
Schisandrin B Attenuates Hepatic Stellate Cell Activation and Promotes Apoptosis to Protect against Liver Fibrosis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
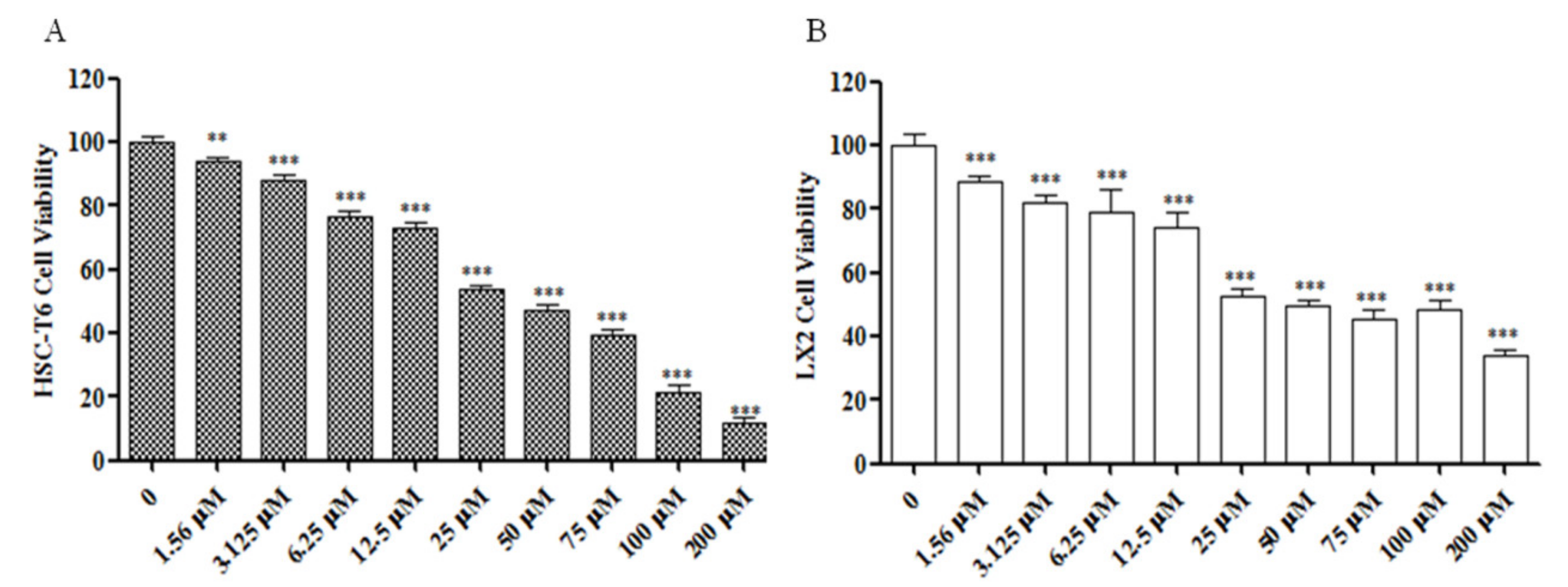
2.1. Sch B Inhibits the Proliferation of Activated Hepatic Stellate Cells
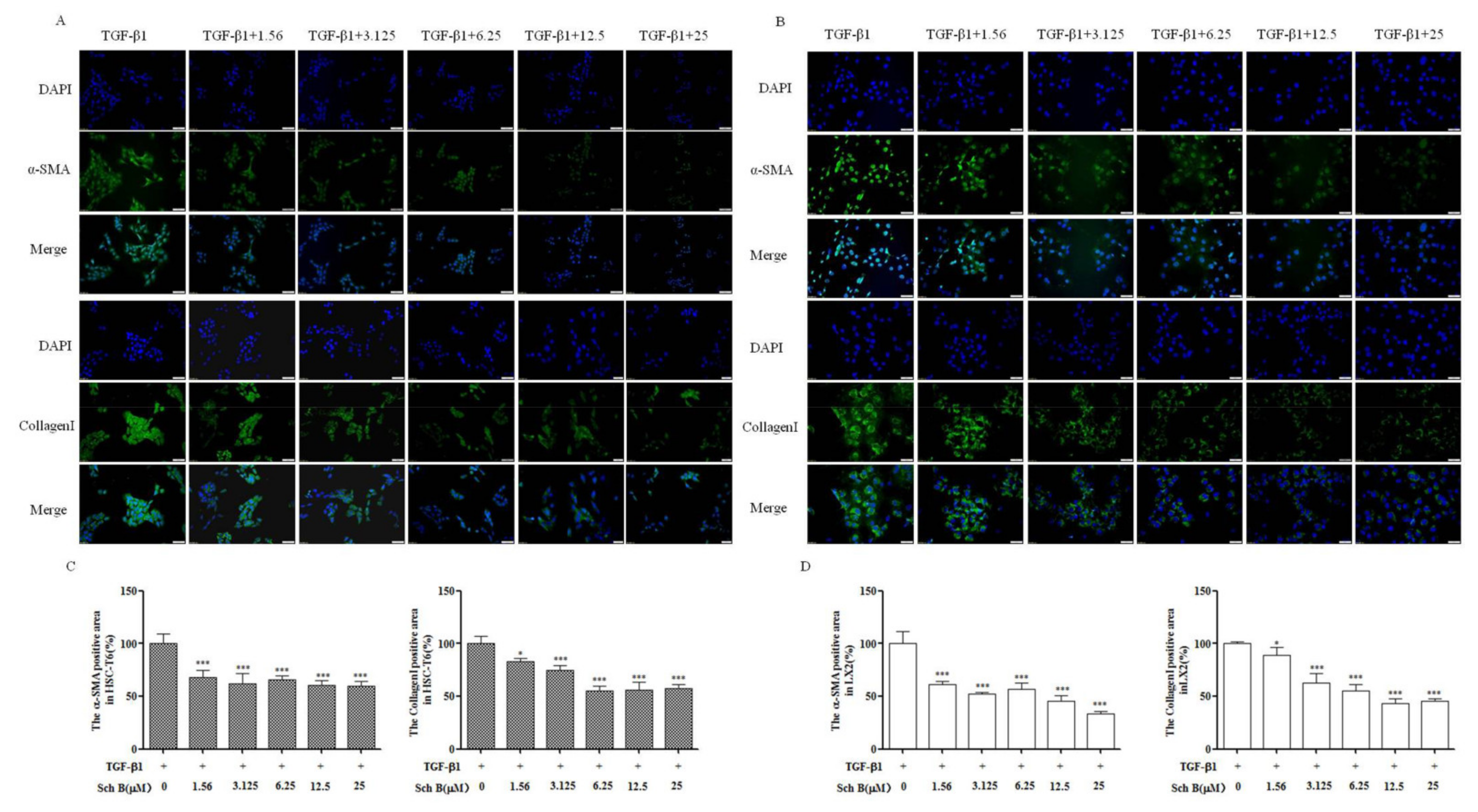
2.2. Sch B Attenuated the Secretion of Profibrotic Proteins by TGF-β1-Activated HSCs
2.3. Sch B Suppressed α-SMA and Collagen I Protein Expression by TGF-β1-Activated HSCs
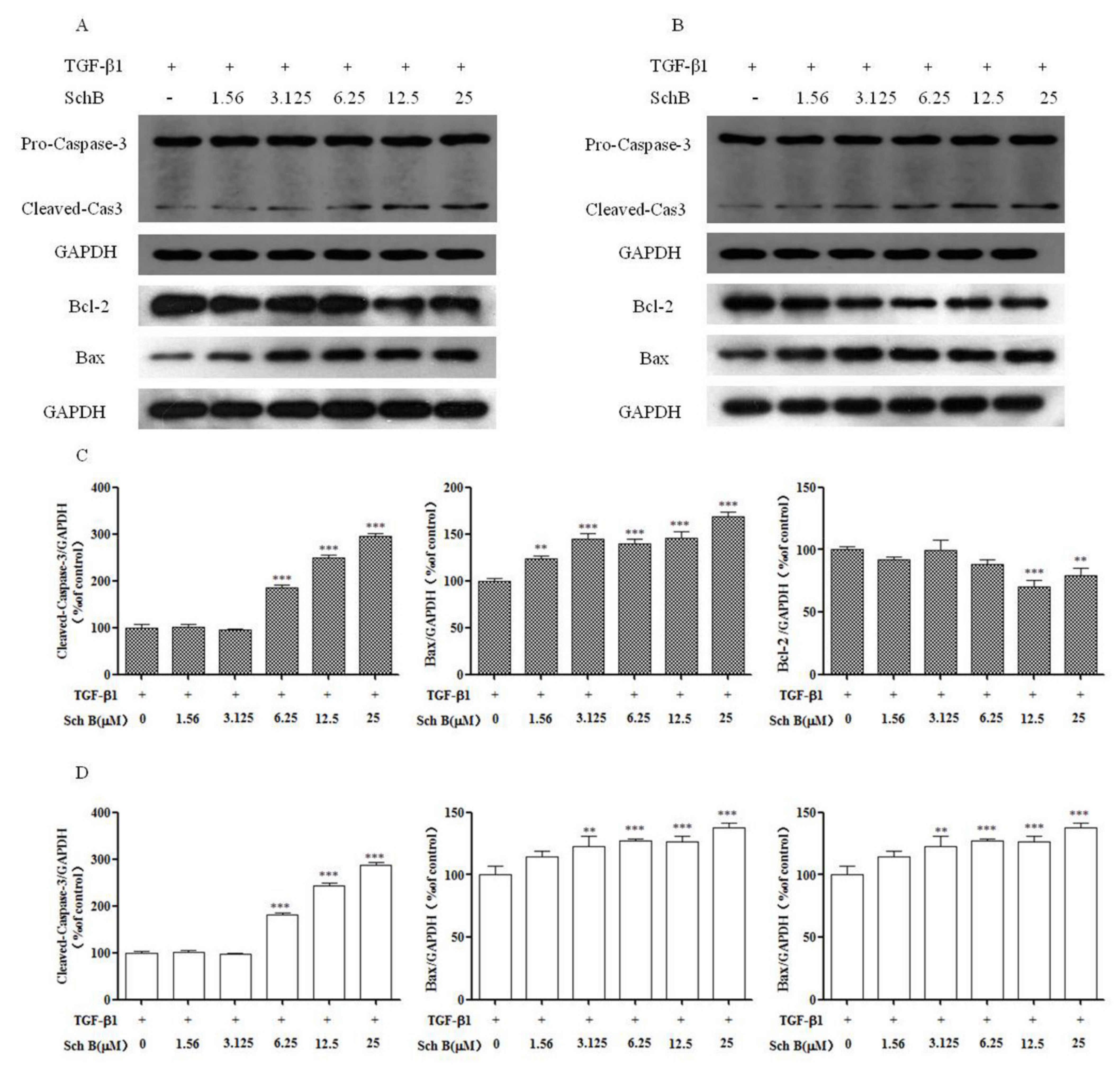
2.4. Sch B Causes Progressive HSCs Apoptosis
2.5. Sch B Promotes Apoptotic Factor Expression of Activated HSCs
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Preparation of Sch B Solutions
4.3. Cell Culture
4.4. CCK-8 Determination
4.5. Western Blotblotting
4.6. Flow Cytometry Analysis
4.7. Immunofluorescence Staining
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability Statement
References
- Koyama, Y.; Xu, J.; Liu, X.; Brenner, D.A. New developments on the treatment of liver fibrosis. Digest Dis. 2016, 34, 589–596. [Google Scholar] [CrossRef] [Green Version]
- Roeb, E. Matrix metalloproteinases and liver fibrosis (translational aspects). Matrix Biol. 2018, 68–69, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Asrani, S.K.; Devarbhavi, H.; Eaton, J.; Kamath, P.S. Burden of liver diseases in the world. J. Hepatol. 2019, 70, 151–171. [Google Scholar] [CrossRef] [PubMed]
