
Cynanchum auriculatum Royle ex Wight., Cynanchum bungei Decne. and Cynanchum wilfordii (Maxim.) Hemsl.: Current Research and Prospects

Abstract
1. Introduction
2. Botany
2.1. Morphological Characteristics
2.2. Identification of CA, CB and CW
2.3. Relationship of CA, CB and CW
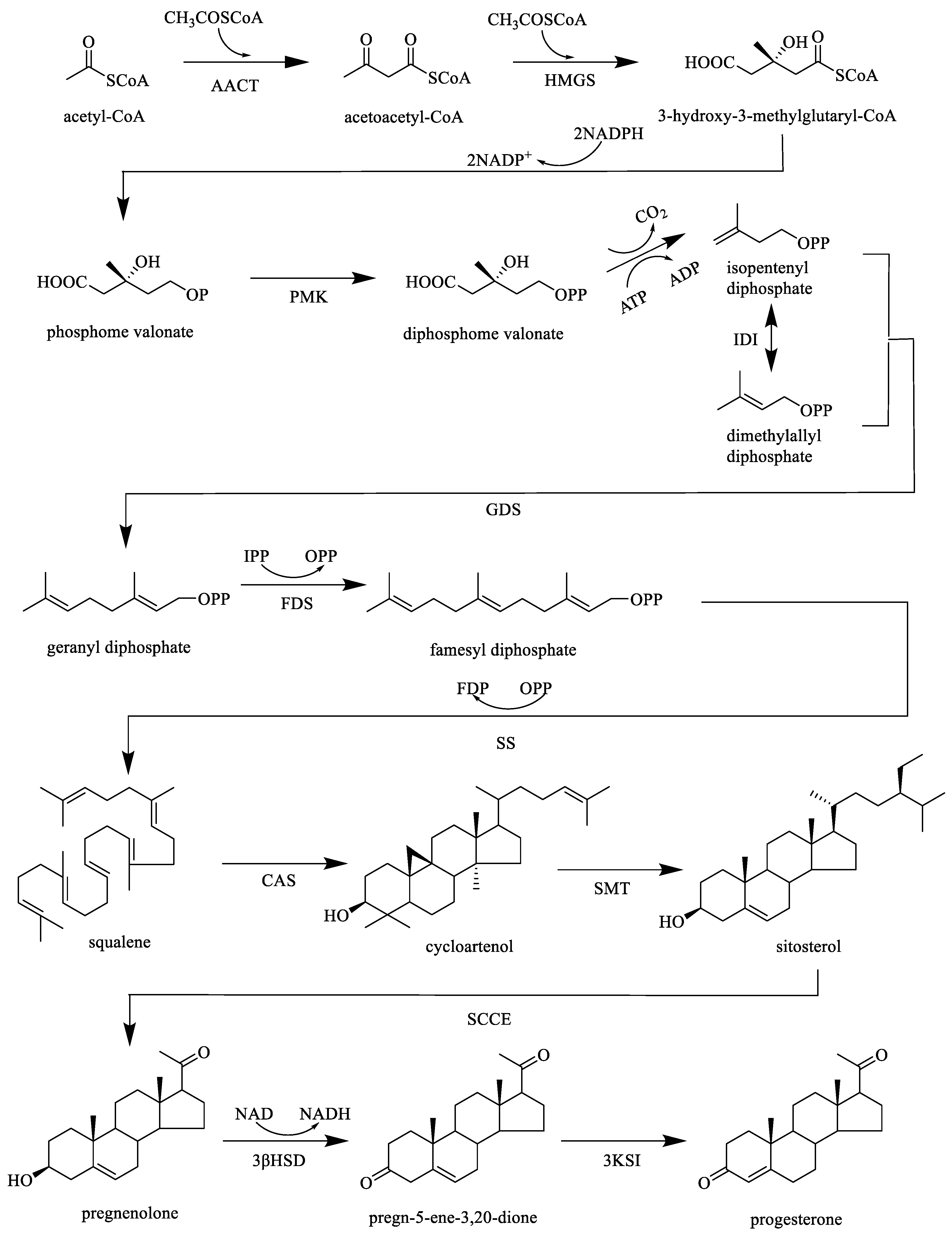
3. Phytochemistry
3.1. Extraction and Separation
3.2. Chemical Constituents
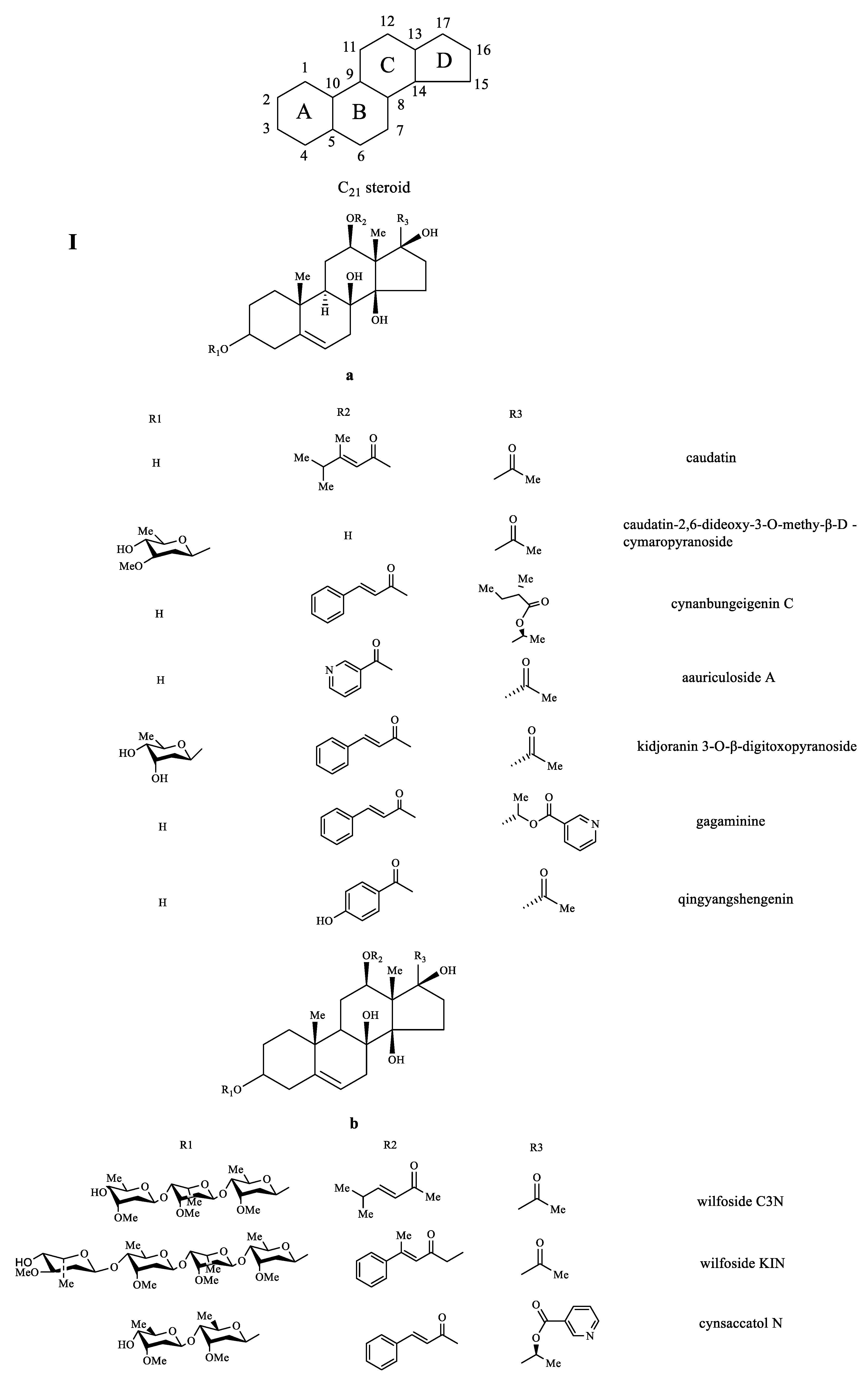
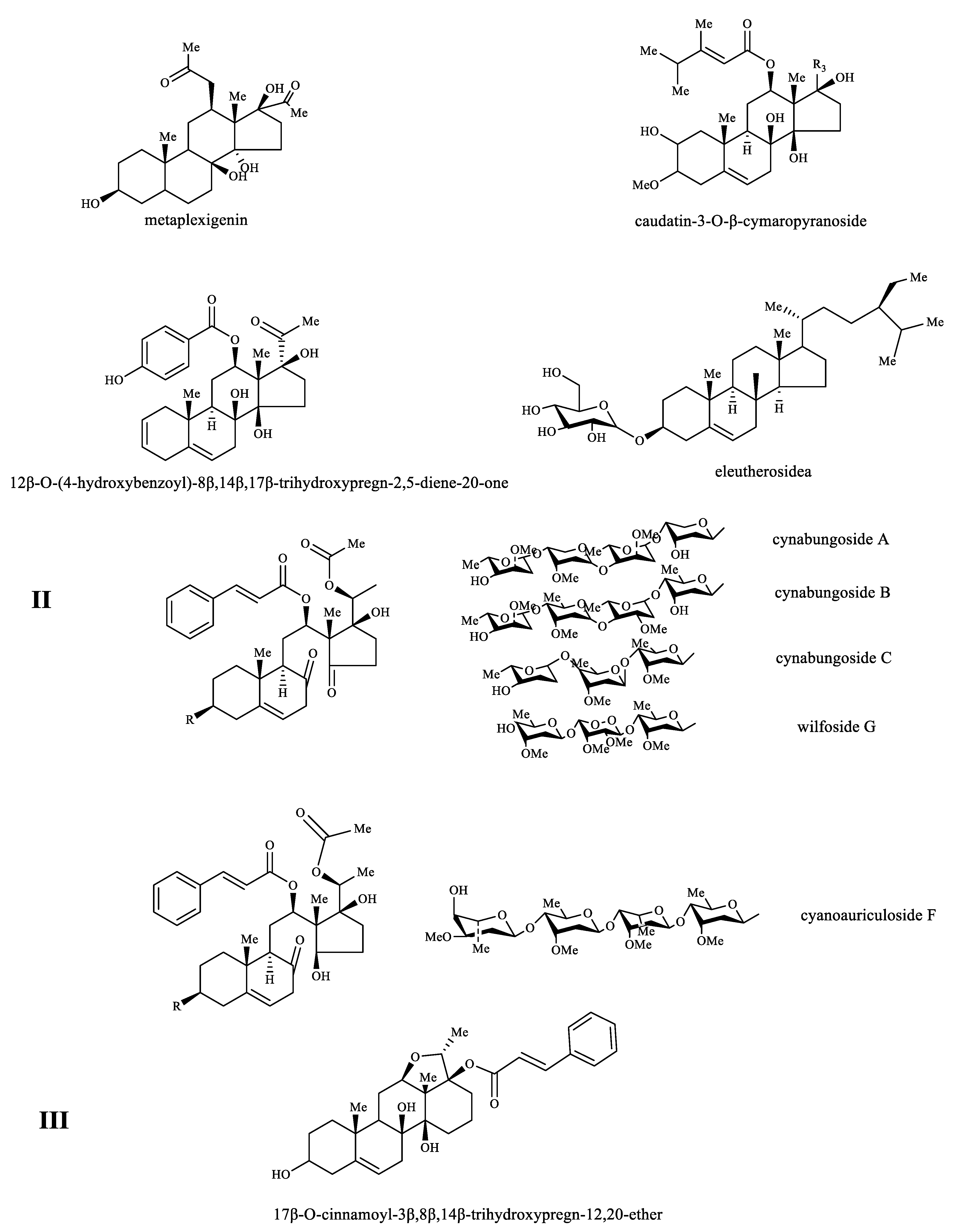
3.2.1. C21-Steroids
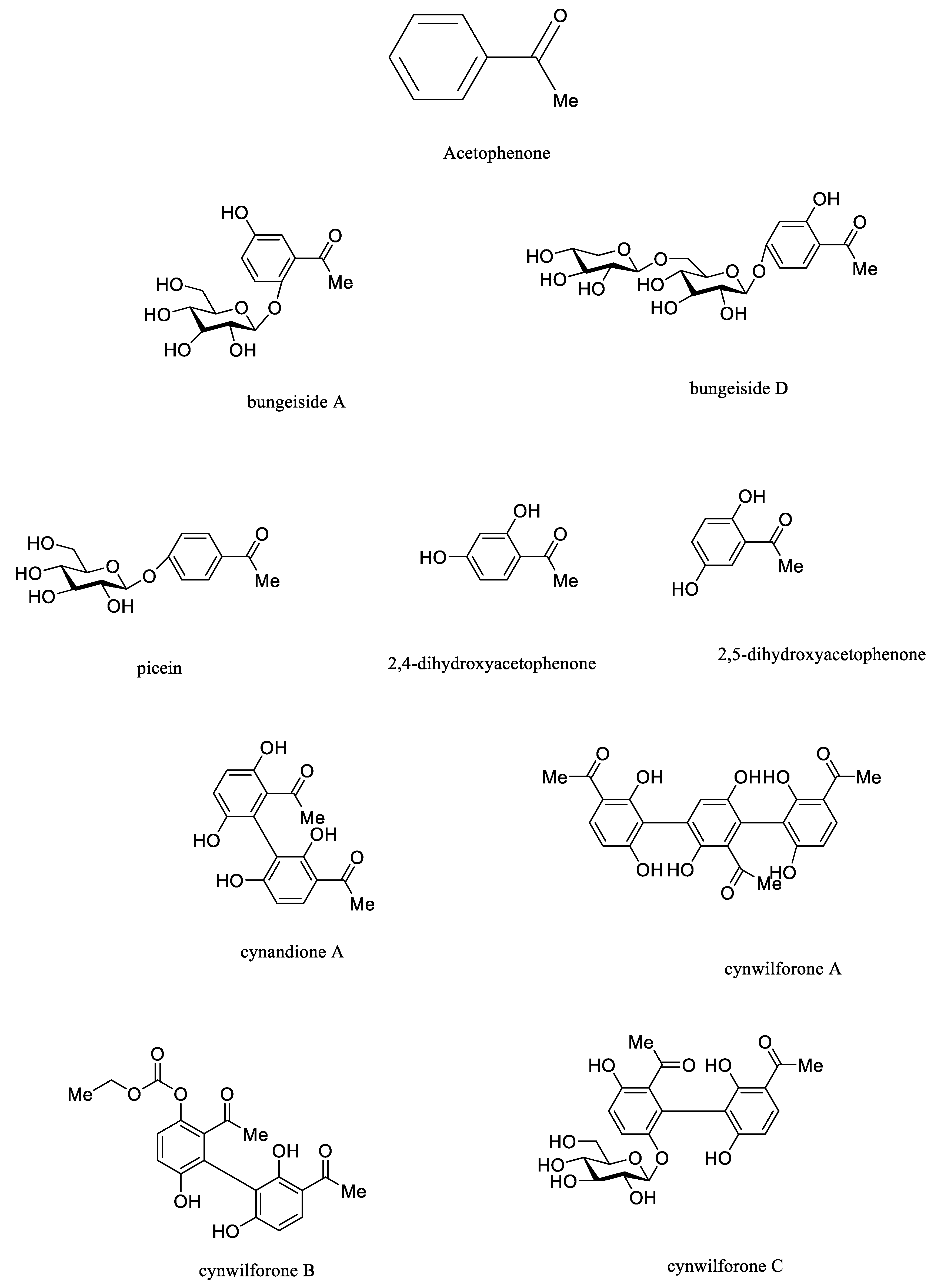
3.2.2. Acetophenones
3.2.3. Terpenoids and Alkaloids
3.2.4. Others
4. Pharmacology
4.1. Anti-Tumor Activity
4.2. Neuroprotection and Organ Protection
4.3. Immunoregulation Activity
4.4. Reducing Liver Lipid and Blood Lipid
4.5. Anti-Inflammatory Activity
4.6. Antioxidant Activity
4.7. Antidepressant Activity
4.8. Antifungal and Antiviral Activities
4.9. Others
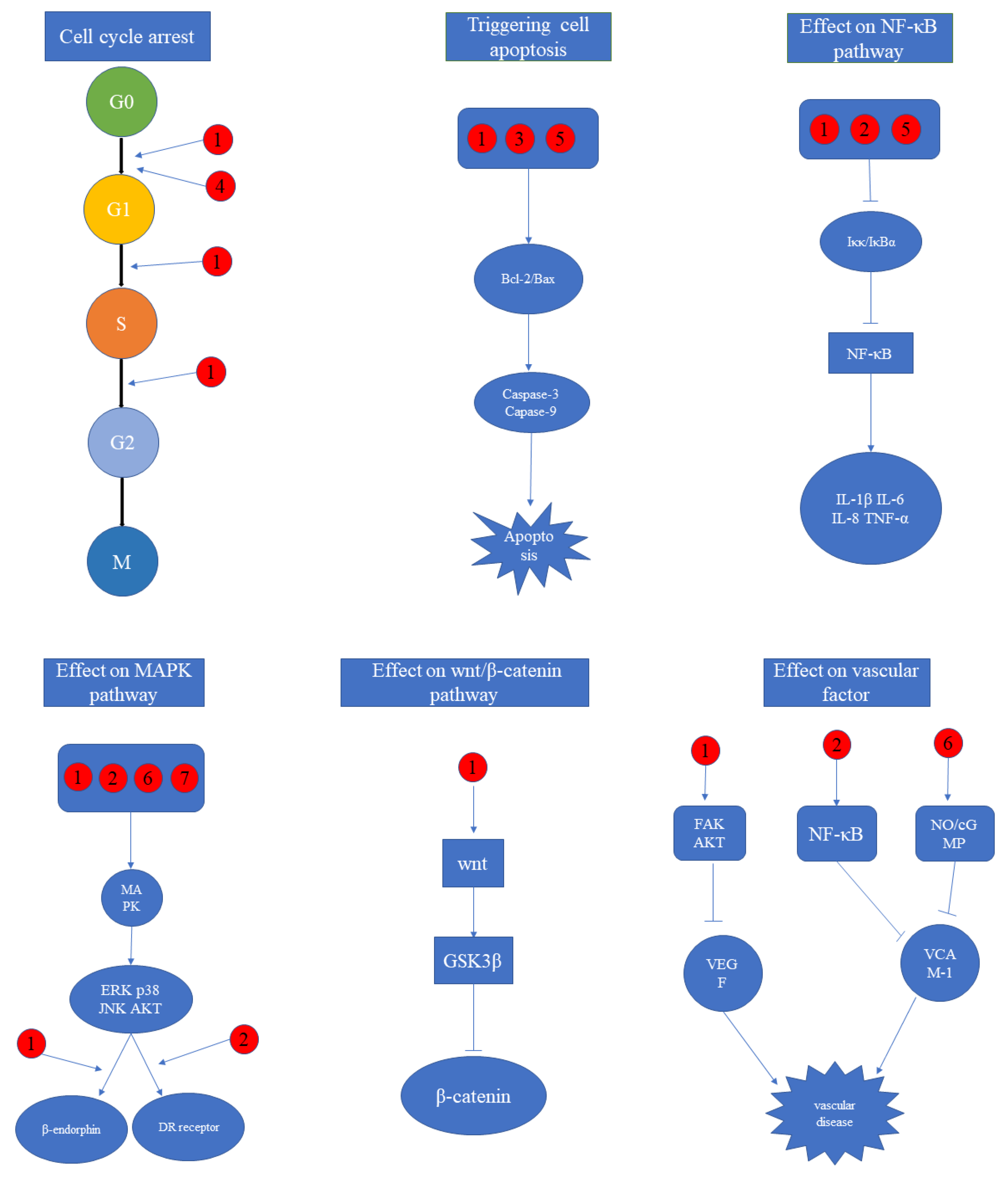
5. Molecular Mechanism
5.1. Cell Cycle Arrest
5.2. Triggering Cell Apoptosis
5.3. Effect on NF-κB Pathway
5.4. Effect on MAPK Pathway
5.5. Effect on Wnt/β-Catenin Pathway
5.6. Effect on Vascular Factor
6. Pharmacokinetics
7. Clinical Safety
8. Conclusions and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
References
- Han, L.; Zhou, X.; Yang, M.; Zhou, L.; Deng, X.; Wei, S.; Wang, W.; Wang, Z.; Qiao, X.; Bai, C. Ethnobotany, Phytochemistry and Pharmacological Effects of Plants in Genus Cynanchum Linn. (Asclepiadaceae). Molecules 2018, 23, 1194. [Google Scholar] [CrossRef]
- Yin, X.; Ding, S.F.; Shao, J.Z.; Wang, M.L.; Zhang, T.; Zhuang, Z.L.; Peng, Y.R. Research progress on Cynanchi bungei Radix. Chin. Tradit. Herbal Drugs 2019, 50, 992–1000. [Google Scholar]
- Choi, D.H.; Lee, Y.J.; Kim, J.S.; Kang, D.G.; Lee, H.S. Cynanchum wilfordii ameliorates hypertension and endothelial dysfunction in rats fed with high fat/cholesterol diets. Immunopharmacol. Immunotoxicol. 2012, 34, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Zhang, T.; Wang, Q.; Chen, J. Chromatographic fingerprint analysis of varietal differences among three species of Baishouwu and simultaneous analysis of three bioactive constituents by use of LC–DAD. Chromatographia 2008, 68, 213–218. [Google Scholar] [CrossRef]
- Jiang, Y.; Choi, H.G.; Li, Y.; Park, Y.M.; Lee, J.H.; Kim, D.H.; Lee, J.H.; Son, J.K.; Na, M.; Lee, S.H. Chemical constituents of Cynanchum wilfordii and the chemotaxonomy of two species of the family Asclepiadacease, C. wilfordii and C. auriculatum. Arch. Pharm. Res. 2011, 34, 2021–2027. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.H.; Zhang, Z.Z.; Ban, Y.F.; Rahman, K.; Ye, B.Z.; Sun, X.L.; Tan, H.Y.; Zheng, X.H.; Liu, H.Y.; Xu, L.C.; et al. Cynanchum bungei Decne and its two related species for “Baishouwu”: A review on traditional uses, phytochemistry, and pharmacological activities. J. Ethnopharmacol. 2019, 243, 112110. [Google Scholar] [CrossRef]
- Li, J.F. Present situation and Countermeasures of industrial development of Binhai baishouwu. South Chin. Agric. 2019, 13, 120–121. [Google Scholar]
- Liang, P.; Zheng, J.S.; Zhao, Z.Z. Bencao literature investigation of Polygonum multiforum (Heshouwu). Chin. J. Chin. Mater. Med. 2016, 41, 4456–4461. [Google Scholar]
- Liu, J.; Niu, J.M.; Zhao, J.H.; He, W.B.; Lv, B.X.; Li, Y.Q. Research Progress of Taishan Baishouwu. Guangdong Chem. Indutry 2019, 46, 119–120. [Google Scholar]
- Wu, C.D.; Zhang, M.; He, M.T.; Gu, M.F.; Lin, M.; Zhang, G. Selection of solvent for extraction of antioxidant components from Cynanchum auriculatum, Cynanchum bungei, and Cynanchum wilfordii roots. Food Sci. Nutr. 2019, 7, 1337–1343. [Google Scholar] [CrossRef]
- Song, H.S.; Koo, H.J.; Park, B.K.; Kwon, J.E.; Jang, S.A.; Baek, H.J.; Kim, S.Y.; Lee, S.R.; Kang, S.C. The suppressive effect of the three-herb extract mixture on vascular and liver inflammation in atherogenic diet with high fructose-fed mice. Pharm. Biol. 2018, 56, 32–42. [Google Scholar] [CrossRef]
- Sato-Masumoto, N.; Uchikura, T.; Sugiwaki, H.; Yoshimura, M.; Masada, S.; Atsumi, T.; Watanabe, M.; Tanaka, N.; Uchiyama, N.; Amakura, Y.; et al. Survey on the original plant species of crude drugs distributed as Cynanchi Wilfordii Radix and its related crude drugs in the Korean and Chinese Markets. Biol. Pharm. Bull. 2017, 40, 1693–1699. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wang, X.J.; Lv, X.H.; Li, Z.L.; Qian, S.H.; Pu, S.B. Chemical constituents from the root bark of Cynanchum auriculatum. Biochem. Syst. Ecol. 2018, 81, 30–32. [Google Scholar] [CrossRef]
- Jang, H.S.; Jeong, B.; Choi, S.Y.; Jang, G.H.; Park, K.C.; Kwon, Y.S.; Yang, H. Conduritol F, the discriminant marker between C. wilfordii and C. auriculatum by 1H NMR spectroscopy. Microchem. J. 2017, 135, 153–157. [Google Scholar] [CrossRef]
- Zhang, B.; Yao, C.F.; Wang, Y.; Jiang, L.; Ma, X.; Sun, J.; Zhang, G.Q.; Li, Y.J. Chemical constituents in water fraction of Cynanchum auriculatum. Chin. Tradit. Herbal Drugs 2018, 49, 786–790. [Google Scholar]
- Peng, X.; Wu, Y.Y.; Zhang, B.; Ma, X.; Sun, J.; Wang, G.C.; Li, Y.J. One new glycoalkaloid from the water-soluble fraction of Cynancum auriculatum. Chin. Med. Mat. 2018, 41, 2820–2822. [Google Scholar]
- Ham, W.S.; Kim, J.; Park, D.J.; Ryu, H.C.; Jang, Y.P. Discrimination of Cynanchum wilfordii and Cynanchum auriculatum by terahertz spectroscopic analysis. Phytochem. Anal. 2018, 29, 472–475. [Google Scholar] [CrossRef]
- Jun, G.; Park, T.M.; Cha, S. Fast and simple chemical fingerprinting analysis of medicinal herbs by Paper Cone Spray Ionization Mass Spectrometry (PCSI MS). Bull. Korean Chem. Soc. 2016, 37, 1337–1343. [Google Scholar] [CrossRef]
- Jo, I.H.; Kim, Y.C.; Kim, D.H.; Kim, K.H.; Hyun, T.K.; Ryu, H.; Bang, K.H. Applications of molecular markers in the discrimination of Panax species and Korean ginseng cultivars (Panax ginseng). J. Ginseng Res. 2017, 41, 444–449. [Google Scholar] [CrossRef] [PubMed]
- Han, E.H.; Cho, K.; Goo, Y.; Kim, M.; Shin, Y.W.; Kim, Y.H.; Lee, S.W. Development of molecular markers, based on chloroplast and ribosomal DNA regions, to discriminate three popular medicinal plant species, Cynanchum wilfordii, Cynanchum auriculatum, and Polygonum multiflorum. Mol. Biol. Rep. 2016, 43, 323–332. [Google Scholar] [CrossRef]
- Moon, B.C.; Choo, B.K.; Cheon, M.S.; Yoon, T.; Ji, Y.; Kim, B.B.; Lee, A.Y.; Kim, H.K. Rapid molecular authentication of three medicinal plant species, Cynanchum wilfordii, Cynanchum auriculatum, and Polygonum multiflorum (Fallopia multiflorum), by the development of RAPD-derived SCAR markers and multiplex-PCR. Plant Biotechnol. Rep. 2010, 4, 1–7. [Google Scholar] [CrossRef]
- Ryuk, J.A.; Lee, H.W.; Ju, Y.S.; Ko, B.S. Monitoring and identification of Cynanchum wilfordii and Cynanchum auriculatum by using molecular markers and real-time polymerase chain reaction. J. Korean Soc. Appl. Biol. Chem. 2014, 57, 245–251. [Google Scholar] [CrossRef]
- Guo, M.Y.; Ren, L.; Pang, X.H. Inspecting the true identity of herbal materials from Cynanchum using ITS2 barcode. Front. Plant Sci. 2017, 8, 1945. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Liu, Z.; Wang, J.; Yang, S.; Li, B.; Xu, N. Aqueous ionic liquid based ultrasonic assisted extraction of four acetophenones from the Chinese medicinal plant Cynanchum bungei Decne. Ultrason. Sonochem. 2013, 20, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Ding, W.; Liu, B.; Liu, D.; Xia, Z.; Zhang, L.; Cui, L.; Luo, Y.; Jia, X.; Feng, L. Anticancer effect of caudatin in diethylnitrosamine-induced hepatocarcinogenesis in rats. Mol. Med. Rep. 2020, 22, 697–706. [Google Scholar] [CrossRef]
