Moisture on Growth Rate and Development of Two Strains of Tenebrio Molitor L. (Coleoptera, Tenebrionidae). J. Stored Prod. Res. 1973, 8, 291–297. [Google Scholar] [CrossRef]
- Ring, M.; Pfeifer, T.A.; Grigliatti, T.A. Identification of a 5’ Truncated Non-LTR- Retrotransposon, YAKPsl, from the Variegated Cutworm, Peridroma Saucia, Using PCR. Insect Biochem. Molec. Biol. 1996, 26, 511–518. [Google Scholar] [CrossRef]
- Yu, Y.; Lee, C.; Kim, J.; Hwang, S. Group-Specific Primer and Probe Sets to Detect Methanogenic Communities Using Quantitative Real-Time Polymerase Chain Reaction. Biotechnol. Bioeng. 2005, 89, 670–679. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Family | Genus | Species | Control | Diet 1 (Bran: PS 4:1) | Diet 2 (Bran:PS 20:1 + H2O) | Diet 3 (Bran:PS 20:1) |
|---|---|---|---|---|---|---|
| Spiroplasmataceae | Spiroplasma | sp. | 68,567 | 55,880 | 66,520 | 77,787 |
| Enterobacteriaceae | Klebsiella | pneumoniae | 31,778 | 12,065 | 3602 | 877 |
| Enterobacteriaceae | Erwinia | oleae | 2038 | 10,977 | 8787 | 3485 |
| Enterococcaceae | Enterococcus | faecium | 602 | 0 | 4689 | 474 |
| Streptococcaceae | Lactococcus | lactis | 217 | 425 | 1542 | 253 |
| Streptococcaceae | Lactococcus | garvieae | 114 | 348 | 390 | 449 |
| Leuconostocaceae | Weissella | cibaria | 210 | 7 | 986 | 25 |
| Enterococcaceae | Enterococcus | mundtii | 0 | 968 | 0 | 0 |
| Clostridiaceae-1 | Clostridium | sp. | 198 | 193 | 103 | 389 |
| Flavobacteriaceae | Chryseobacterium | sp. | 122 | 157 | 88 | 201 |
| Pseudomonadaceae | Pseudomonas | sp. | 19 | 53 | 0 | 45 |
| Ruminococcaceae | Faecalibacterium | prausnitzii | 13 | 19 | 0 | 6 |
| Enterobacteriaceae | Proteus | sp. | 21 | 5 | 8 | 0 |
| Xanthomonadaceae | Stenotrophomonas | Rhizophila | 0 | 14 | 0 | 14 |
| Total n of reads | 103,953 | 81,188 | 86,776 | 84,133 | ||
| Family | Genus | Species | Control | Diet 1 (Bran:PS 4:1) | Diet 2 (Bran:PS 20:1 + H2O) | Diet 3 (Bran:PS 20:1) |
|---|---|---|---|---|---|---|
| Sporidiobolaceae | Sporobolomyces | roseus | 0 | 0 | 0 | 36 |
| Pleosporaceae | Alternaria | alternata | 0 | 0 | 0 | 27 |
| Trichocomaceae | Aspergillus | versicolor | 0 | 0 | 0 | 27 |
| Pleosporaceae | Pithomyces | chartarum | 16 | 0 | 0 | 0 |
| Tremellaceae | Cryptococcus | carnescens | 5 | 0 | 0 | 10 |
| Trichocomaceae | Aspergillus | sp. | 0 | 13 | 0 | 0 |
| Davidiellaceae | Cladosporium | sp. | 0 | 0 | 0 | 9 |
| Davidiellaceae | Cladosporium | herbarum | 0 | 8 | 0 | 0 |
| Malasseziaceae | Malassezia | sp. | 0 | 0 | 8 | 0 |
| Pleosporaceae | unclassified (F: Pleosporaceae) | sp. | 0 | 6 | 0 | 0 |
| Trichocomaceae | unclassified (F: Trichocomaceae) | sp. | 0 | 6 | 0 | 0 |
| unclassified (Dothideomycetes) | unclassified (C:Dothideomycetes) | sp. | 5 | 0 | 0 | 0 |