- Gur, C.; Doron, S.; Kfir-Erenfeld, S.; Horwitz, E.; Abu-Tair, L.; Safadi, R.; Mandelboim, O. NKp46-mediated killing of human and mouse hepatic stellate cells attenuates liver fibrosis. Gut 2012, 61, 885–893. [Google Scholar] [CrossRef] [PubMed]
- Koyama, Y.; Brenner, D.A. Liver inflammation and fibrosis. J. Clin. Investig. 2017, 127, 55–64. [Google Scholar] [CrossRef]
- Higashi, T.; Friedman, S.L.; Hoshida, Y. Hepatic stellate cells as key target in liver fibrosis. Adv. Drug Deliv. Rev. 2017, 121, 27–42. [Google Scholar] [CrossRef]
- Lee, Y.A.; Wallace, M.C.; Friedman, S.L. Pathobiology of liver fibrosis: A translational success story. Gut 2015, 64, 830–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meindl-Beinker, N.M.; Matsuzaki, K.; Dooley, S. TGF-β signaling in onset and progression of hepatocellular carcinoma. Digest Dis. 2012, 30, 514–523. [Google Scholar] [CrossRef] [PubMed]
- Kisseleva, T.; Cong, M.; Paik, Y.; Scholten, D.; Jiang, C.; Benner, C.; Iwaisako, K.; Moore-Morris, T.; Scott, B.; Tsukamoto, H.; et al. Myofibroblasts revert to an inactive phenotype during regression of liver fibrosis. Proc. Natl. Acad. Sci. USA 2012, 109, 9448–9453. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.W.; LI, R.F.; Hu, L.J.; Wen, Z.Q.; Geng, D.M.; Wang, Y.M. Research on health care functions of domestic food. China J. Tradit. Chin. Med. Pharm. 2016, 31, 4749–4751. [Google Scholar]