- Yoon, M.Y.; Choi, N.H.; Min, B.S.; Choi, G.J.; Choi, Y.H.; Jang, K.S.; Han, S.S.; Cha, B.; Kim, J.C. Potent in vivo antifungal activity against powdery mildews of pregnane glycosides from the roots of Cynanchum wilfordii. J. Agric. Food Chem. 2011, 59, 12210–12216. [Google Scholar] [CrossRef]
- Yang, Q.X.; Ge, Y.C.; Huang, X.Y.; Sun, Q.Y. Cynanauriculoside C-E, three new antidepressant pregnane glycosides from Cynanchum auriculatum. Phytochem. Lett. 2011, 4, 170–175. [Google Scholar] [CrossRef]
- Li, Y.; Piao, D.; Zhang, H.; Woo, M.H.; Lee, J.H.; Moon, D.C.; Lee, S.H.; Chang, H.W.; Son, J.K. Quality assessment and discrimination of the roots of Cynanchum auriculatum and Cynanchum wilfordii by HPLC-UV analysis. Arch. Pharm. Res. 2013, 36, 335–344. [Google Scholar] [CrossRef]
- Wu, H.Y.; Chen, L.; Bian, Q.Y.; Wang, J. Determination of Acetophenone in Baishouwu by one assay and multiple evaluation method. Chin. Med. Mat. 2015, 38, 2339–2341. [Google Scholar]
- Yin, M.; Chen, Y.; Wang, M.; Dong, Y.; Xia, B.; Feng, X. Chemical constituents of the root of Cynanchum auriculatum. Chin. Med. Mat. 2007, 30, 1245–1247. [Google Scholar]
- Guo, N.; Li, X.P.; Xu, Z.; Han, W.J.; Lin, X.T.; Xu, X.X. Isolation and identification of C21 steroids from Cynanchum auriculatum. J. Shenyang Pharm. Univ. 2016, 33, 28–33. [Google Scholar] [CrossRef]
- Huang, L.J.; Wang, B.; Zhang, J.X.; Yan, C.; Mu, S.Z.; Hao, X.J. Studies on cytotoxic pregnane sapogenins from Cynanchum wilfordii. Fitoterapia 2015, 101, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Yao, N.; Gu, X.J.; Li, Y.B. C21 steroids study on antitumor activity of steroidal saponins of Baishouwu. Chin. Tradit. Pat. Med. 2010, 32, 1975–1978. [Google Scholar]
- Lee, G.; Choi, C.-Y.; Jun, W. Effects of aqueous extracts of Cynanchum wilfordii in rat models for postmenopausal hot flush. Prev. Nutr. Food Sci. 2016, 21, 373–377. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gu, X.J.; Yao, N.; Qian, S.H.; Li, Y.B.; Li, P. Four new C21 steroidal glycosides from the roots of Cynanchum auriculatum. Helv. Chim. Acta 2009, 92, 88–97. [Google Scholar] [CrossRef]
- Teng, H.L.; Lu, Y.; Li, J.; Yang, G.Z.; Mei, Z.N. Two new steroidal glycosides from the root of Cynanchum auriculatum. Chin. Chem. Lett. 2011, 22, 77–80. [Google Scholar] [CrossRef]
- Liu, S.; Chen, Z.; Wu, J.; Wang, L.; Wang, H.; Zhao, W. Appetite suppressing pregnane glycosides from the roots of Cynanchum auriculatum. Phytochemistry 2013, 93, 144–153. [Google Scholar] [CrossRef]
- Zhang, J.F.; Li, Y.B.; Qian, S.H.; Li, C.L.; Jiang, J.Q. Study on chemical constituents of Baishouwu. Chin. J. Chin. Pharmacol. 2006, 2006, 814–816. [Google Scholar]
- Zhang, X.; Shan, L.; Huang, H.; Yang, X.; Liang, X.; Xing, A.; Huang, H.; Liu, X.; Su, J.; Zhang, W. Rapid identification of acetophenones in two Cynanchum species using liquid chromatography-electrospray ionization tandem mass spectrometry. J. Pharm. Biomed. Anal. 2009, 49, 715–725. [Google Scholar] [CrossRef]
- Yeo, H.; Kim, J. A benzoquinone from Cynanchum wilfordii. Phytochemistry 1997, 46, 1103–1105. [Google Scholar] [CrossRef]
- He, J.B.; Jia, J.; Gou, H.P.; Zhang, X.M.; Miao, X.S.; He, F.; Tao, J. Chemical constituents of Cynanchum auriculatum. Chin. Med. Mat. 2015, 38, 2318–2320. [Google Scholar]
- Uchikura, T.; Tanaka, H.; Sugiwaki, H.; Yoshimura, M.; Sato-Masumoto, N.; Tsujimoto, T.; Uchiyama, N.; Hakamatsuka, T.; Amakura, Y. Preliminary quality evaluation and characterization of phenolic constituents in Cynanchi Wilfordii Radix. Molecules 2018, 23, 656. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Kadota, S.; Kawata, Y.; Hattori, M.; Xu, G.J.; Namba, T. Constituents of the roots of Cynanchum bungei Decne. Isolation and structures of four new glucosides, bungeiside-A, -B, -C, and -D. Chem. Pharm. Bull. 1992, 40, 3133–3137. [Google Scholar] [CrossRef] [PubMed]
- Jang, W.; Kim, K.Y.; Kim, K.; Lee, S.C.; Park, H.S.; Lee, J.; Seong, R.S.; Shim, Y.H.; Sung, S.H.; Yang, T.J. The complete chloroplast genome sequence of Cynanchum auriculatum Royle ex Wight (Apocynaceae). Mitochondrial DNA. Part A DNA Mapp. Seq. Anal. 2016, 27, 4549–4550. [Google Scholar] [CrossRef]
- Park, H.S.; Kim, K.Y.; Kim, K.; Lee, S.C.; Lee, J.; Seong, R.S.; Shim, Y.H.; Sung, S.H.; Yang, T.J. The complete chloroplast genome sequence of an important medicinal plant Cynanchum wilfordii (Maxim.) Hemsl. (Apocynaceae). Mitochondrial DNA Part A 2016, 27, 3747–3748. [Google Scholar] [CrossRef]
- Kim, J.H.; Moon, J.C.; Kang, T.S.; Kwon, K.; Jang, C.S. Development of cpDNA markers for discrimination between Cynanchum wilfordii and Cynanchum auriculatum and their application in commercial C. wilfordii food products. Appl. Biol. Chem. 2017, 60, 79–86. [Google Scholar] [CrossRef]
- Kim, C.K.; Kim, Y.K. The multipartite mitochondrial genome of Cynanchum auriculatum (Gentianales: Apocynaceae). Mitochondrial DNA B Resour. 2019, 4, 3360–3361. [Google Scholar] [CrossRef]
- Liu, S.; Yi, L.Z.; Liang, Y.Z. Traditional Chinese medicine and separation science. J. Sep. Sci. 2008, 31, 2113–2137. [Google Scholar] [CrossRef]
- Chen, Y.; Yao, F.K.; Ming, K.; Wang, D.Y.; Hu, Y.L.; Liu, J.G. Polysaccharides from traditional Chinese medicines: Extraction, purification, modification, and biological activity. Molecules 2016, 21, 1705. [Google Scholar] [CrossRef]
- Mustafa, A.; Turner, C. Pressurized liquid extraction as a green approach in food and herbal plants extraction: A review. Anal. Chim. Acta 2011, 703, 8–18. [Google Scholar] [CrossRef]
- Yang, Y.; Khan, B.M.; Zhang, X.P.; Zhao, Y.J.; Cheong, K.L.; Liu, Y. Advances in separation and purification of bioactive polysaccharides through high-speed counter-current chromatography. J. Chromatogr. Sci. 2020, 58, 992–1000. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Sun, Y.; Wang, J.; Zhu, H.; Zhou, H.; Hu, J.; Wang, J. Preparative isolation and purification of acetophenones from the Chinese medicinal plant Cynanchum bungei Decne. by high-speed counter-current chromatography. Sep. Purif. Technol. 2008, 64, 247–252. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, Z.; Wang, J.; Xiang, L.; Zhu, L. Separation and purification of baishouwubenzophenone, 4-hydroxyacetophenone and 2,4-dihydroxyacetophenone from Cynanchum auriculatum Royle ex Wight by HSCC. Chromatographia 2009, 70, 1–6. [Google Scholar] [CrossRef]
- Li, W.; Zhao, L.C.; Sun, Y.S.; Lei, F.J.; Wang, Z.; Gui, X.B.; Wang, H. Optimization of pressurized liquid extraction of three major acetophenones from Cynanchum bungei using a box-behnken design. Int. J. Mol. Sci. 2012, 13, 14533–14544. [Google Scholar] [CrossRef]
- Fei, H.R.; Liang, T.J.; Wang, L.A.; Zhou, H.L. Study on extraction and purification technology of C21 steroidal glycosides from root of Cynanchum auriculatum Royle ex Wight. Shizhen Guoyi Guoyao 2015, 26, 1091–1093. [Google Scholar] [CrossRef]
