
Screening the Key Region of Sunlight Regulating the Flavonoid Profiles of Young Shoots in Tea Plants (Camellia sinensis L.) Based on a Field Experiment


Abstract
:1. Introduction
2. Results
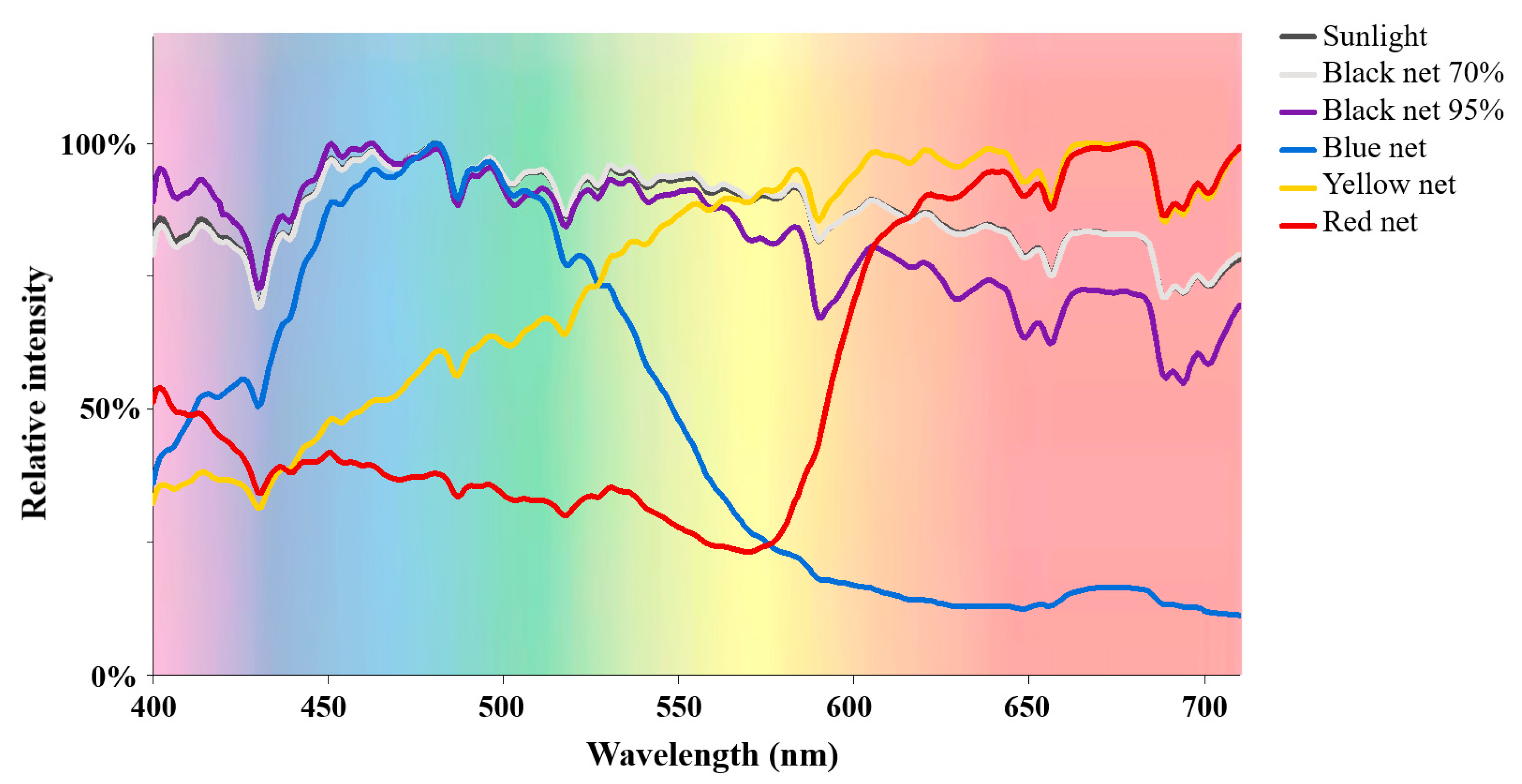
2.1. Alteration Effects of Different Shade Treatments on the Light Condition Underneath
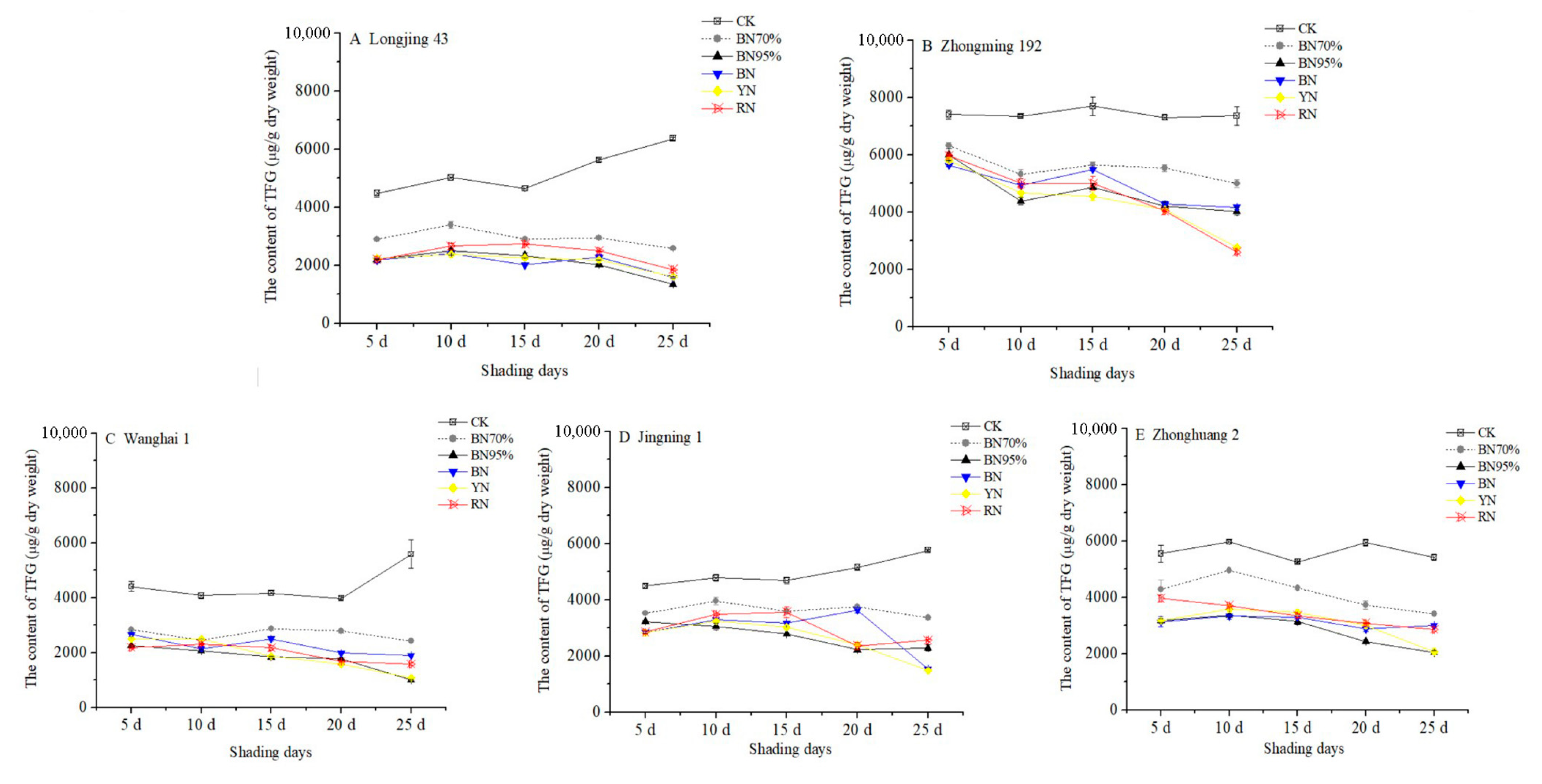
2.2. Impacts of Shade Treatments on the Profiles of Catechins in the Young Shoots of Different Tea Cultivars
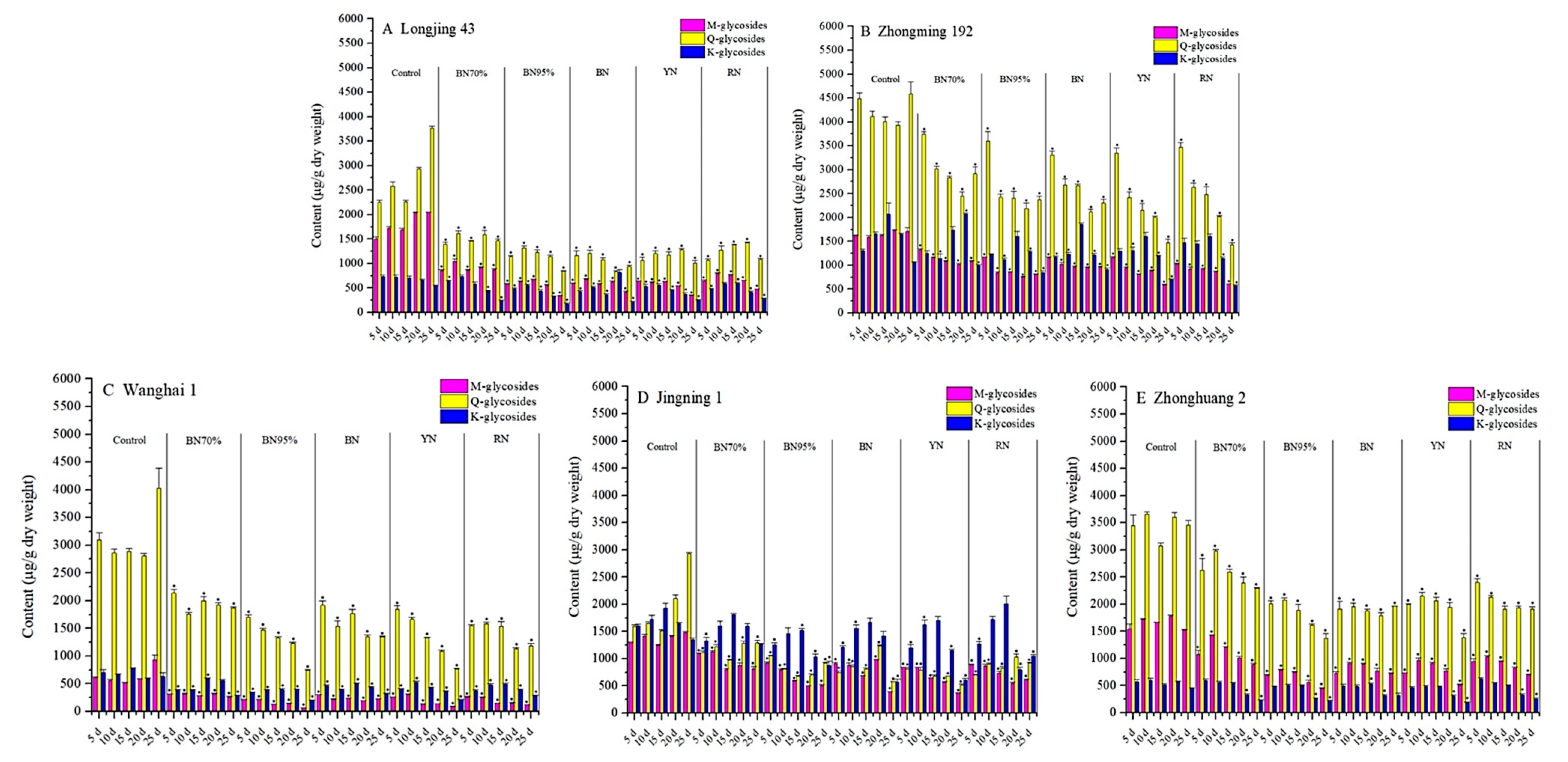
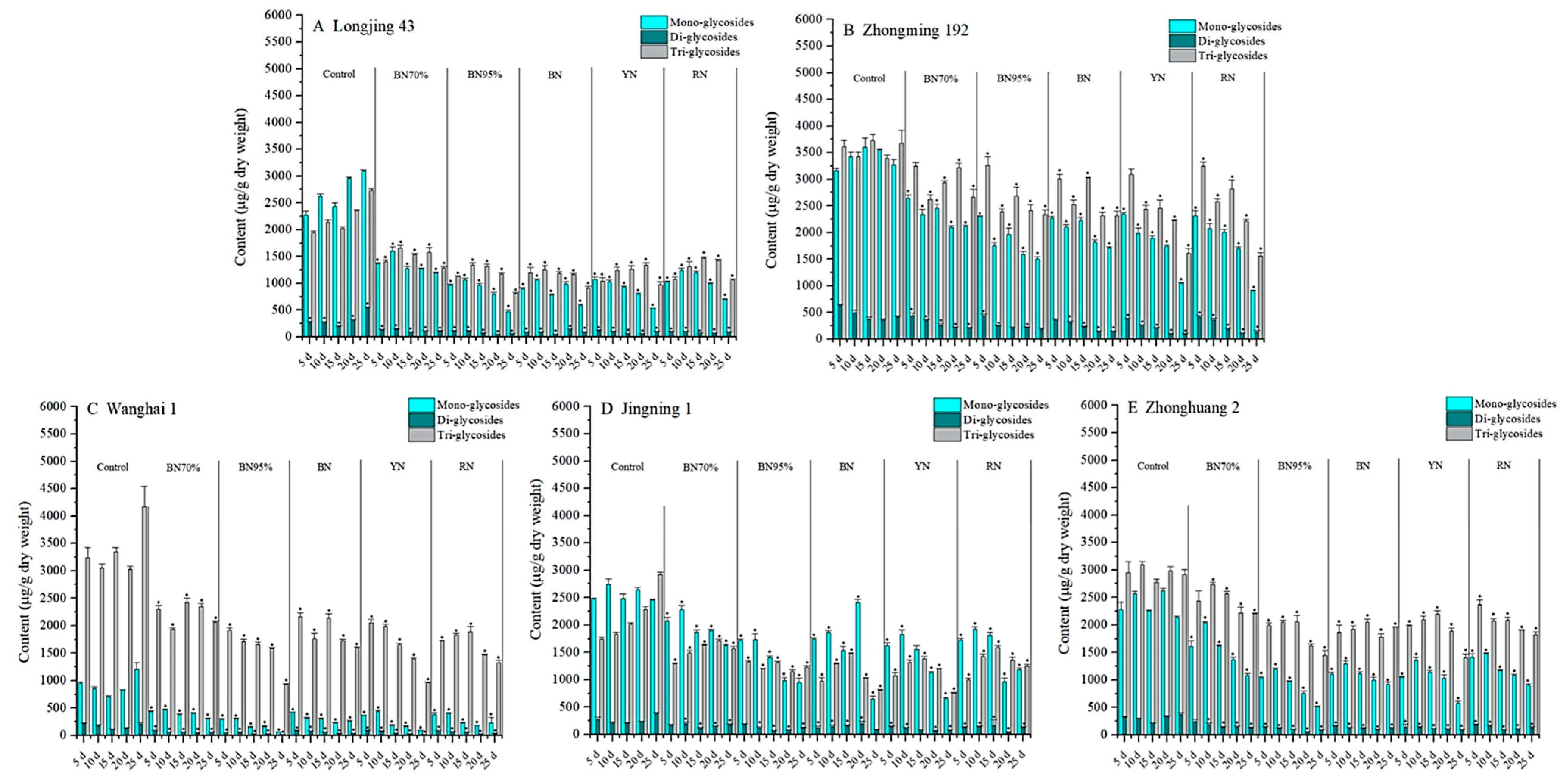
2.3. Impacts of Shade Treatments on the Profiles of Flavonol Glycosides in the Young Shoots of Different Tea Cultivars
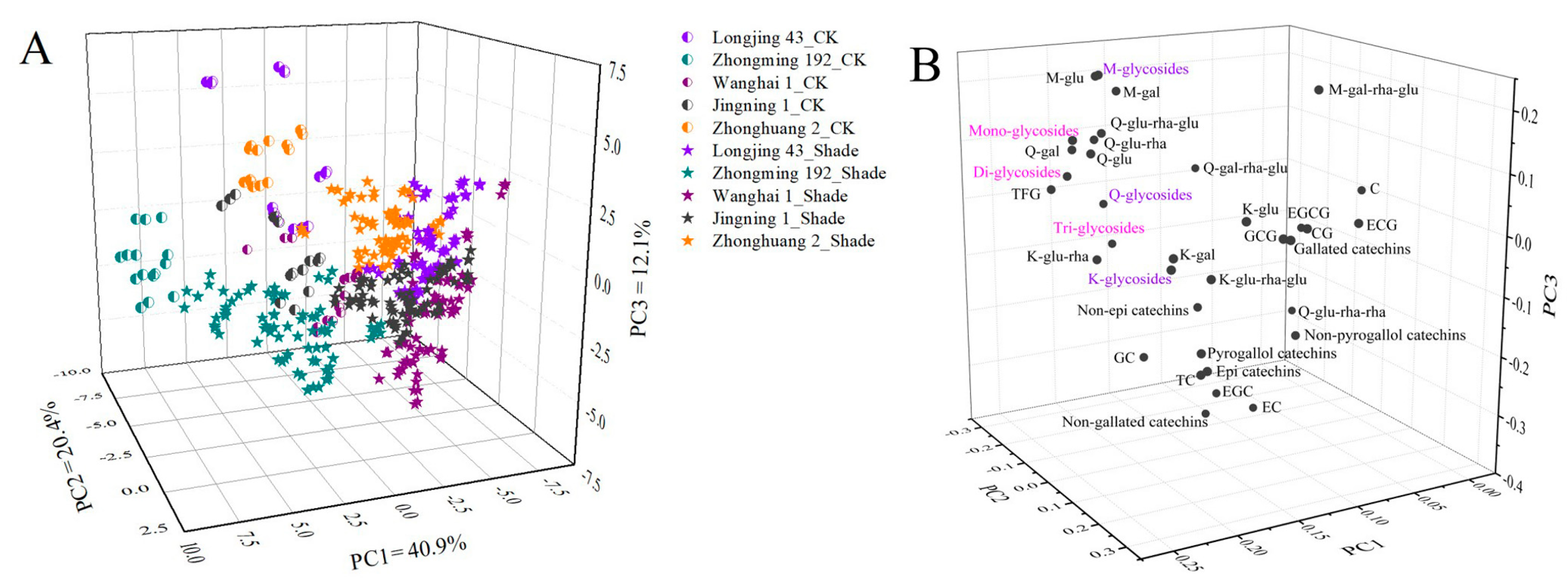
2.4. Screening the Key Factor of the Profiles of Flavonoids Based on PCA
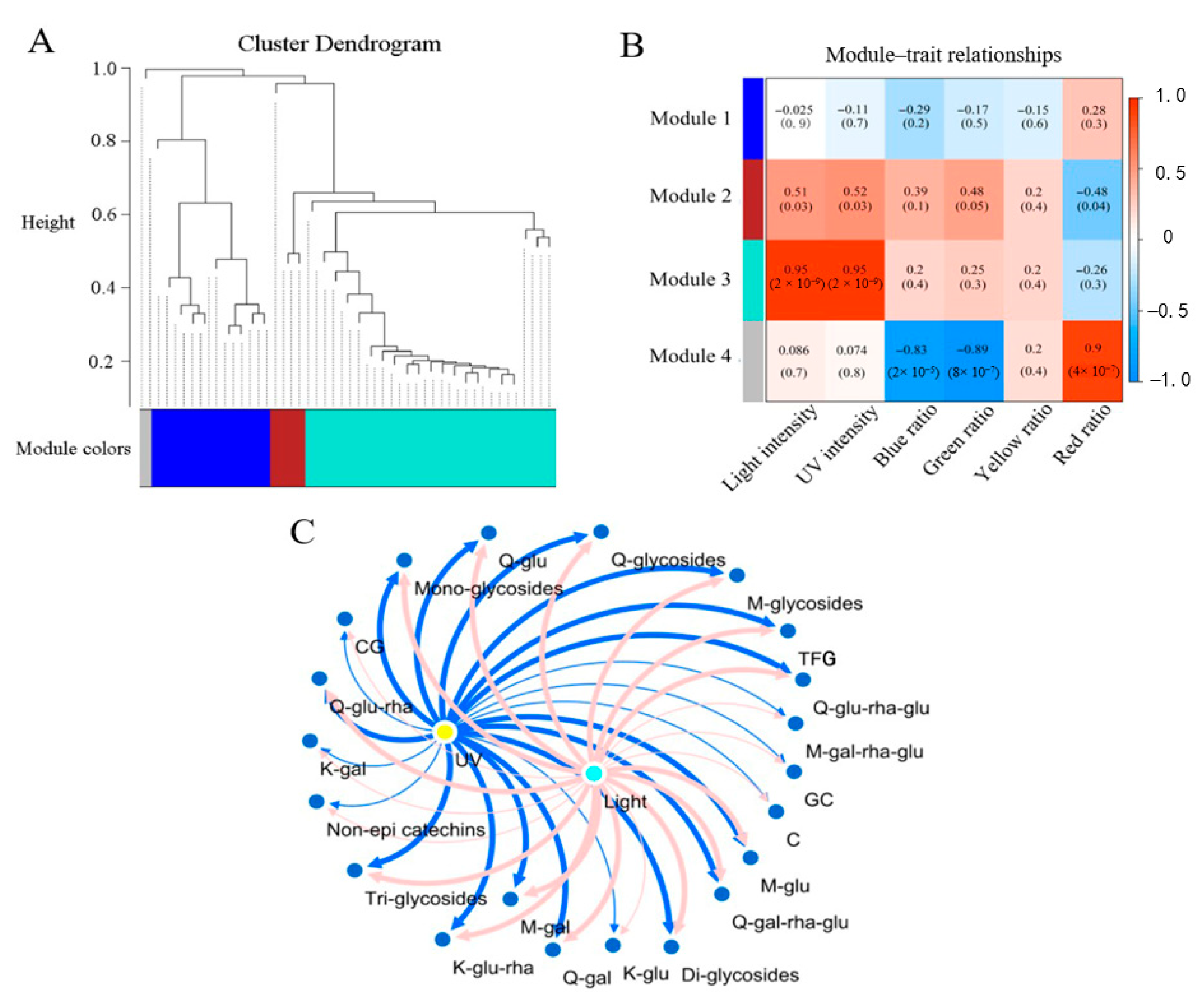
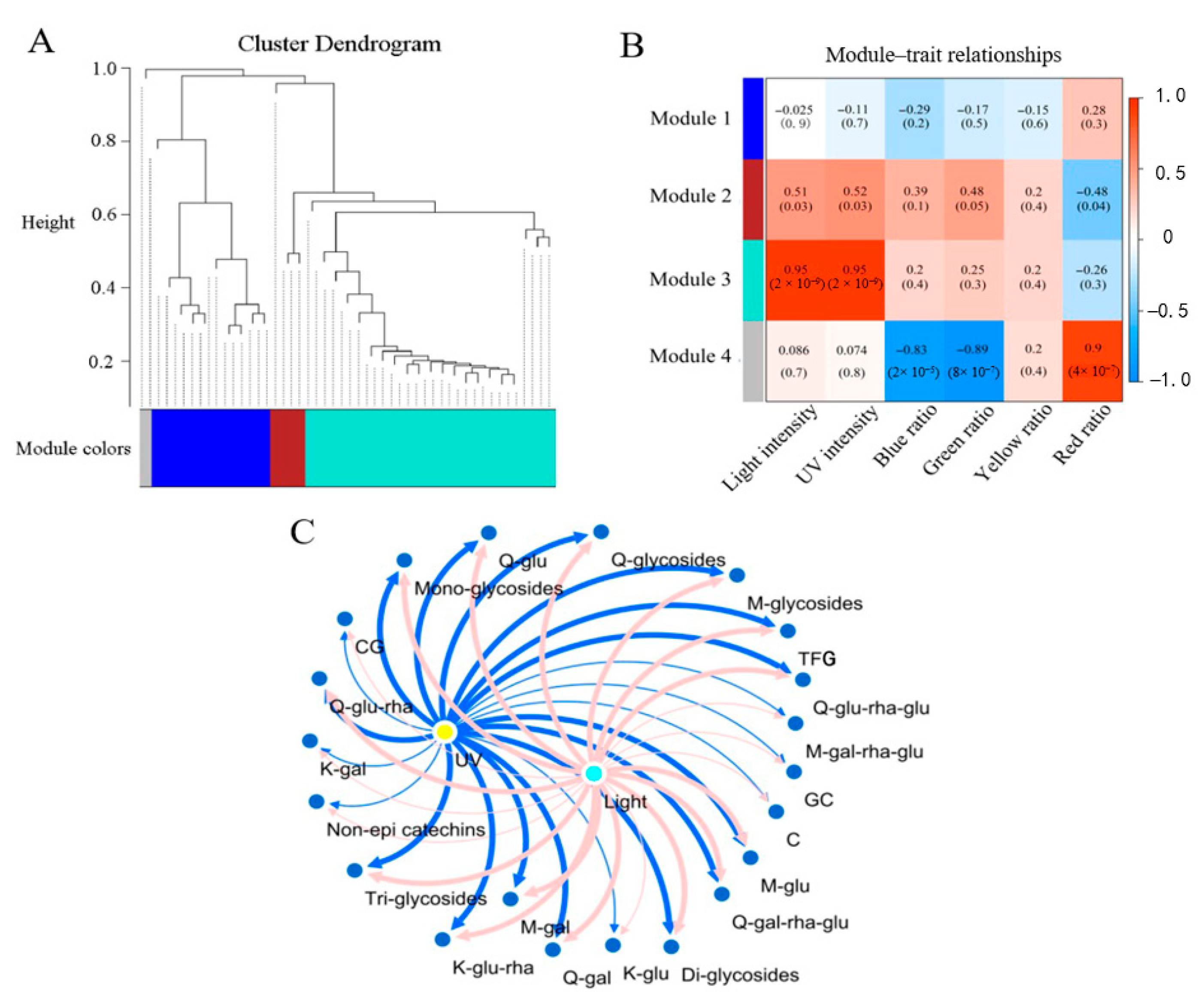
2.5. Association of Flavonoid Compositions with the Light Condition Parameters
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Implementation of Shade Treatments
4.3. Preparation of Fixed Tea Samples
4.4. Analysis of Catechins and Flavonol Glycosides in Tea Samples
4.5. Association of Flavonoid Compounds with the Light Condition Parameters
4.6. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Sample Availability
References
- Pinto, M.D. Tea: A new perspective on health benefits. Food Res. Int. 2013, 53, 558–567. [Google Scholar] [CrossRef]
- Khan, N.; Mukhtar, H. Tea polyphenols in promotion of human health. Nutrients 2019, 11, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodgson, J.M.; Croft, K.D. Tea flavonoids and cardiovascular health. Mol. Asp. Med. 2010, 31, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.Q.; Nie, Y.; Gao, Y.; Huang, B.; Ye, J.H.; Lu, J.L.; Liang, Y.R. Screening the cultivar and processing factors based on the flavonoid profiles of dry teas using principal component analysis. J. Food Compos. Anal. 2018, 67, 29–37. [Google Scholar] [CrossRef]