- Dai, Z.; Xin, H.; Fu, Q.; Hao, H.; Li, Q.; Liu, Q.; Jin, Y. Exploration and optimization of conditions for quantitative analysis of lignans in Schisandra chinensis by an online supercritical fluid extraction with supercritical fluid chromatography system. J. Sep. Sci. 2019, 42, 2444–2454. [Google Scholar] [CrossRef] [PubMed]
- Yuan, F.; Liang, X.; Chen, X.; Qin, X.; Tan, C.; Wang, L. CYP2C19 is involved in the effect of Wuzhi tablet (Schisandra sphenanthera extract) and its constituents on the pharmacokinetics of intravenous voriconazole. Die Pharm. 2020, 75, 559–564. [Google Scholar]
- Ma, Z.; Xu, G.; Shen, Y.; Hu, S.; Lin, X.; Zhou, J.; Zhao, W.; Liu, J.; Wang, J.; Guo, J. Schisandrin B-mediated TH17 cell differentiation attenuates bowel inflammation. Pharmacol. Res. 2021, 166, 105459. [Google Scholar] [CrossRef]
- Xu, J.B.; Gao, G.C.; Yuan, M.J.; Huang, X.; Zhou, H.Y.; Zhang, Y.; Zheng, Y.X.; Wu, Z.; Feng, J.M.; Wu, J.M. Lignans from Schisandra chinensis ameliorate alcohol and CCl4-induced long-term liver injury and reduce hepatocellular degeneration via blocking ETBR. J. Ethnopharmacol. 2020, 258, 112813. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Sun, J.; Fan, X.; Guan, L.; Li, D.; Zhou, Y.; Zeng, X.; Chen, Y.; Zhang, H.; Xu, L.; et al. Schisandrol B promotes liver regeneration after partial hepatectomy in mice. Eur. J. Pharmacol. 