- Zhu, H.Q.; Feng, J.; Li, C.Y.; Wu, C.D.; Zhang, M.; Roth, C.M. Optimization of ultrasonic assisted extraction process and structural characterization of Polysaccharide from Baishouwu by Response Surface Methodology. Sci. Technol. Food Ind. 2021, 42, 153–159. [Google Scholar]
- Lee, M.K.; Yeo, H.; Kim, J.; Markelonis, G.J.; Oh, T.H.; Kim, Y.C. Cynandione A from Cynanchum wilfordii protects cultured cortical neurons from toxicity induced by H2O2, L-glutamate, and kainate. J. Neurosci. Res. 2000, 59, 259–264. [Google Scholar] [CrossRef]
- Kim, S.; Yoon, Y.Y.; Park, Y.W.; Whang, W.K.; Park, S.Y.; Hwang, K.W. Cynandione A from Cynanchum wilfordii inhibits hepatic de novo lipogenesis by activating the LKB1/AMPK pathway in HepG2 cells. J. Nat. Med. 2020, 74, 142–152. [Google Scholar] [CrossRef]
- Wang, X.J.; Li, Z.L.; Lv, X.H.; Zuo, Q.Y.; Zhao, Y.M.; Ding, Y.F.; Pu, S.B.; Qian, S.H.; Peng, Y.R. Antitumor evaluation and multiple analysis on different extracted fractions of the root of Cynanchum auriculatum Royle ex Wight. J. Sep. Sci. 2017, 40, 3054–3063. [Google Scholar] [CrossRef] [PubMed]
- Ke, S.Y. Recent progress of novel steroid derivatives and their potential biological properties. Mini Rev. Med. Chem. 2018, 18, 745–775. [Google Scholar] [CrossRef]
- Qi, L.W.; Gu, X.J.; Li, P.; Liang, Y.; Hao, H.; Wang, G. Structural characterization of pregnane glycosides from Cynanchum auriculatum by liquid chromatography on a hybrid ion trap time-of-flight mass spectrometer. Rapid Commun. Mass Spectrom. 2009, 23, 2151–2160. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.W.; Xie, S.; Hu, S.Y.; Liao, T.; Liu, P.; Peng, K.H.; Yang, X.Z.; He, Z.L.; Tang, H.Y.; Cui, Y.; et al. Caudatin targets TNFAIP1/NF-kappa B and cytochrome c/caspase signaling to suppress tumor progression in human uterine cancer. Int. J. Oncol. 2016, 49, 1638–1650. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.W.; Kim, C.; Le, D.U. Effect of culture conditions on the biosynthesis of gagaminine, a potent antioxidant from the roots of Cynanchum wilfordii. Biol. Pharm. Bull. 2001, 24, 1451–1453. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Li, Y.; Zhang, J.; Gu, X.; Peng, Y.; Huang, W.; Qian, S. Two new cytotoxic pregnane glycosides from Cynanchum auriculatum. Planta Med. 2008, 74, 551–554. [Google Scholar] [CrossRef]
- Tarkowska, D. Plants are capable of synthesizing animal steroid hormones. Molecules 2019, 24, 2585. [Google Scholar] [CrossRef]
- Tarkowska, D.; Strnad, M. Isoprenoid-derived plant signaling molecules: Biosynthesis and biological importance. Planta 2018, 247, 1051–1066. [Google Scholar] [CrossRef]
- Lindemann, P. Steroidogenesis in plants—Biosynthesis and conversions of progesterone and other pregnane derivatives. Steroids 2015, 103, 145–152. [Google Scholar] [CrossRef]
- Ziarani, G.M.; Kheilkordi, Z.; Mohajer, F. Recent advances in the application of acetophenone in heterocyclic compounds synthesis. J. Iranian Chem. Soc. 2020, 17, 247–282. [Google Scholar] [CrossRef]
- Lee, M.K.; Yeo, H.; Kim, J.; Kim, Y.C. Protection of rat hepatocytes exposed to CCl4 in vitro by cynandione A, a biacetophenone from Cynanchum wilfordii. J. Pharm. Pharmacol. 2000, 52, 341–345. [Google Scholar] [CrossRef]
- Sawamoto, A.; Kanazaki, A.; Amakura, Y.; Yoshimura, M.; Masumoto, N.; Uchiyama, N.; Hakamatsuka, T.; Okuyama, S.; Furukawa, Y.; Nakajima, M. Cynandione A induces adipogenesis and beige adipocyte-related phenotype in 3T3-L1 cells. Phytochem. Lett. 2020, 39, 84–89. [Google Scholar] [CrossRef]
- Kim, J.Y.; Lee, J.W.; Lee, J.S.; Jang, D.S.; Shim, S.H. Inhibitory effects of compounds isolated from roots of Cynanchum wilfordii on oxidation and glycation of human low-density lipoprotein (LDL). J. Funct. Foods 2019, 59, 281–290. [Google Scholar] [CrossRef]
- Yang, W.Q.; Chen, X.; Li, Y.L.; Guo, S.F.; Wang, Z.; Yu, X.L. Advances in pharmacological activities of terpenoids. Nat. Prod. Commun. 2020, 15, 1934578X20903555. [Google Scholar] [CrossRef]
- Zhou, J.L.; Qiu, S.Y.; Chen, X.Y. Properties and research progress of terpenoids in Jatropha curcas. Guizhou Chem. Ind. 2008, 2, 11–18. [Google Scholar]
- Li, J.L.; Gao, Z.B.; Zhao, W.M. Identification and evaluation of antiepileptic activity of C21 steroidal glycosides from the roots of Cynanchum wilfordii. J. Nat. Prod. 2016, 79, 89–97. [Google Scholar] [CrossRef]
- Debnath, B.; Singh, W.S.; Das, M.; Goswami, S.; Singh, M.K.; Maiti, D.; Manna, K. Role of plant alkaloids on human health: A review of biological activities. Mater. Today Chem. 2018, 9, 56–72. [Google Scholar] [CrossRef]
- Zhao, Y.; Yan, B.C.; Wang, Z.Y.; Li, M.J.; Zhao, W. Natural polysaccharides with immunomodulatory activities. Mini Rev. Med. Chem. 2020, 20, 96–106. [Google Scholar] [CrossRef]
- Xu, Y.; Ren, R. Isolation and physic-chemical properties of polysaccharides from the root of Cynanchum auriculatum Royle ex Wight. China J. Chin. Mat. Med. 1991, 16, 163–166. [Google Scholar]
- Jang, M.; Lim, T.G.; Lee, E.; Hong, H.D.; Rhee, Y.K.; Lee, Y.J.; Lee, J.H.; Cho, C.W. Immunostimulatory activities of a high molecular weight fraction of Cynanchum auriculatum Royle ex Wight root obtained by ultrafiltration. Pharmacogn. Mag. 2020, 16, 493–498. [Google Scholar] [CrossRef]
- Cho, C.W.; Ahn, S.; Lim, T.G.; Hong, H.D.; Rhee, Y.K.; Yang, D.C.; Jang, M. Cynanchum wilfordii polysaccharides suppress dextran sulfate sodium-induced acute colitis in mice and the production of inflammatory mediators from macrophages. Mediat. Inflamm. 2017, 2017, 3859856. [Google Scholar] [CrossRef] [PubMed]
- Chai, Z.; Huang, W.; Zhao, X.; Wu, H.; Zeng, X.; Li, C. Preparation, characterization, antioxidant activity and protective effect against cellular oxidative stress of polysaccharide from Cynanchum auriculatum Royle ex Wight. Int. J. Biol. Macromol. 2018, 119, 1068–1076. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Ye, Y.; Shen, Y.; Liang, H. Studies on cytotoxic constituents of Cynanchum auriculatum Royle ex Wight. Acta Pharm. Sincia 2000, 35, 431–437. [Google Scholar]
- Lu, Y.; Teng, H.L.; Yang, G.Z.; Mei, Z.N. Three new steroidal glycosides from the roots of Cynanchum auriculatum. Molecules 2011, 16, 1901–1909. [Google Scholar] [CrossRef] [PubMed]
- Song, J.M.; Tao, G.J.; Tang, J.; Ding, X.L. Isolation and characterization of C21 steroid glycosides from Cynanchum auriculatum. J. Wuxi Univ. Light Ind. 2002, 21, 176–178. [Google Scholar]
- Li, Z.G.; Wu, J.; Chen, S.Q.; Zhu, L.; Jiang, R.; Cai, Y.S. One new C21 steroid compound from Cynanchum auriculatum. Chin. Tradit. Herbal Drugs 2020, 51, 5921–5923. [Google Scholar]
- Gan, H.; Xiang, W.J.; Ma, L.; Hu, L.H. Six new C21 steroid glycosides from Cynanchum bungei Dence. Helv. Chim. Acta 2008, 91, 2222–2234. [Google Scholar] [CrossRef]
- Qin, J.J.; Chen, X.; Lin, Z.M.; Xu, Y.S.; Li, Y.M.; Zuo, J.P.; Zhao, W.M. C21-steroidal glycosides and sesquiterpenes from the roots of Cynanchum bungei and their inhibitory activities against the proliferation of B and T lymphocytes. Fitoterapia 2018, 124, 193–199. [Google Scholar] [CrossRef]
- Hao, S.J.; Gao, L.J.; Xu, S.F.; Ye, Y.P.; Li, X.Y. Six new steroidal glycosides from roots of Cynanchum bungei. Phytochem. Lett. 2018, 23, 26–32. [Google Scholar] [CrossRef]
- Xiang, W.J.; Ma, L.; Hu, L.H. C21 Steroidal glycosides from Cynanchum wilfordii. Helv. Chim. Acta 2009, 92, 2659–2674. [Google Scholar] [CrossRef]