- Mamati, G.E.; Liang, Y.R.; Lu, J.L. Expression of basic genes involved in tea polyphenol synthesis in relation to accumulation of catechins and total tea polyphenols. J. Sci. Food Agric. 2006, 86, 459–464. [Google Scholar] [CrossRef]
- Fang, Z.T.; Lv, Y.Q.; Song, C.J.; Jin, J.; Lu, J.L.; Xu, H.R.; Ye, J.H. Simultaneous preparation of abundant flavonol triglycosides from Tea Leaves. Molecules 2020, 25, 5140. [Google Scholar] [CrossRef] [PubMed]
- Scharbert, S.; Hofmann, T. Molecular definition of black tea taste by means of quantitative studies, taste reconstitution, and omission experiments. J. Agric. Food Chem. 2005, 53, 5377–5384. [Google Scholar] [CrossRef]
- Treutter, D. Significance of flavonoids in plant resistance: A review. Environ. Chem. Lett. 2006, 4, 147–157. [Google Scholar] [CrossRef]
- Mouradov, A.; Spangenberg, G. Flavonoids: A metabolic network mediating plants adaptation to their real estate. Front. Plant. Sci. 2014, 5, 1–16. [Google Scholar] [CrossRef]
- Doerr, O.S.; Zimmermann, B.F.; Kogler, S.; Mibus, H. Influence of leaf temperature and blue light on the accumulation of rosmarinic acid and other phenolic compounds in Plectranthus scutellarioides (L.). Environ. Exp. Bot. 2019, 167, 103830. [Google Scholar] [CrossRef]
- Ryan, K.G.; Swinny, E.E.; Markham, K.R.; Winefield, C. Flavonoid gene expression and UV photoprotection in transgenic and mutant Petunia leaves. Phytochemistry 2002, 59, 23–32. [Google Scholar] [CrossRef]
- Wang, Y.S.; Gao, L.P.; Shan, Y.; Liu, Y.J.; Tian, Y.W.; Xia, T. Influence of shade on flavonoid biosynthesis in tea (Camellia sinensis (L.) O. Kuntze). Sci. Hortic-Amst. 2012, 141, 7–16. [Google Scholar] [CrossRef]
- Zoratti, L.; Karppinen, K.; Escobar, A.L.; Haggman, H.; Jaakola, L. Light-controlled flavonoid biosynthesis in fruits. Front. Plant Sci. 2014, 5, 534. [Google Scholar] [CrossRef]