| Saccharomycetaceae | Candida | albicans | 0 | 0 | 0 | 5 |
| Total n of reads | 26 | 33 | 8 | 114 | ||
| Target Analytes | Average Mass Fractions in Frass (μg/mg) (±SD) | |||
|---|---|---|---|---|
| Control | Diet 1 (Bran:PS 4:1) | Diet 2 (Bran:PS 20:1 + H2O) | Diet 3 (Bran:PS 20:1) | |
| 1,1-diphenyl ethylene | 0 | 0.016 (±0.002) | 0.008 (±0.001) | 0.007 (±0.002) |
| Styrene | 0 | 0.005 (±0.001) | 0.003 (±0.001) | 0.002 (±0.001) |
| 2,4-diphenyl-1-butene | 0 | 0.014 (±0.001) | 0.004 (±0.001) | 0.004 (±0.001) |
| trans-1,2-diphenyl cyclobutane | 0 | 0.013 (±0.002) | 0.005 (±0.002) | 0.003 (±0.001) |
| 1e-tetrealin * | 0 | 0.006 (±0.001) | 0.004 (±0.001) | 0.005 (±0.001) |
| 1a-tetralin ** | 0 | 0.014 (±0.002) | 0.006 (±0.001) | 0.005 (±0.001) |
| Acetophenone | 0 | 0.180 (±0.020) | 0.070 (±0.014) | 0.050 (±0.012) |
| 2,4-di-tert butyl phenol | 0 | 0.100 (±0.010) | 0.050 (±0.011) | 0.040 (±0.010) |
| 2,4,6-triphenyl-1-hexane | 0 | 0.173 (±0.019) | 0.051 (±0.013) | 0.030 (±0.011) |
| Component | Protein (%) | Starch (%) | Fat Content (%) | Soluble Fibers (%) | Insoluble Fibers (%) | Ash Content (%) |
|---|---|---|---|---|---|---|
| Amount | 14.7 | 22.0 | 4.5 | 2.8 | 43.2 | 5.6 |
| Feeding Diets | ||||
|---|---|---|---|---|
| Control | Diet 1 | Diet 2 | Diet 3 | |
| Rolled barley (g) * | 20.0 ± 0.01 | 3.99 ± 0.06 | 20.1 ± 0.03 | 20.1 ± 0.03 |
| Polystyrene (PS) (mg) | 0.0 | 1000 ± 10 | 980 ± 16 | 1015 ± 17 |
| Water (mL) ** | 0 | 0 | 1.5 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsochatzis, E.; Berggreen, I.E.; Tedeschi, F.; Ntrallou, K.; Gika, H.; Corredig, M. Gut Microbiome and Degradation Product Formation during Biodegradation of Expanded Polystyrene by Mealworm Larvae under Different Feeding Strategies. Molecules 2021, 26, 7568. https://doi.org/10.3390/molecules26247568
Tsochatzis E, Berggreen IE, Tedeschi F, Ntrallou K, Gika H, Corredig M. Gut Microbiome and Degradation Product Formation during Biodegradation of Expanded Polystyrene by Mealworm Larvae under Different Feeding Strategies. Molecules. 2021; 26(24):7568. https://doi.org/10.3390/molecules26247568
Chicago/Turabian StyleTsochatzis, Emmanouil, Ida Elizabeth Berggreen, Francesca Tedeschi, Konstantina Ntrallou, Helen Gika, and Milena Corredig. 2021. "Gut Microbiome and Degradation Product Formation during Biodegradation of Expanded Polystyrene by Mealworm Larvae under Different Feeding Strategies" Molecules 26, no. 24: 7568. https://doi.org/10.3390/molecules26247568
APA StyleTsochatzis, E., Berggreen, I. E., Tedeschi, F., Ntrallou, K., Gika, H., & Corredig, M. (2021). Gut Microbiome and Degradation Product Formation during Biodegradation of Expanded Polystyrene by Mealworm Larvae under Different Feeding Strategies. Molecules, 26(24), 7568. https://doi.org/10.3390/molecules26247568