- Zhao, J.W. Taishan Baishouwu: Isolation and Identification of C21 Steroids and Their Inhibition of Hedgehog Signal Pathway Activity. Master’s Thesis, Zhejiang Academy of Medical Sciences, Hangzhou, China, 2017. [Google Scholar]
- Zhang, M.; Wang, D.; Li, B. Neuroprotection of two C21 steroidal glycosides from Cynanchum auriculatum against H2O2-induced damage on PC12 cells. Nat. Prod. Res. 2019, 35, 1752–1755. [Google Scholar] [CrossRef]
- Li, X.Y.; Zhou, L.F.; Gao, L.J.; Wei, Y.; Xu, S.F.; Chen, F.Y.; Huang, W.J.; Tan, W.F.; Ye, Y.P. Cynanbungeigenin C and D, a pair of novel epimers from Cynanchum bungei, suppress hedgehog pathway-dependent medulloblastoma by blocking signaling at the level of Gli. Cancer Lett. 2018, 420, 195–207. [Google Scholar] [CrossRef]
- Peng, Y.R.; Li, Y.B.; Liu, X.D.; Zhang, J.F.; Duan, J.A. Antitumor activity of C21 steroidal glycosides from Cynanchum auriculatum Royle ex Wight. Phytomedicine 2008, 15, 1016–1020. [Google Scholar] [CrossRef]
- Zhang, R.; Liu, Y.; Wang, Y.; Ye, Y.; Li, X. Cytotoxic and apoptosis-inducing properties of auriculoside A in tumor cells. Chem. Biodivers. 2007, 4, 887–892. [Google Scholar] [CrossRef]
- Chen, J.J.; Zhang, Z.X.; Zhou, J. Structures of cynauricusidc A, B and C. Acta Bot. Yunnanica 1990, 2, 197–210. [Google Scholar]
- Hwang, B.Y.; Kim, S.E.; Kim, Y.H.; Kim, H.S.; Hong, Y.S.; Ro, J.S.; Lee, K.S.; Lee, J.J. Pregnane glycoside multidrug-resistance modulators from Cynanchum wilfordii. J. Nat. Prod. 1999, 62, 640–643. [Google Scholar] [CrossRef]
- Hayashi, K.; Mitsuhashi, H. Studies on the constituents of Asclepiadaceae plants: Aglycones from Cynanchum wilfordi HEMSLEY. Chem. Pharm. Bull. 1975, 23, 139–143. [Google Scholar] [CrossRef][Green Version]
- Qian, X.; Li, B.; Li, P.; Wang, D.; Dai, W.; Zhang, M. C21 steroidal glycosides from Cynanchum auriculatum and their neuroprotective effects against H2O2-induced damage in PC12 cells. Phytochemistry 2017, 140, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.J.; Zhang, Z.X.; Zhou, J. Chemical constituents of Cynanchum auriculatum Royle ex Wight. Acta Bot. Yunnanica 1989, 3, 358–360. [Google Scholar]
- Jiang, H.W.; Gu, S.S.; Cao, L.; Jiang, S.S.; Lin, J.; Chen, Y.; Wang, L.; Jiao, H.; Li, J.Y.; Zhao, W.M.; et al. Potential hypoglycemic effect of acetophenones from the root bark of Cynanchum wilfordii. Nat. Prod. Res. 2019, 33, 2314–2321. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.W.; Lin, J.; Wang, G.M.; Zhang, J.J.; Gu, S.S.; Cao, L.; Chen, Y.; Wang, L.; Jiao, H.; Zhu, W.L.; et al. Acetophenone derivatives from the root bark of Cynanchum wilfordii as potential neuroprotective agents. Phytochem. Lett. 2018, 24, 179–183. [Google Scholar] [CrossRef]
- Sun, Y.; Lin, H.; Wang, J.; Hu, J.; Liu, Z.; Gao, A. An application of high-speed counter-current chromatography for separation and purification of bungeiside-A, bungeiside-B and baishouwubenzophenone from Cynanchum bungei Decne. Phytochem. Anal. 2011, 22, 526–531. [Google Scholar] [CrossRef]
- Xie, K.Q. Study on Chemical Constituents and Bioactivity of Cynanchum wilfordii (Maxim.) Hemsl. Master’s Thesis, Guizhou University, Guiyang, China, 2017. [Google Scholar]
- Hwang, B.Y.; Kim, Y.H.; Ro, J.S.; Lee, K.S.; Lee, J.J. Acetophenones from the roots of Cynanchum wilfordii (Maxim.) Hemsl. Arch. Pharm. Res. 1999, 22, 72–74. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xu, B.X.; Liang, G.Y.; Cao, P.X. Studies on the Chemical constituents of Cynanchum auriculatum Royle ex Wight. Nat. Prod. Res. Dev. 2008, 20, 1012–1021. [Google Scholar]
- Chen, B.Y.; Qiu, R.C.; Liu, F.; Li, X.; Li, Y.Y.; Wu, G.Z.; Shan, L. Study on acetophenone compounds and their antioxidant activity in Cynanchum auriculatum Royle ex Wight. J. Pharm. Prac. 2013, 31, 351–354. [Google Scholar]
- Chen, Y. Study on Chemical Constituents of Cynanchum wilfordii (Maxim.) Hemsl., a National Medicine; Guizhou University: Guiyang, China, 2008. [Google Scholar]
- Pang, Y.J. Studies on Chemical Constituents and Activities of Taishan baishouwu; Shandong University of Traditional Chinese Medicine: Jinan, China, 2011. [Google Scholar]
- Wu, Z.H.; Meng, X.; Ding, Y.F.; Peng, Y.R.; Wu, Z.H.; Meng, X.; Ding, Y.F.; Peng, Y.R. Effects of total C21 steroid glycosides from Baishouwu on proliferation invasion and apoptosis of human hepatic cancer HepG2 cells. China J. Chin. Mat. Med. 2018, 43, 1915–1921. [Google Scholar]
- Ding, Y.F.; Peng, Z.X.; Ding, L.; Peng, Y.R. Baishouwu extract suppresses the development of hepatocellular carcinoma via TLR4/MyD88/ NF-κB pathway. Front. Pharmacol. 2019, 10, 389. [Google Scholar] [CrossRef]
- Zhen, X.; Choi, H.S.; Kim, J.H.; Kim, S.L.; Liu, R.; Ko, Y.C.; Yun, B.S.; Lee, D.S. Caudatin isolated from Cynanchum auriculatum inhibits breast cancer stem cell formation via a GR/YAP signaling. Biomolecules 2020, 10, 925. [Google Scholar] [CrossRef]
- Zhu, L.Z.; Hou, Y.J.; Zhao, M.; Yang, M.F.; Fu, X.T.; Sun, J.Y.; Fu, X.Y.; Shao, L.R.; Zhang, H.F.; Fan, C.D.; et al. Caudatin induces caspase-dependent apoptosis in human glioma cells with involvement of mitochondrial dysfunction and reactive oxygen species generation. Cell Biol. Toxicol. 2016, 32, 333–345. [Google Scholar] [CrossRef]
- Li, X.F.; Zhang, X.T.; Liu, X.Z.; Tan, Z.W.; Yang, C.L.; Ding, X.F.; Hu, X.; Zhou, J.L.; Xiang, S.L.; Zhou, C.; et al. Caudatin induces cell apoptosis in gastric cancer cells through modulation of Wnt/β-catenin signaling. Oncol. Rep. 2013, 30, 677–684. [Google Scholar] [CrossRef]
- Wang, Y.Q.; Zhang, S.J.; Lu, H.; Yang, B.; Ye, L.F.; Zhang, R.S. A C21-steroidal glycoside isolated from the roots of Cynanchum auriculatum induces cell cycle arrest and apoptosis in human gastric cancer SGC-7901 cells. Evid. Based. Complement. Alternat. Med. eCAM 2013, 2013, 180839. [Google Scholar] [CrossRef][Green Version]
- Liu, K.; Chen, F.; Zhang, H. Antitumor effects by Wilfoside C3N treatment in ECA109 cells. Anticancer Drugs 2010, 21, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Fei, H.R.; Cui, L.Y.; Zhang, Z.R.; Zhao, Y.; Wang, F.Z. Caudatin inhibits carcinomic human alveolar basal epithelial cell growth and angiogenesis through modulating GSK3β/β-catenin pathway. J. Cell. Biochem. 2012, 113, 3403–3410. [Google Scholar] [CrossRef]
- Yue, R.; Yuan, X.; Liu, X.; Zhang, J.; Jiang, P.; He, C.; Shan, L.; Yu, Y.; Zhang, W. Cynandione A mitigates ischemic injuries in rats with cerebral ischemia. J. Neurochem. 2012, 121, 451–464. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.W.; Peng, Y.R.; Ding, Y.F. Effects of total C21 steroid glucosides from Cynanchum auriculatum on oxidative stress pathway in mice with liver injury. China J. Chin. Mat. Med. 2019, 44, 2960–2965. [Google Scholar]
- Zhang, W.; Dong, Z.; Zhao, B.; Zeng, Z. Effect of polysaccharide from Cynanchum auriculatum Royle ex Wight against CCl4-induced liver injury in mice. Mod. Food Sci. Technol. 2011, 36, 57–63. [Google Scholar]
- Shan, L.; Liu, R.H.; Shen, Y.H.; Zhang, W.D.; Zhang, C.; Wu, D.Z.; Min, L.; Su, J.; Xu, X.K. Gastroprotective effect of a traditional Chinese herbal drug “Baishouwu” on experimental gastric lesions in rats. J. Ethnopharmacol. 2006, 107, 389–394. [Google Scholar] [CrossRef]