- Wagner, J.R.; Brunzelle, J.S.; Forest, K.T.; Vierstra, R.D. A light-sensing knot revealed by the structure of the chromophore-binding domain of phytochrome. Nature 2005, 438, 325–331. [Google Scholar] [CrossRef]
- Costa, T.P.C.; Rodrigues, E.M.; Dias, L.P.; Pupin, B.; Ferreira, P.C.; Rangel, D.E.N. Different wavelengths of visible light influence the conidial production and tolerance to ultra-violet radiation of the plant pathogens Colletotrichum acutatum and Fusarium fujikuroi. Eur. J. Plant Pathol. 2021, 159, 105–115. [Google Scholar] [CrossRef]
- Lin, N.; Liu, X.Y.; Zhu, W.F.; Cheng, X.; Wang, X.H.; Wan, X.C.; Liu, L.L. Ambient Ultraviolet B signal modulates tea flavor characteristics via shifting a metabolic flux in flavonoid biosynthesis. J. Agric. Food Chem. 2021, 69, 3401–3414. [Google Scholar] [CrossRef]
- Park, Y.J.; Runkle, E.S. Spectral effects of light-emitting diodes on plant growth, visual color quality, and photosynthetic photon efficacy: White versus blue plus red radiation. PLoS ONE 2018, 13, e0202386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, C.; Ma, J.Q.; Ma, C.L.; Shen, S.Y.; Liu, Y.F.; Chen, L. Regulation of growth and flavonoid formation of tea plants (Camellia sinensis) by blue and green Light. J. Agric. Food Chem. 2019, 67, 2408–2419. [Google Scholar] [CrossRef]
- Wang, P.J.; Chen, S.R.; Gu, M.Y.; Chen, X.M.; Chen, X.J.; Yang, J.F.; Zhao, F.; Ye, N.X. Exploration of the effects of different blue LED light intensities on flavonoid and lipid metabolism in tea plants via transcriptomics and metabolomics. Int. J. Mol. Sci. 2020, 21, 4606. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.X.; Zhang, Y.C.; Yang, X.F.; Xiao, J.P.; Zhang, H.Q.; Zhang, Z.F.; Wang, Y.Z.; Jiang, G.H. Colored light-quality selective plastic films affect anthocyanin content, enzyme activities, and the expression of flavonoid genes in strawberry (Fragaria x ananassa) fruit. Food Chem. 2016, 207, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.-H.; Lv, Y.-Q.; Liu, S.-R.; Jin, J.; Wang, Y.-F.; Wei, C.-L.; Zhao, S.-Q. Effects of light intensity and spectral composition on the transcriptome profiles of leaves in shade-grown tea plants (Camellia sinensis L.) and regulatory network of flavonoid biosynthesis. Molecules 2021, 26, 5836. [Google Scholar] [CrossRef]
- Sano, S.; Takemoto, T.; Ogihara, A.; Suzuki, K.; Masumura, T.; Satoh, S.; Takano, K.; Mimura, Y.; Morita, S. Stress responses of shade-treated tea leaves to high light exposure after removal of shading. Plants 2020, 9, 302. [Google Scholar] [CrossRef] [Green Version]
- Gai, Z.S.; Wang, Y.; Ding, Y.Q.; Qian, W.J.; Qiu, C.; Xie, H.; Sun, L.T.; Jiang, Z.W.; Ma, Q.P.; Wang, L.J.; et al. Exogenous abscisic acid induces the lipid and flavonoid metabolism of tea plants under drought stress. Sci. Rep. 2020, 10, 12275. [Google Scholar] [CrossRef]
- Wang, Y.S.; Gao, L.P.; Wang, Z.R.; Liu, Y.J.; Sun, M.L.; Yang, D.Q.; Wei, C.L.; Shan, Y.; Xia, T. Light-induced expression of genes involved in phenylpropanoid biosynthetic pathways in callus of tea (Camellia sinensis (L.) O. Kuntze). Sci. Hortic-Amst. 2012, 133, 72–83. [Google Scholar] [CrossRef]
- Zhang, Q.; Shi, Y.; Ma, L.; Yi, X.; Ruan, J. Metabolomic analysis using ultra-performance liquid chromatography-quadrupole-time of flight mass spectrometry (UPLC-Q-TOF MS) uncovers the effects of light intensity and temperature under shading treatments on the metabolites in tea. PLoS ONE 2014, 9, e112572. [Google Scholar] [CrossRef]