- Liu, T.; Yang, S.; Li, N.; Pan, J.; Lu, Y.; Li, Y.; Zhong, M.; Sun, J. The inhibitory effects of water-soluble fraction of Cynanchum wilfordii on atropine-induced gastrointestinal disfunction in mice. J. Guizhou Med. Univ. 2018, 43, 1252–1255. [Google Scholar] [CrossRef]
- Ding, H.Y.; Chang, T.S.; Shen, H.C.; Tai, S.S. Murine tyrosinase inhibitors from Cynanchum bungei and evaluation of in vitro and in vivo depigmenting activity. Exp. Dermatol. 2011, 20, 720–724. [Google Scholar] [CrossRef]
- Jang, S.A.; Lee, S.; Sohn, E.H.; Yang, J.; Park, D.W.; Jeong, Y.J.; Kim, I.; Kwon, J.E.; Song, H.S.; Cho, Y.M.; et al. Cynanchum wilfordii Radix attenuates liver fat accumulation and damage by suppressing hepatic cyclooxygenase-2 and mitogen-activated protein kinase in mice fed with a high-fat and high-fructose diet. Nutr. Res. 2016, 36, 914–924. [Google Scholar] [CrossRef]
- Lee, H.S.; Choi, J.H.; Kim, Y.E.; Kim, I.H.; Kim, B.M.; Lee, C.H. Effects of the Cynanchum wilfordii ethanol extract on the serum lipid profile in hypercholesterolemic rats. Prev. Nutr. Food Sci. 2013, 18, 157–162. [Google Scholar] [CrossRef]
- Kim, S.H.; Lee, T.H.; Lee, S.M.; Park, J.H.; Park, K.H.; Jung, M.; Jung, H.; Mohamed, M.A.A.; Baek, N.I.; Chung, I.S.; et al. Cynandione A attenuates lipopolysaccharide-induced production of inflammatory mediators via MAPK inhibition and NF-κB inactivation in RAW264.7 macrophages and protects mice against endotoxin shock. Exp. Biol. Med. 2015, 240, 946–954. [Google Scholar]
- Koo, H.J.; Sohn, E.H.; Pyo, S.; Woo, H.G.; Park, D.W.; Ham, Y.M.; Jang, S.A.; Park, S.Y.; Kang, S.C. An ethanol root extract of Cynanchum wilfordii containing acetophenones suppresses the expression of VCAM-1 and ICAM-1 in TNF-α-stimulated human aortic smooth muscle cells through the NF-κB pathway. Int. J. Mol. Med. 2015, 35, 915–924. [Google Scholar] [CrossRef]
- Wu, Z.; Wang, Y.; Meng, X.; Wang, X.; Li, Z.; Qian, S.; Wei, Y.; Shu, L.; Ding, Y.; Wang, P.; et al. Total C21 steroid glycosides, isolated from the root tuber of Cynanchum auriculatum Royle ex Wight, attenuate hydrogen peroxide-induced oxidative injury and inflammation in L02 cells. Int. J. Mol. Med. 2018, 42, 3157–3170. [Google Scholar] [CrossRef] [PubMed]
- Ji, C.X.; Li, X.Y.; Jia, S.B.; Liu, L.L.; Ge, Y.C.; Yang, Q.X.; Zhang, J.J. The antidepressant effect of Cynanchum auriculatum in mice. Pharm. Bio. 2012, 50, 1067–1072. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Huang, L.J.; Wang, B.; Zhang, J.X.; Yuan, C.M.; Gu, W.; Mu, S.Z.; Hao, X.J. Synthesis and evaluation of antifungal activity of C21-steroid derivatives. Bioorg. Med. Chem. Lett. 2016, 26, 2040–2043. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.Y.; Liu, A.L.; Liu, S.J.; Xu, X.W.; Huang, L.F. Screening for neuraminidase inhibitory activity in traditional Chinese medicines used to treat influenza. Molecules 2016, 21, 1138. [Google Scholar] [CrossRef]
- Kim, M.S.; Baek, J.H.; Park, J.A.; Hwang, B.Y.; Kim, S.E.; Lee, J.J.; Kim, K.W. Wilfoside K1N isolated from Cynanchum wilfordii inhibits angiogenesis and tumor cell invasion. Int. J. Oncol. 2005, 26, 1533–1539. [Google Scholar] [CrossRef]
- Lee, G.; Kim, J.; Pan, S.O.; Kim, M.; Jun, W.; Choi, C.Y. Aphrodisiac property of the aqueous extract of Cynanchum wilfordii. J. Food Nutr. Res. 2016, 4, 713–719. [Google Scholar] [CrossRef]
- Lee, E.; Hong, H.D.; Jang, M.; Lim, T.G.; Kim, T.; Ha, H.; Lee Jeong, H.; Cho, C.W. Selective activation of the estrogen receptor-β by the polysaccharide from Cynanchum wilfordii alleviates menopausal syndrome in ovariectomized mice. Int. J. Biol. Macromol. 2020, 165, 1029–1037. [Google Scholar] [CrossRef]
- Lee, G.; Shin, J.; Choi, H.; Jo, A.; Pan, S.; Bae, D.; Lee, Y.; Choi, C. Cynanchum wilfordii ameliorates testosterone-induced benign prostatic hyperplasia by regulating 5α-reductase and androgen receptor activities in a rat model. Nutrients 2017, 9, 1070. [Google Scholar] [CrossRef]
- Jun, W.; Choi, C.Y.; Lee, G.; Pan, S.; Kim, J. Bone regenerative effect of aqueous Cynanchum wilfordii extract in receptor activator of nuclear factor-κB ligand-induced osteoclast differentiation and estrogen deficiency-induced osteoporosis. Int. J. Phytomed. 2017, 9, 461–470. [Google Scholar] [CrossRef]
- Lee, H.; Kim, M.H.; Choi, Y.Y.; Hong, J.; Yang, W.M. Effects of Cynanchum wilfordii on osteoporosis with inhibition of bone resorption and induction of bone formation. Mol. Med. Rep. 2018, 17, 3758–3762. [Google Scholar] [CrossRef]
- Luo, Y.; Sun, Z.G.; Li, Y.B.; Liu, L.N.; Cai, X.T.; Li, Z.L. Caudatin inhibits human hepatoma cell growth and metastasis through modulation of the Wnt/β-catenin pathway. Oncol. Rep. 2013, 30, 2923–2928. [Google Scholar] [CrossRef] [PubMed]
- Park, K.H.; Kim, J.; Lee, E.H.; Lee, T.H. Cynandione A inhibits lipopolysaccharide-induced cell adhesion via suppression of the protein expression of VCAM-1 in human endothelial cells. Int. J. Mol. Med. 2018, 41, 1756–1764. [Google Scholar] [CrossRef] [PubMed]
- Saitou, T.; Imamura, T. Quantitative imaging with Fucci and mathematics to uncover temporal dynamics of cell cycle progression. Dev. Growth Differ. 2016, 58, 6–15. [Google Scholar] [CrossRef]
- Fei, H.R.; Chen, H.L.; Xiao, T.; Chen, G.; Wang, F.Z. Caudatin induces cell cycle arrest and caspase-dependent apoptosis in HepG2 cell. Mol. Biol. Rep. 2012, 39, 131–138. [Google Scholar] [CrossRef]
- Fu, X.Y.; Zhang, S.; Yang, M.F.; Fan, C.D.; Sun, B.L.; Wang, K.; Sun, B.L. Caudatin inhibits human glioma cells growth through triggering DNA damage-mediated cell cycle arrest. Cell. Mol. Neurobiol. 2015, 35, 953–959. [Google Scholar] [CrossRef]
- Lowe, S.W.; Lin, A.W. Apoptosis in cancer. Carcinogenesis 2000, 21, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Pai, P.; Sukumar, S. HOX genes and the NF-kappa B pathway: A convergence of developmental biology, inflammation and cancer biology. Biochim. Biophys. Acta Rev. Cancer 2020, 1874, 188450. [Google Scholar] [CrossRef]
- Napetschnig, J.; Wu, H. Molecular basis of NF-kappa B signaling. Annu. Rev. Biophys. 2013, 42, 443–468. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.B.; Lee, S.M.; Park, J.H.; Lee, T.H.; Baek, N.I.; Park, H.J.; Lee, H.; Kim, J. Cynandione A from Cynanchum wilfordii attenuates the production of inflammatory mediators in LPS-induced BV-2 microglial cells via NF-κB inactivation. Biol. Pharm. Bull. 2014, 37, 1390–1396. [Google Scholar] [CrossRef] [PubMed]
- Shiryaev, A.; Moens, U. Mitogen-activated protein kinase p38 and MK2, MK3 and MK5: Menage a trois or menage a quatre? Cell. Signal. 2010, 22, 1185–1192. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Mao, X.F.; Wu, H.Y.; Liu, H.; Sun, M.L.; Wang, X.; Wang, Y.X. Cynandione A attenuates neuropathic pain through p38β MAPK-mediated spinal microglial expression of β-endorphin. Brain Behav. Immun. Health 2017, 62, 64–77. [Google Scholar] [CrossRef]
- Fei, H.R.; Yuan, C.; Wang, G.L.; Zhao, Y.; Li, Z.J.; Du, X.; Wang, F.Z. Caudatin potentiates the anti-tumor effects of TRAIL against human breast cancer by upregulating DR5. Phytomedicine 2019, 62, 152950. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.; Cho, C.W.; Han, A.R.; Rhee, Y.K.; Hong, H.D.; Kim, K.T.; Lee, S.Y.; Lee, K.M.; Jung, S.K.; Lim, T.G. MMP-1 suppressing activity of high-molecular weight fraction isolated from the roots of Cynanchum wilfordii. J. Funct. Foods 2018, 40, 329–335. [Google Scholar] [CrossRef]