- Neugart, S.; Krumbeinand, A.; Zrenner, R. Influence of light and temperature on gene expression leading to accumulation of specific flavonol glycosides and hydroxycinnamic acid derivatives in Kale (Brassica oleracea var. sabellica). Front. Plant Sci. 2016, 7, 326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, K.G.; Swinny, E.E.; Winefield, C.; Markham, K.R. Flavonoids and UV photoprotection in Arabidopsis mutants. Z. Fur Nat. Sect. C J. Biosci. 2001, 56, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Majer, P.; Neugart, S.; Krumbein, A.; Schreiner, M.; Hideg, E. Singlet oxygen scavenging by leaf flavonoids contributes to sunlight acclimation in Tilia platyphyllos. Environ. Exp. Bot. 2014, 100, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Su, X.J.; Wang, W.Z.; Xia, T.; Gao, L.P.; Shen, G.A.; Pang, Y.Z. Characterization of a heat responsive UDP: Flavonoid glucosyltransferase gene in tea plant (Camellia sinensis). PLoS ONE 2018, 13, e0207212. [Google Scholar] [CrossRef]
- Jones, P.; Messner, B.; Nakajima, J.I.; Schaffner, A.R.; Saito, K. UGT73C6 and UGT78D1, glycosyltransferases involved in flavonol glycoside biosynthesis in Arabidopsis thaliana. J. Biol. Chem. 2003, 278, 43910–43918. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Yue, C.; Cao, H.; Zhou, Y.; Zeng, J.; Yang, Y.; Wang, X. Biochemical and transcriptome analyses of a novel chlorophyll-deficient chlorina tea plant cultivar. BMC Plant Biol. 2014, 14, 352. [Google Scholar] [CrossRef] [Green Version]
- Li, N.N.; Lu, J.L.; Li, Q.S.; Zheng, X.Q.; Wang, X.C.; Wang, L.; Wang, Y.C.; Ding, C.Q.; Liang, Y.R.; Yang, Y.J. Dissection of chemical composition and associated gene expression in the pigment-deficient tea cultivar ‘Xiaoxueya’ reveals an albino phenotype and metabolite formation. Front. Plant Sci. 2019, 10, 1543. [Google Scholar] [CrossRef]
- Guo, Y.; Zhu, C.; Zhao, S.; Zhang, S.; Wang, W.; Fu, H.; Li, X.; Zhou, C.; Chen, L.; Lin, Y.; et al. De novo transcriptome and phytochemical analyses reveal differentially expressed genes and characteristic secondary metabolites in the original oolong tea (Camellia sinensis) cultivar ‘Tieguanyin’ compared with cultivar ‘Benshan’. BMC Genom. 2019, 20, 265. [Google Scholar] [CrossRef] [Green Version]