- Harb, J.; Lin, P.J.; Hao, J.J. Recent development of wnt signaling pathway inhibitors for cancer therapeutics. Curr. Oncol. Rep. 2019, 21, 12. [Google Scholar] [CrossRef] [PubMed]
- Fallah, A.; Sadeghinia, A.; Kahroba, H.; Samadi, A.; Heidari, H.R.; Bradaran, B.; Zeinali, S.; Molavi, O. Therapeutic targeting of angiogenesis molecular pathways in angiogenesis-dependent diseases. Biomed. Pharmacother. 2019, 110, 775–785. [Google Scholar] [CrossRef]
- Wang, X.Z.; Fu, X.T.; Zhao, S.J.; Fu, X.Y.; Zhang, H.F.; Shao, L.R.; Li, G.; Fan, C.D. Antiangiogenic properties of caudatin in vitro and in vivo by suppression of VEGF-VEGFR2-AKT/FAK signal axis. Mol. Med. Rep. 2017, 16, 8937–8943. [Google Scholar] [CrossRef]
- Choi, D.H.; Lee, Y.J.; Oh, H.C.; Cui, Y.L.; Kim, J.S.; Kang, D.G.; Lee, H.S. Improved endothelial dysfunction by Cynanchum wilfordii in apolipoprotein E-/- mice fed a high fat/cholesterol diet. J. Med. Food 2012, 15, 169–179. [Google Scholar] [CrossRef]
- Yellepeddi, V.; Rower, J.; Liu, X.X.; Kumar, S.; Rashid, J.; Sherwin, C.M.T. State-of-the-Art review on physiologically based pharmacokinetic modeling in pediatric drug development. Clin. Pharmacokinet. 2019, 58, 1–13. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Shan, L.; Xu, W.; Yang, M.; Zhang, W.D.; Li, H.Y. LC-MS/MS method for the determination of cynandione A in rat plasma and tissues. Biomed. Chromatogr. 2008, 22, 1173–1179. [Google Scholar] [CrossRef]
- Zhu, Q.Q.; Hu, Y.Y.; Shan, Y.Y.; Wang, Y.Y.; Wu, X.L.; Mao, B.P.; Ge, R.S. Determination of caudatin in rat plasma by UPLC-MS/MS: Application to a preclinical pharmacokinetic study. Pharmacology 2015, 96, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.R.; Li, Y.B.; Wang, D.W.; Liu, X.D.; Zhang, J.F.; Qian, S.H.; Duan, J.N. Determination of caudatin-2,6-dideoxy-3-O-methy-β-d-cymaropyranoside in rat plasma using liquid chromatography-tandem mass spectrometry. Biomed. Chromatogr. 2008, 22, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Shan, L.; Liu, X.; Li, H.; Hu, L.; Zhang, W. Determination of two C21 steroidal glycosides of Baishouwu and their metabolites in mice plasma and tumor homogenate by liquid chromatography-tandem mass spectrometry. J. Chromatogr. B. Analyt. Technol. Biomed. Life Sci. 2008, 876, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Rao, D.R.; Sordillo, J.E.; Kopel, L.S.; Gaffin, J.M.; Sheehan, W.J.; Hoffman, E.; Ozonoff, A.; Gold, D.R.; Phipatanakul, W. Eosinophilic esophagitis associated with Cynanchum wilfordii. Ann. Allergy. Asthma. Immunol. 2015, 114, 257–265. [Google Scholar] [CrossRef]
- Youn, J.S.; Ham, Y.M.; Yoon, W.J.; Choi, H.C.; Lee, J.E.; Cho, B.; Kim, J.Y. Cynanchum wilfordii etanolic extract controls blood cholesterol: A double-blind, randomized, placebo-controlled, parallel trial. Nutrients 2019, 11, 836. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.M.; Cho, Y.M.; Kwon, J.E.; Lee, S.R.; Kang, S.C. Supplementation with Cynanchum wilfordii radix extract for 8 weeks lowers serum total cholesterol: A controlled, randomized, double-blind clinical trial. Phytother. Res. 2020, 34, 2313–2322. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
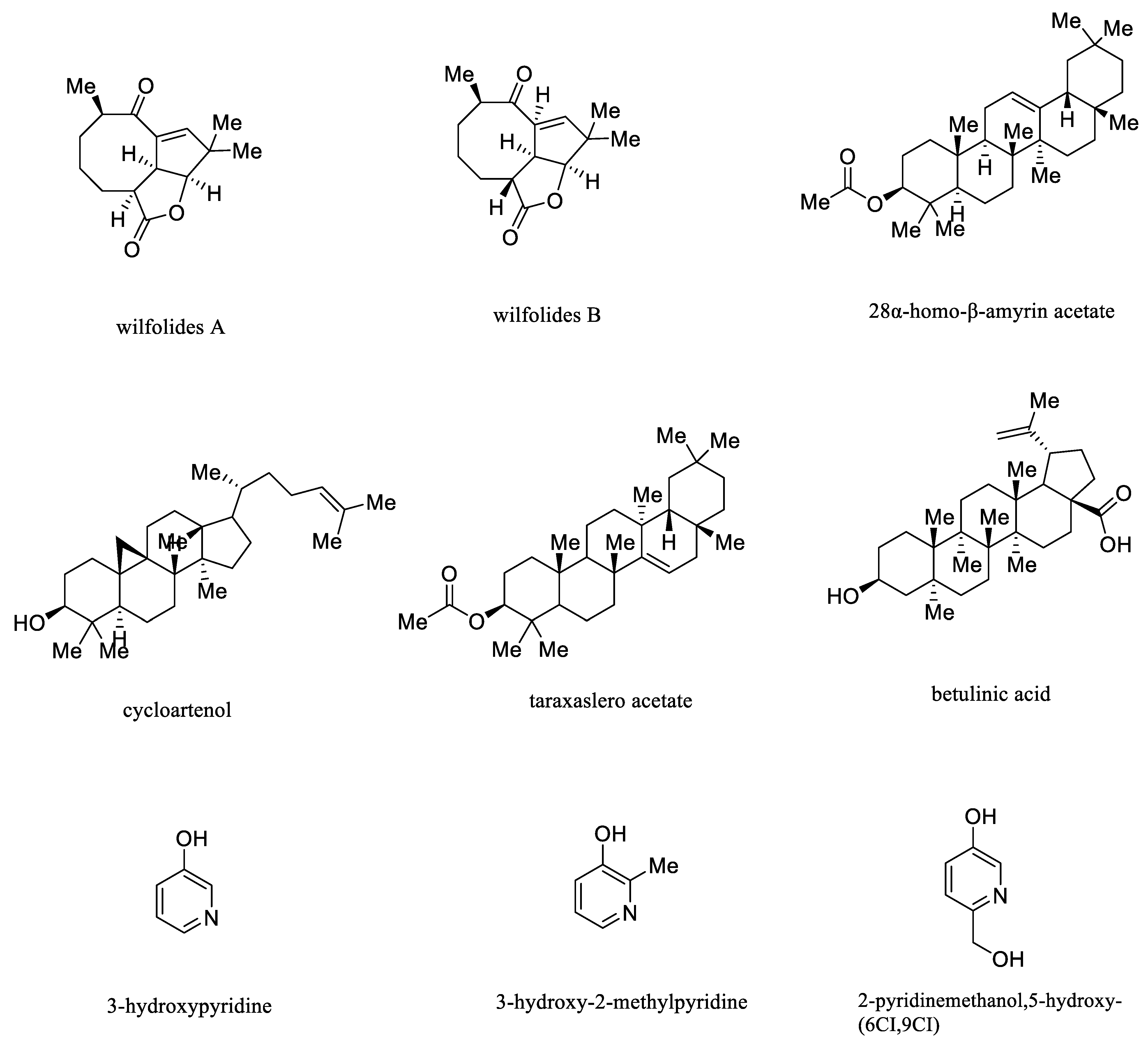{kind=link}
{kind=link}
{kind=link}
{kind=link}
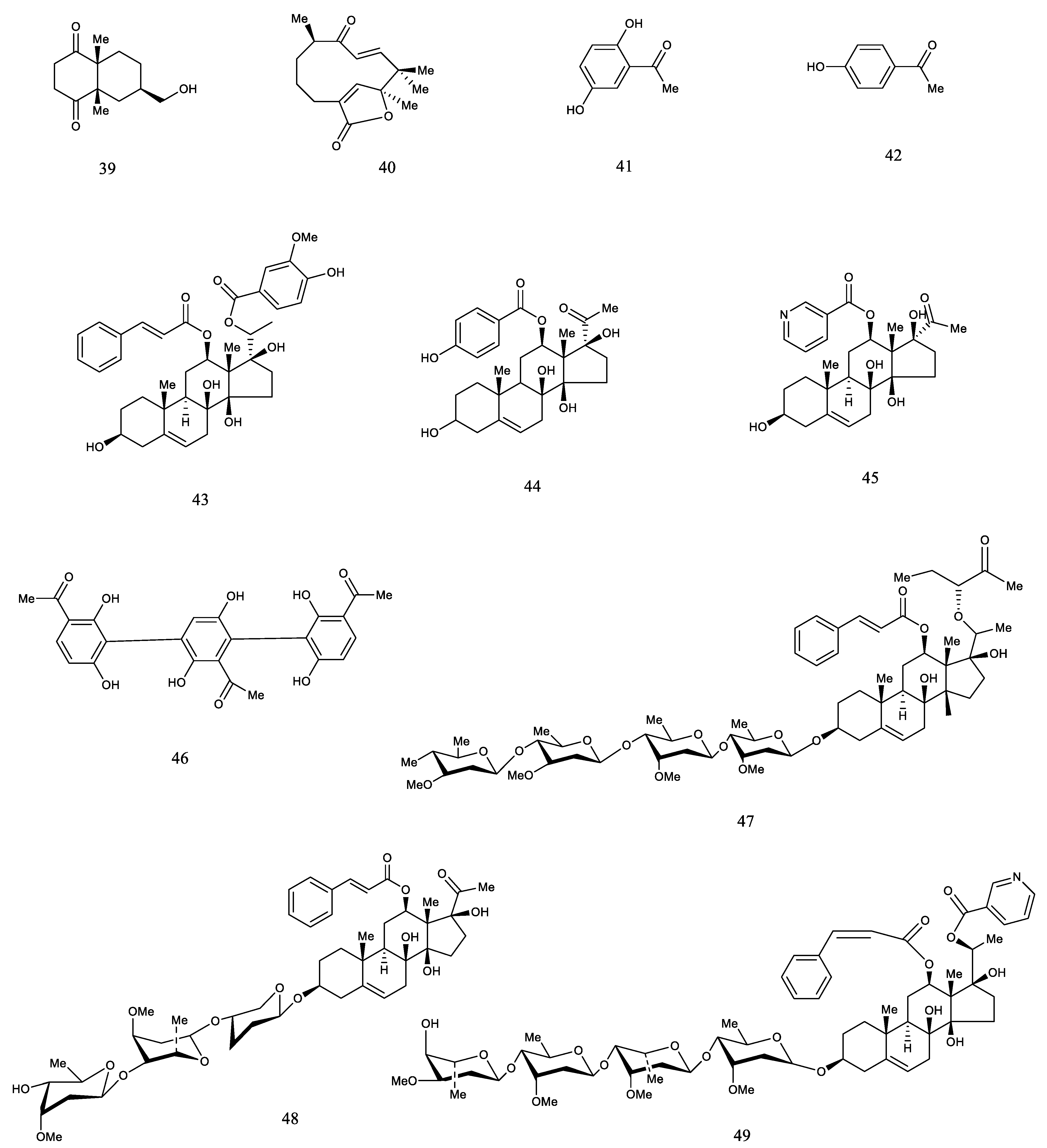{kind=link}
{kind=link}
| Number | Compounds | Species | References |
|---|---|---|---|
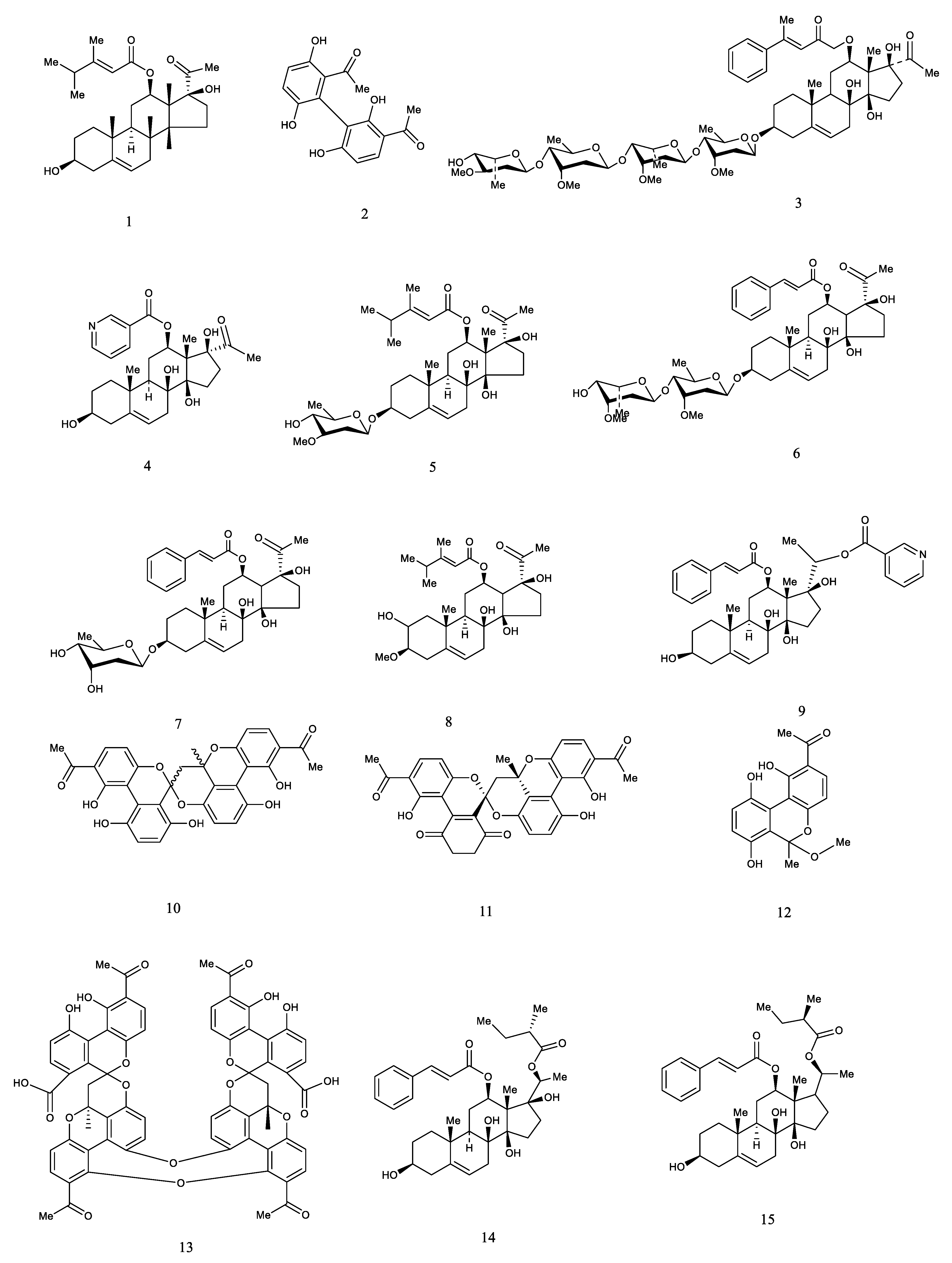
| 1 | Caudatin | CA CB CW | [25,26,27] |
| 2 | Qingyangshengenin | CA | [31] |
| 3 | Deacylmetaplexigenin | CA CW | [31,32] |
| 4 | Rostratamine | CA CW | [31,32] |
| 5 | Gagaminine | CA | [31] |
| 6 | Daucosterol | CA | [31] |
| 7 | Cautatin-3-O-β-d-digitoxo-pyranoside | CA | [31] |
| 8 | Caudatin-3-O-β-d-cymaropyranosyl-(1→4)-β-d-digitoxopyranoside | CA | [31] |
| 9 | Caudatin 3-O-β-d-cymaropyranosyl-(1→4)-β-d-cymaropyranoside | CA | [31] |
| 10 | Otophylloside B | CA | [31] |
| 11 | Caudatin 3-O-β-d-oleandropyranosyl-(1→4)-β-d-digitoxopyranosyl-(1→4)-β-d-cymaropyranoside | CA | [31] |
| 12 | Gagaminine3-O-α-l-cymropyranosyl-(1→4)-β-d-cymropyranosyl-(1→4)-β-d-cymropyranoside | CA | [31] |
| 13 | Auriculosides A | CA | [81] |
| 14 | Auriculosides B | CA | [81] |
| 15 | Cyanoauriculoside C | CA | [82] |
| 16 | Cyanoauriculoside D | CA | [82] |
| 17 | Cyanoauriculoside E | CA | [82] |
| 18 | Cynanauriculoside II | CA | [82] |
| 19 | Wilfoside K1N | CA | [82] |
| 20 | Auriculoside IV | CA | [82] |
| 21 | Kidjolanin | CA | [83] |
| 22 | 3-Formyloxymetaplexigenin | CA | [84] |
| 23 | Metaplexigenin | CA | [84] |
| 24 | Auriculoside I | CA | [33] |
| 25 | Auriculoside II | CA | [33] |
| 26 | Auriculoside III | CA | [33] |
| 27 | Wilfoside C1G | CA CW | [33] |
| 28 | Cynanauriculoside I | CA | [33] |
| 29 | Cynauricuoside A | CA | [33] |
| 30 | Gagaminin3-O-β-l-cymaropyranosyl-(1→4)-β-d-cymaropyranosyl-(1→4)-α-l-diginopyranosyl-(1→4)-β-d-digitoxopyranoside | CB | [85] |
| 31 | Gagaminin3-O-β-L-cymaropyranosyl-(1→4)-β-d-cymaropyranosyl-(1→4)-α-l-diginopyranosyl-(1→4)-β-d-cymaropyranoside | CB | [85] |
| 32 | 12-O-nicotinoylsarcostin3-O-β-l-cymaropyranosyl-(1→4)-β-d-cymaropyranosyl-(1→4)-α-l-diginopyranosyl-(1→4)-β-d-cymaropyranoside | CB | [85] |
| 33 | Penupogenin3-O-β-d-glucopyranosyl-(1→4)-β-l-cymaropyranosyl-(1→4)-β-d-cymaropyranosyl-(1→4)-α-l-diginopyranosyl-(1→4)-β-d-cymaropyranoside | CB | [85] |
| 34 | 12-O-acetylsarcostin3-O-β-l-cymaropyranosyl-(1→4)-β-d-cymaropyranosyl-(1→4)-β-l-cymaropyranosyl-(1→4)-β-d-digitoxopyranosyl-(1→4)-β-d-digitoxopyranoside | CB | [85] |
| 35 | 12-O-acetylsarcostin3-O-β-l-cymaropyranosyl-(1→4)-β-d-digitoxopyranosyl-(1→4)-β-l-cymaropyranosyl-(1→4)-β-d-cymaropyranosyl-(1→4)-α-l-diginopyranosy-(1→4)-β-d-cymaropyranoside | CB | [85] |
| 36 | Cynabungoside A | CB CW | [26,86] |
| 37 | Cynabungoside B | CB | [86] |
| 38 | Cynabungoside C | CB | [86] |
| 39 | Cynabungolide | CB | [86] |
| 40 | Cynabungone | CB | [86] |
| 41 | Wilfoside C1N | CB | [86] |
| 42 | Wilfolide A | CB | [86] |
| 43 | Deacylmetaplexigenin3-O-α-cymaropyranosyl-(1→4)-β-cymaropyranosyl-(1→4)-α-cymaropyranosyl-(1→4)-β-cymaropyranosyl-(1→4)-β-cymaropyranoside | CB | [86] |
| 44 | Cynanbungeinoside A | CB | [87] |
| 45 | Cynanbungeinoside B | CB | [87] |
| 46 | Cynanbungeinoside C | CB | [87] |
| 47 | Cynanbungeinoside D | CB | [87] |
| 48 | Cynanbungeinoside E | CB | [87] |
| 49 | Cynanbungeinoside F | CB | [87] |
| 50 | Wilfoside B | CW | [88] |
| 51 | Wilfoside C | CW | [88] |
| 52 | Wilfoside D | CW | [88] |
| 53 | Wilfoside E | CW | [88] |
| 54 | Wilfoside F | CW | [88] |
| 55 | Wilfoside G | CW | [88] |
| 56 | Wilfoside H | CW | [88] |
| 57 | 20-O-salicyl-kidjoranin | CW | [32] |
| 58 | 12β-O-(4-hydroxybenzoyl)-8β,14β,17β-trihydroxypregn2,5-diene-20-one | CW | [32] |
| 59 | 3-O-methyl-caudatin | CW | [32] |
| 60 | 20-O-(4-hydroxybenzoyl)-kidjoranin | CW | [32] |
| 61 | 20-O-vanilloyl-kidjoranin | CW | [32] |
| 62 | 12-O-vanilloyl-deacymetaplexigenin | CW | [32] |
| 63 | 17β-O-cinnamoyl-3β,8β,14β-trihydroxypregn-12,20-ether | CW | [32] |
| 64 | Deacymetaplexigenin | CW | [32] |
| 65 | 12-O-benzoyldeacymetaplexigenin | CW | [32] |
| 66 | 12β-O-benzoyl8β,14β,17β-trihydroxypregn-2,5-diene-20-one | CW | [32] |
| 67 | Sarcostin | CW | [32] |
| 68 | Deacylcynanchogenin | CW | [32] |