- Fang, Z.T.; Yang, W.T.; Li, C.Y.; Li, D.; Dong, J.J.; Zhao, D.; Xu, H.R.; Ye, J.H.; Zheng, X.Q.; Liang, Y.R.; et al. Accumulation pattern of catechins and flavonol glycosides in different varieties and cultivars of tea plant in China. J. Food Compos. Anal. 2021, 97, 103772. [Google Scholar] [CrossRef]
- Liu, G.F.; Han, Z.X.; Feng, L.; Gao, L.P.; Gao, M.J.; Gruber, M.Y.; Zhang, Z.L.; Xia, T.; Wan, X.C.; Wei, S. Metabolic flux redirection and transcriptomic reprogramming in the albino tea cultivar ‘Yu-Jin-Xiang’ with an emphasis on catechin Production. Sci. Rep. 2017, 7, 45062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.F.; Liu, M.Y.; Ruan, J.Y. Metabolomics analysis reveals the metabolic and functional roles of flavonoids in light-sensitive tea leaves. BMC Plant Biol. 2017, 17, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.F.; Zhao, H.; Guo, F.; Shi, X.P.; Ye, C.; Yang, P.X.; Liu, B.Y.; Ni, D.J. Transcriptomic analysis reveals mechanism of light-sensitive albinism in tea plant Camellia sinensis ‘Huangjinju’. BMC Plant Biol. 2020, 20, 216. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Su, H.; Jin, R.; Mao, Y.; Xu, A.; Cheng, H.; Wang, Y.; Meng, Q. Shading effects on leaf color conversion and biosynthesis of the major secondary metabolites in the albino tea cultivar “Yujinxiang”. J. Agric. Food Chem. 2020, 68, 2528–2538. [Google Scholar] [CrossRef]
- Zhao, X.C.; Zeng, X.S.; Lin, N.; Yu, S.W.; Fernie, A.R.; Zhao, J. CsbZIP1-CsMYB12 mediates the production of bitter-tasting flavonols in tea plants (Camellia sinensis) through a coordinated activator-repressor network. Hortic Res. 2021, 8, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Gam, D.T.; Khoi, P.H.; Ngoc, P.B.; Linh, L.K.; Hung, N.K.; Anh, P.T.L.; Thu, N.T.; Hien, N.T.T.; Khanh, T.D.; Ha, C.H. LED Lights Promote Growth and Flavonoid Accumulation of Anoectochilus roxburghii and are linked to the enhanced expression of several related genes. Plants 2020, 9, 1344. [Google Scholar] [CrossRef]
- Liu, L.L.; Li, Y.Y.; She, G.B.; Zhang, X.C.; Jordan, B.; Chen, Q.; Zhao, J.; Wan, X.C. Metabolite profiling and transcriptomic analyses reveal an essential role of UVR8-mediated signal transduction pathway in regulating flavonoid biosynthesis in tea plants (Camellia sinensis) in response to shading. BMC Plant Biol. 2018, 18, 233. [Google Scholar] [CrossRef]
- Yang, L.Y.; Wang, L.T.; Ma, J.H.; Ma, E.D.; Li, J.Y.; Gong, M. Effects of light quality on growth and development, photosynthetic characteristics and content of carbohydrates in tobacco (Nicotiana tabacum L.) plants. Photosynthetica 2017, 55, 467–477. [Google Scholar] [CrossRef]
- Li, H.S.; Lin, Y.L.; Chen, X.H.; Bai, Y.; Wang, C.Q.; Xu, X.P.; Wang, Y.; Lai, Z.X. Effects of blue light on flavonoid accumulation linked to the expression of miR393, miR394 and miR395 in longan embryogenic calli. PLoS ONE 2018, 13, e0191444. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, D.T.P.; Kitayama, M.; Lu, N.; Takagaki, M. Improving secondary metabolite accumulation, mineral content, and growth of coriander (Coriandrum sativum L.) by regulating light quality in a plant factory. J. Hortic. Sci. Biotech. 2020, 95, 356–363. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sunlight | Black Net 70% | Black Net 95% | Blue Net | Yellow Net | Red Net | |
|---|---|---|---|---|---|---|
| Light intensity (lux) | 12,686 ± 407 | 5165 ± 729 | 892 ± 300 | 1713 ± 357 | 1945 ± 196 | 2938 ± 272 |
| UV intensity (µw/cm2) | 4823 ± 258 | 1765 ± 120 | 211 ± 89 | 185 ± 45 | 202 ± 110 | 640 ± 54 |