| 69 | Kidjoranine3-O-β-d-glucopyranosyl(1→4)-α-l-cymaropyranosyl(1→4)-β-d-cymaropyranosyl(1→4)-α-l-diginopyranosyl(1→4)-β-d-cymaropyranoside | CB | [89] |
| 70 | Caudatin3-O-β-d-glucopyranosyl(1→4)-α-l-cymaropyranosyl(1→4)-β-d-cymaropyranosyl(1→4)-α-l-diginopyranosyl(1→4)-β-d-cymaropyranoside | CB | [89] |
| 71 | Kidjoranine3-O-β-d-glucopyranosyl(1→4)-α-l-diginopyranosyl(1→4)-β-d-cymaropyranoside | CB | [89] |
| 72 | O-nicotinoyl-20-O-cinnamonacylsarcostin-O-β-d-glucopyranosyl(1→4)-α-l-cymaropyranosyl(1→4)-β-d-cymaropyranosyl(1→4)-α-l-diginopyranosyl(1→4)-β-d-digitoxopyranoside | CB | [89] |
| 73 | Penupogenin3-O-β-d-glucopyranosyl(1→4)-α-l-cymaropyranosyl(1→4)-β-d-cymaropyranosyl(1→4)-α-l-diginopyranosyl(1→4)-β-d-digitoxopyranoside | CB | [89] |
| 74 | Kidjoranine3-O-β-d-glucopyranosyl(1→4)-α-l-cymaropyranosyl(1→4)-β-d-cymaropyranosyl(1→4)-α-l-diginopyranosyl(1→4)-β-d-digitoxopyranoside | CB | [89] |
| 75 | Caudatin3-O-β-d-glucopyranosyl(1→4)-α-l-diginopyranosyl(1→4)-β-d-cymaropyranoside | CB | [89] |
| 76 | 12-O-benzoyl-deacetylmetaplexigenin3-O-β-d-glucopyranosyl(1→4)-α-l-cymaropyranosyl(1→4)-β-d-cymaropyranosyl(1→4)-α-L-diginopyranosyl(1→4)-β-d-cymaropyranosi-de | CB | [89] |
| 77 | Caudatin3-O-β-d-glucopyranosyl(1→4)-α-l-cymaropyranosyl(1→4)-β-d-cymaropyranosyl(1→4)-α-l-cymaropyranosyl(1→4)-β-d-cymaropyranosyl(1→4)-α-l-diginopyranosyl(1→4)-β-d-cymaropyranoside | CB | [89] |
| 78 | Caudatin3-O-α-l-cymaropyranosyl(1→4)-β-d-cymaropyranosyl(1→4)-α-l-diginopyranosyl(1→4)-β-d-cymaropyran-oside | CB | [89] |
| 79 | Caudatin3-O-α-l-diginopyranosyl(1→4)-β-d-cymaropyran-oside | CB | [89] |
| 80 | Kidjoranine3-O-β-d-cymaropyranosyl(1→4)-α-l-diginopyranosyl(1→4)-β-d-cymaropyranoside | CB | [89] |
| 81 | Kidjoranine3-O-α-l-diginopyranosyl(1→4)-β-d-cymaropyranoside | CB | [89] |
| 82 | Caudatin3-O-β-d-cymaropyranosyl(1→4)-α-l-diginopy-ranosyl(1→4)-β-d-cymaropyranoside | CB | [89] |
| 83 | O-benzoyl-deacetylmetaplexigenin | CB | [89] |
| 84 | 12-O-acetyl-20-O-(2-methylbutanoyl)-sarcostin | CB | [89] |
| 85 | 20-O-Acetyl-penupogenin | CB | [89] |
| 86 | Gracigenin | CB | [89] |
| 87 | 8,14-seco-caudatin | CB | [89] |
| 88 | Penupogenin | CB | [89] |
| 89 | (20S)-12-cinnamoyloxy-20-acetyloxy-3,5,17-trihydroxy-8,14-seco-5,17-pregn-6-ene-8,14-dione | CB | [89] |
| 90 | Isoikemagenin | CB | [89] |
| 91 | O-cinnamoyl-20-O-(S)-(+)-α-methylbutyryl–sarcostin | CB | [89] |
| 92 | 12-O-cinnamoyl-20-O-(R)-(-)-α-methylbutyrylsarcostin | CB | [89] |
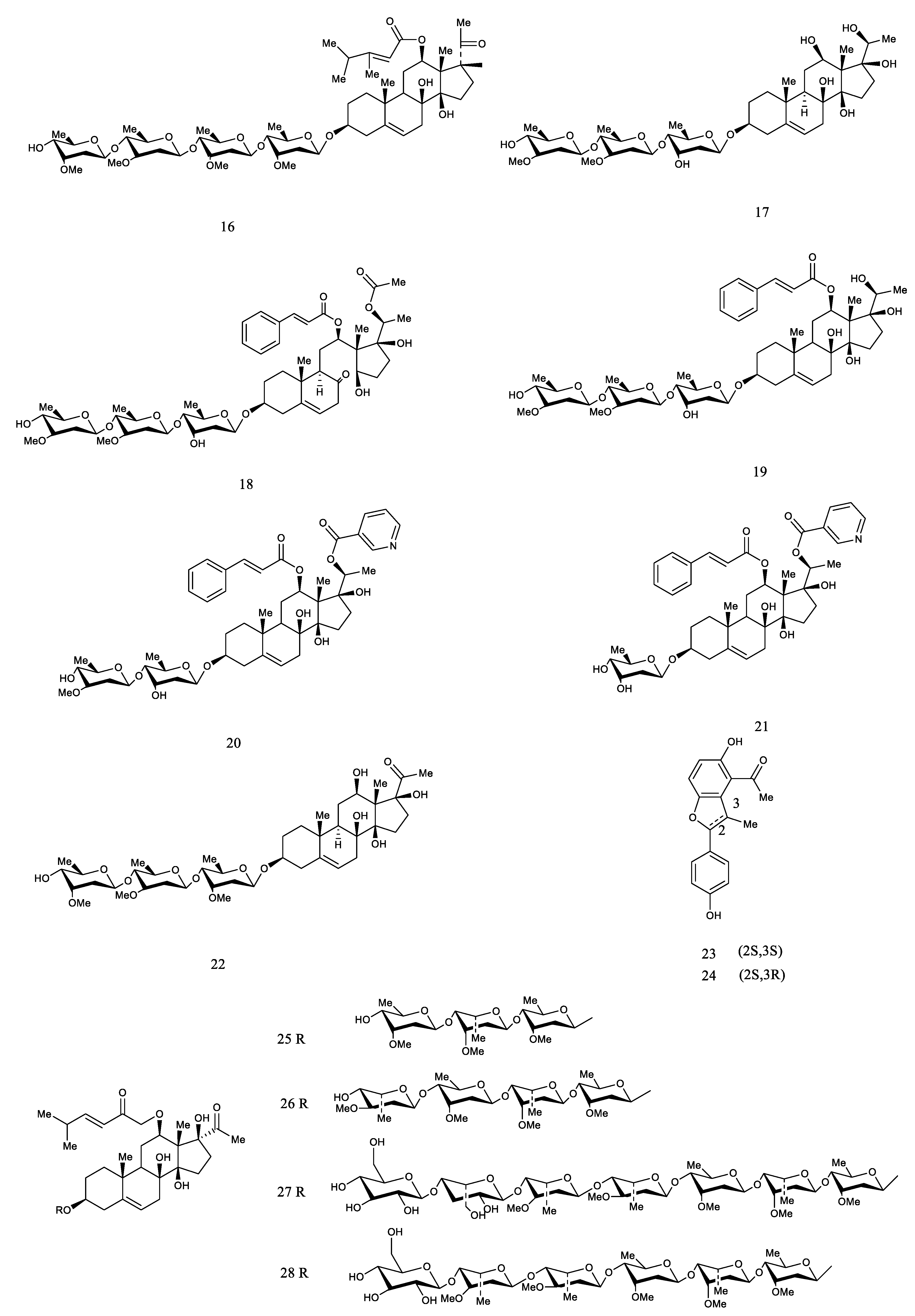
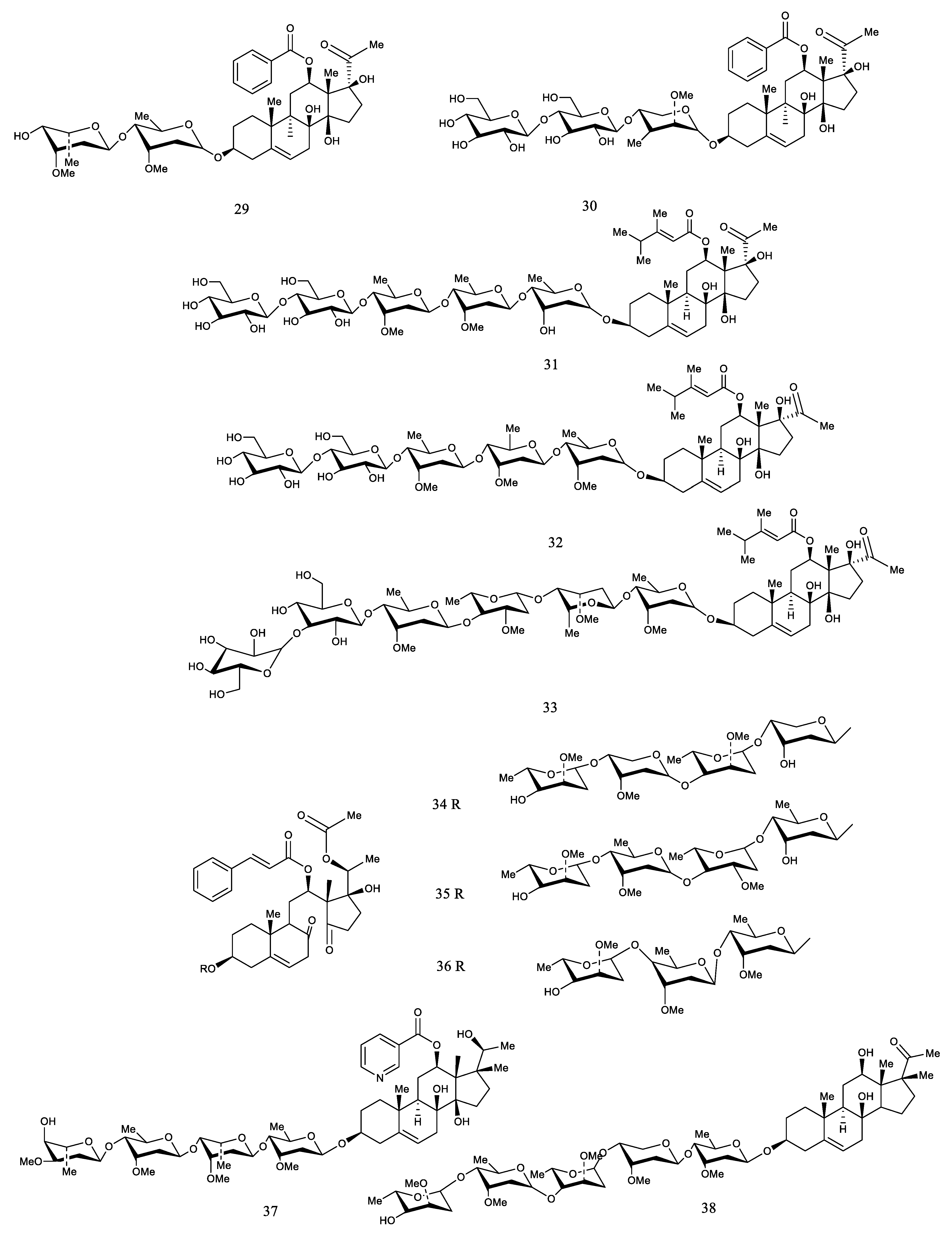
| 93 | Cynanauriculoside C | CA | [27] |
| 94 | Cynanauriculoside D | CA | [27] |
| 95 | Cynanauriculoside E | CA CW | [27,34] |
| 96 | Otophylloside L | CA | [27] |
| 97 | Wilfoside C3N | CA | [35] |
| 98 | Kidjoranin | CA CW | [35] |
| 99 | Cynanauriculoside I | CA | [35] |
| 100 | Cynanauriculoside II | CA | [35] |
| 101 | Cyanoauriculoside F | CA | [82] |
| 102 | Cyanoauriculoside G | CA | [82] |
| 103 | Cyanoauriculoside H | CA | [82] |
| 104 | 3-O-α-l-cymaropyranosyl-(1→4)-β-Dcymaropyranosyl-(1→4)-α-l-diginopyranosyl-(1→4)-β-d-cymaropyranoside | CA | [82] |
| 105 | Wilfoside D1N | CA | [82] |
| 106 | Cyanoauriculoside A | CA CW | [36] |
| 107 | Cyanoauriculoside B | CA | [36] |
| 108 | Cynsaccatol Q | CA | [90] |
| 109 | Saccatol K | CA | [90] |
| 110 | Cynanbungeigenin C | CB | [91] |
| 111 | Cynanbungeigenin D | CB | [91] |
| 112 | Wilfoside A | CW | [88] |
| 113 | Wilfoside C1G | CA | [92] |
| 114 | Cynauricuoside C | CA | [92] |
| 115 | Caudatin-2,6-dideoxy-3-O-methy-β-d-cymaropyranoside | CA | [92] |
| 116 | Kidjoranin3-O-αdiginopyranosyl-(1→4)-β-cymaropyranosi-de | CA | [64] |
| 117 | Kidjoranin 3-O-β-digitoxopyranoside | CA | [64] |
| 118 | Caudatin 3-O-β-cymaropyranoside | CA | [64] |
| 119 | Auriculoside A | CA | [93] |
| 120 | Wilfoside C1GG | CW | [26] |
| 121 | Wilfoside C1G | CW | [26] |
| 122 | Wilfoside K1GG | CW | [26] |
| 123 | Wilfoside M1N | CA CW | [34,37] |
| 124 | Cynauricuside A | CW | [37] |
| 125 | Cynauricoside B | CW | [37] |
| 126 | Cynauricoside C | CW | [37] |
| 127 | Cynauricoside D | CW | [37] |
| 128 | Cynauricoside E | CW | [37] |
| 129 | Cynauricoside F | CW | [37] |
| 130 | Cynauricoside G | CW | [37] |
| 131 | Cynauricoside H | CW | [37] |
| 132 | Cynauricoside I | CW | [37] |
| 133 | Eleutherosidea | CA | [38] |
| 134 | Caudatin-3-O-β-cymaropyranoside | CA | [59] |
| 135 | Cynauricuoside B | CA | [94] |
| 136 | Gagaminin-3-O-α-l-cymaropyranosyl-(1→4)-β-d-cymaropyranosyl-(1→4)-α-l-diginopyranosyl-(1→4)-β-d–digitoxopyranoside | CW | [74] |
| 137 | Cynawilfoside A | CW | [74] |
| 138 | Cynawilfoside B | CW | [74] |
| 139 | Cynawilfoside C | CW | [74] |
| 140 | Cynawilfoside D | CW | [74] |
| 141 | Cynawilfoside E | CW | [74] |
| 142 | Cynawilfoside F | CW | [74] |
| 143 | Cynawilfoside G | CW | [74] |
| 144 | Cynawilfoside H | CW | [74] |
| 145 | Cynawilfoside I | CW | [74] |
| 146 | Gagaminin-3-O-β-d-cymaropyranosyl-(1→4)-β-Doleandropyranosyl-(1→4)-β-d-cymaropyranosyl-(1→4)-β-d–cymaropyranosid | CW | [95] |
| 147 | Lneolon | CW | [96] |
| 148 | Saccatol D | CA | [97] |
| 149 | Saccatol E | CA | [97] |
| 150 | Saccatol F | CA | [97] |
| 151 | Saccatol G | CA | [97] |
| 152 | Saccatol H | CA | [97] |
| 153 | Saccatol I | CA | [97] |
| 154 | Saccatol J | CA | [97] |
| 155 | Saccato K | CA | [97] |
| 156 | Cynsaccatol I | CA | [97] |
| 157 | Cynsaccatol J | CA | [97] |
| 158 | Cynsaccatol K | CA | [97] |
| 159 | Cynsaccatol L | CA | [97] |
| 160 | Cynsaccatol M | CA | [97] |
| 161 | Cynsaccatol N | CA | [97] |
| 162 | Cynsaccatol O | CA | [97] |
| 163 | Cynsaccatol P | CA | [97] |
| 164 | Cynsaccatol Q | CA | [97] |
| 165 | Cynsaccatol R | CA | [97] |
| 166 | Cynsaccatol S | CA | [97] |
| 167 | Cynsaccatol T | CA | [97] |
| 168 | Cynsaccatol U | CA | [97] |
| 169 | Cynsaccatol V | CA | [97] |
| 170 | Cynsaccatol W | CA | [97] |
| 171 | Gagamine | CA | [98] |
| Number | Compounds | Species | References |
|---|---|---|---|
| 172 | 2,4-Dihydroxyacetophenone | CA CB CW | [24,28] |
| 173 | 2,5-Dihydroxyacetophenone | CA CB CW | [24,28] |
| 174 | 4-Hydroxyacetophenone | CA CB CW | [24,28] |
| 175 | Cynandione A | CA CW | [28,30] |
| 176 | Cynandione B | CA CW | [39] |
| 177 | Cynandione E | CA CW | [39] |
| 178 | Baishouwubenzophenone | CA CB CW | [24,29] |
| 179 | Cynwilforone A | CW | [99] |
| 180 | Cynwilforone B | CW | [99] |
| 181 | Cynwilforone C | CW | [99] |
| 182 | 2-O-β-laminaribiosyl-4-hydroxyacetophenone | CB CW | [42,43] |
| 183 | Bungeiside-C | CB CW | [42,43] |
| 184 | Bungeiside-D | CB CW | [42,43] |
| 185 | (+) Cynwilforone D | CW | [100] |
| 186 | (-) Cynwilforone D | CW | [100] |
| 187 | (+) Cynwilforone E | CW | [100] |
| 188 | (-) Cynwilforone E | CW | [100] |
| 189 | (+) Cynwilforone F | CW | [100] |
| 190 | (-) Cynwilforone F | CW | [100] |
| 191 | Cynwilforone G | CW | [100] |
| 192 | Bungeiside-A | CB | [101] |
| 193 | Bungeiside-B | CB | [101] |
| 194 | Picein | CA | [15] |
| 195 | 4′-Hydroxy-3′-methoxyacetophenone | CW | [40] |
| 196 | 1-(2-Hydroxy-4,5-dimethoxyphenyl) ethanone | CA | [102] |
| 197 | Cynanchone A | CW | [103] |
| 198 | 2,4-Dihydroxy-5-methoxyacetophenone | CA | [104] |
| 199 | Cynantetrone | CA | [105] |
| Number | Compounds | Species | References |
|---|---|---|---|
| 200 | β-sitosterol | CA | [38] |
| 201 | β-amyrin acetate | CA | [38] |