| Blue ratio in the light spectrum (%) | 31.7 ± 0.3 | 31.5 ± 0.1 | 34.5 ± 0.3 | 50.4 ± 0.9 | 18.8 ± 0.2 | 22.5 ± 0.2 |
| Green ratio in the light spectrum (%) | 25.8 ± 0.1 | 25.7 ± 0.1 | 26.6 ± 0.3 | 34.5 ± 0.3 | 24.6 ± 0.1 | 13.2 ± 0.3 |
| Yellow ratio in the light spectrum (%) | 6.4 ± 0.1 | 6.4 ± 0.1 | 6.1 ± 0.1 | 3.1 ± 0.1 | 7.5 ± 0.1 | 3.3 ± 0.1 |
| Red ratio in the light spectrum (%) | 36.2 ± 0.3 | 36.4 ± 0.1 | 32.8 ± 0.1 | 12.0 ± 0.5 | 49.1 ± 0.3 | 61.0 ± 0.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, J.; Lv, Y.-Q.; He, W.-Z.; Li, D.; Ye, Y.; Shu, Z.-F.; Shao, J.-N.; Zhou, J.-H.; Chen, D.-M.; Li, Q.-S.; et al. Screening the Key Region of Sunlight Regulating the Flavonoid Profiles of Young Shoots in Tea Plants (Camellia sinensis L.) Based on a Field Experiment. Molecules 2021, 26, 7158. https://doi.org/10.3390/molecules26237158
Jin J, Lv Y-Q, He W-Z, Li D, Ye Y, Shu Z-F, Shao J-N, Zhou J-H, Chen D-M, Li Q-S, et al. Screening the Key Region of Sunlight Regulating the Flavonoid Profiles of Young Shoots in Tea Plants (Camellia sinensis L.) Based on a Field Experiment. Molecules. 2021; 26(23):7158. https://doi.org/10.3390/molecules26237158
Chicago/Turabian StyleJin, Jing, Yi-Qing Lv, Wei-Zhong He, Da Li, Ying Ye, Zai-Fa Shu, Jing-Na Shao, Jia-Hao Zhou, Ding-Mi Chen, Qing-Sheng Li, and et al. 2021. "Screening the Key Region of Sunlight Regulating the Flavonoid Profiles of Young Shoots in Tea Plants (Camellia sinensis L.) Based on a Field Experiment" Molecules 26, no. 23: 7158. https://doi.org/10.3390/molecules26237158
APA StyleJin, J., Lv, Y.-Q., He, W.-Z., Li, D., Ye, Y., Shu, Z.-F., Shao, J.-N., Zhou, J.-H., Chen, D.-M., Li, Q.-S., & Ye, J.-H. (2021). Screening the Key Region of Sunlight Regulating the Flavonoid Profiles of Young Shoots in Tea Plants (Camellia sinensis L.) Based on a Field Experiment. Molecules, 26(23), 7158. https://doi.org/10.3390/molecules26237158