| 202 | Wilfolides A | CW | [74] |
| 203 | Wilfolides B | CW | [74] |
| 204 | Cycloartenol | CA | [13] |
| 205 | 28α-Homo-β-amyrin acetate | CA | [13] |
| 206 | 11α, 12α-Epoxytaraxer-14-en-3β-yl-acetate | CA | [13] |
| 207 | δ-Amyrine acetate | CA | [13] |
| 208 | Taraxaslero acetate | CA | [30] |
| 209 | Betulinic acid | CA | [30] |
| 210 | Oleanolic acid | CA | [106] |
| 211 | Lupeol | CB | [107] |
| Number | Compounds | Species | References |
|---|---|---|---|
| 212 | 3-Hydroxypyridine | CA | [15] |
| 213 | 3-Hydroxy-2-methylpyridine | CA | [15] |
| 214 | 2-Pyridinemethanol, 5-hydroxy | CA | [15] |
| 215 | 1H-imidazole-5-carboxylic acid | CA | [13] |
| 216 | 6-[(β-d-xylopyranosyl) methyl]-3-pyridinol | CA | [16] |
| 217 | 2-Methyl-6-(2′,3′,4′-trihydroxybutyl)-pyrazine | CA | [15] |
| Number | Compounds | Species | References |
|---|---|---|---|
| 218 | Ferulic acid methylester | CA | [38] |
| 219 | Islariciresinol | CA | [15] |
| 220 | Vomifoliol | CA | [15] |
| 221 | n-Hexacos-5,8,11-trienoic acid | CA | [13] |
| 222 | Isocopoletin | CA | [15] |
| 223 | Isofraxidin | CA | [15] |
| 224 | Adenosine | CA | [15] |
| 225 | (+)-isolariciresinol | CA | [15] |
| 226 | 4,4-dimethyl heptanedioic acid | CA | [15] |
| 227 | Leucanthemitol | CA CW | [5,41] |
| 228 | Suceinie acid | CA CW | [5,41] |
| 229 | Sucrose | CA CW | [38,40] |
| 230 | Methyleugenol | CA CW | [5] |
| 231 | Conduritol F | CW | [5] |
| 232 | 3-(β-d-ribofuranosyl)-2,3-dihydro-6H-1,3-oxazine-2,6-dione | CW | [14] |
| Number | Bioactive Constituents and Parts | Species | Pharmacology | References |
|---|---|---|---|---|
| 1 | Caudatin | CW CA CA CB CB CA | Anti-hepatocellular carcinoma Anti-breast cancer Anti-uterine cancer Anti-human glioma Anti-gastric cancer Anti-lung cancer | [136] [110] [62] [111] [112] [115] |
| 2 | Cynandione A | CA CW CW CW CW CW | Neuroprotection Hepatoprotection Anti-inflammatory Reducing liver lipid Reducing blood lipid Hypoglycemic | [57] [58] [137] [70] [71] [99] |
| 3 | Wilfoside KIN | CW CW CA | Anti-angiogenic Anti-epileptic Appetite suppression | [130] [74] [37] |
| 4 | Aauriculoside A | CA | Anti-breast cancer | [93] |
| 5 | Caudatin-2,6-dideoxy-3-O-methy-β-d-cymaropyranoside | CA | Anti-hepatocellular carcinoma | [92] |
| 6 | Kidjoranin-3-O-α-diginopyranosyl-(1→4)-β-cymaropyranoside | CA | Anti-cervical cancer Anti-hepatocellular carcinoma Anti-breast cancer | [64] |
| 7 | Kidjoranin-3-O-β-digitoxopyranoside | CA | Anti-cervical cancer Anti-hepatocellular carcinoma Anti-breast cancer | [64] |
| 8 | Caudatin-3-O-β-cymaropyranoside | CA | Anti-cervical cancer Anti-hepatocellular carcinoma Anti-breast cancer | [64] |
| 9 | Gagaminine | CW | Antioxidant | [63] |
| 10 | Cynandione B | CA | Antioxidant | [105] |
| 11 | Cynandione E | CA | Antioxidant | [105] |
| 12 | Cynanchone A | CA | Antioxidant | [105] |
| 13 | Cynantetrone | CA | Antioxidant | [105] |
| 14 | Cynanbungeigenin C | CB | Anti-human glioma | [91] |
| 15 | Cynanbungeigenin D | CB | Anti-human glioma | [91] |
| 16 | Caudatin-3-O-β-d-cymaropyranosyl-(1→4)-β-d-oleandropyranosyl-(1→4)-β-d-cymaropyranosyl-(1→4)-β-d-cymaropyranoside | CA | Anti-gastric cancer | [113] |
| 17 | Cynsaccatol Q | CA | Neuroprotection | [90] |
| 18 | Saccatol K | CA | Neuroprotection | [90] |
| 19 | Cynsaccatol I | CA | Neuroprotection | [97] |
| 20 | Cynsaccatol N | CA | Neuroprotection | [97] |
| 21 | Cynsaccatol O | CA | Neuroprotection | [97] |
| 22 | Cynsaccatol S | CA | Neuroprotection | [97] |
| 23 | (+) Cynwiforones F | CW | Neuroprotection | [100] |
| 24 | (−) Cynwiforones F | CW | Neuroprotection | [100] |
| 25 | Wilfoside C3N | CA | Anti-esophageal cancer | [114] |
| 26 | Wilfoside C1N | CW CB CW CA | Antifungal Immunoregulation Anti-epileptic Appetite suppression | [26] [86] [74] [37] |
| 27 | Wilfoside C1G | CW | Antifungal | [26] |
| 28 | Wilfoside C1GG | CW | Antifungal | [26] |
| 29 | Cynanauriculoside C | CA | Antidepressant | [27] |
| 30 | Cynanauriculoside D | CA | Antidepressant | [27] |
| 31 | Cynanauriculoside E | CA | Antidepressant | [27] |
| 32 | Otophylloside L | CA | Antidepressant | [27] |
| 33 | Cynauricuoside C | CA | Antidepressant | [27] |
| 34 | Cynabungoside A | CB | Immunoregulation | [86] |
| 35 | Cynabungoside B | CB | Immunoregulation | [86] |
| 36 | Cynabungoside C | CB | Immunoregulation | [86] |
| 37 | 12-O-Nicotinoylsarcostin-3-O-β-lcymaropyranosyl -(1→4)-β-d-cymaropyranosyl-(1→4)-α-l-diginopyranosyl-(1→4)-β-d-cymaropyranoside | CB | Immunoregulation | [86] |
| 38 | Deacylmetaplexigenin-3-O-α-cymaropyranosyl-(1→4) -β-cymaropyranosyl-(1→4)-α-cymaropyranosyl-(1→4)-β-cymaropyranosyl-(1→4)-β-cymaropyranoside | CB | Immunoregulation | [86] |
| 39 | Cynabungone | CB | Immunoregulation | [86] |
| 40 | Cynabungolide | CB | Immunoregulation | [86] |
| 41 | 2,5-Dihydroxyacetophenone | CB CW | Skin protection Reducing blood lipid | [121] [71] |
| 42 | 4-Hydroxyacetophenone | CW | Anti-inflammatory | [125] |
| 43 | 20-O-Salicyl-kidjoranin | CW | Anti-leukemic | [32] |
| 44 | Qingyangshengenin | CW | Anti-leukemic | [32] |
| 45 | Rostratamin | CW | Anti-leukemic | [32] |
| 46 | Cynwilforone A | CW | Hypoglycemic | [99] |
| 47 | Cynawilfoside A | CW | Anti-epileptic | [74] |
| 48 | Cynauricoside A | CW | Anti-epileptic | [74] |
| 49 | Cyanoauriculoside G | CW | Anti-epileptic | [74] |
| 50 | C21 steroidal glycoside | CA CA | Hepatoprotection Anti-inflammatory | [117] [126] |
| 51 | Polysaccharides | CW CW CA CA CA | Anti-inflammatory Antioxidant Menopause suppression Hepatoprotection Immunoregulation | [79] [80] [132] [118] [78] |
| 52 | Ethanol extract | CW CA CW CW CA CB CW CW | Anti-hepatocellular carcinoma Reducing blood lipid Gastric protection Reducing liver lipid Antiviral effect | [25,109] [123] [119] [122] [129] |
| 53 | Water extract | CW CW CW | Anti-prostatic Hyperplasia Aphrodisiac Bone-strengthening | [133] [131] [134] |
| 54 | Methanol extract | CA | Antidepressant activity | [127] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Cai, F.; Zhao, W.; Tian, J.; Kong, D.; Sun, X.; Liu, Q.; Chen, Y.; An, Y.; Wang, F.; et al. Cynanchum auriculatum Royle ex Wight., Cynanchum bungei Decne. and Cynanchum wilfordii (Maxim.) Hemsl.: Current Research and Prospects. Molecules 2021, 26, 7065. https://doi.org/10.3390/molecules26237065
Wang L, Cai F, Zhao W, Tian J, Kong D, Sun X, Liu Q, Chen Y, An Y, Wang F, et al. Cynanchum auriculatum Royle ex Wight., Cynanchum bungei Decne. and Cynanchum wilfordii (Maxim.) Hemsl.: Current Research and Prospects. Molecules. 2021; 26(23):7065. https://doi.org/10.3390/molecules26237065
Chicago/Turabian StyleWang, Lu, Fujie Cai, Wei Zhao, Jinli Tian, Degang Kong, Xiaohui Sun, Qing Liu, Yueru Chen, Ying An, Fulin Wang, and et al. 2021. "Cynanchum auriculatum Royle ex Wight., Cynanchum bungei Decne. and Cynanchum wilfordii (Maxim.) Hemsl.: Current Research and Prospects" Molecules 26, no. 23: 7065. https://doi.org/10.3390/molecules26237065
APA StyleWang, L., Cai, F., Zhao, W., Tian, J., Kong, D., Sun, X., Liu, Q., Chen, Y., An, Y., Wang, F., Liu, X., Wu, Y., & Zhou, H. (2021). Cynanchum auriculatum Royle ex Wight., Cynanchum bungei Decne. and Cynanchum wilfordii (Maxim.) Hemsl.: Current Research and Prospects. Molecules, 26(23), 7065. https://doi.org/10.3390/molecules26237065