2018, 818, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Lai, Q.; Luo, Z.; Wu, C.; Lai, S.; Wei, H.; Li, T.; Wang, Q.; Yu, Y. Attenuation of cyclosporine A induced nephrotoxicity by schisandrin B through suppression of oxidative stress, apoptosis and autophagy. Int. Immunopharmacol. 2017, 52, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, H.; Ma, R.; Wang, L. Schisandrin B inhibits epithelial-mesenchymal transition and stemness of large-cell lung cancer cells and tumorigenesis in xenografts via inhibiting the NF-κB and p38 MAPK signaling pathways. Oncol. Rep. 2021, 45, 115. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.N.; Zhang, C.Y.; Jin, X.D.; Li, Y.Z.; Zheng, X.Z.; Li, L. Inhibitory effect of schisandrin B on gastric cancer cells in vitro. World J. Gastroenterol. 2007, 13, 6506–6511. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Lu, C.; Liu, Z.; Zhang, P.; Guo, H.; Wang, T. Schizandrin B protects LPS-induced sepsis via TLR4/NF-κB/MyD88 signaling pathway. Am. J. Transl. Res. 2018, 10, 1155–1163. [Google Scholar] [PubMed]
- Novo, E.; Cannito, S.; Paternostro, C.; Bocca, C.; Miglietta, A.; Parola, M. Cellular and molecular mechanisms in liver fibrogenesis. Arch. Biochem. Biophys. 2014, 548, 20–37. [Google Scholar] [CrossRef] [PubMed]
- Wynn, T.A. Cellular and molecular mechanisms of fibrosis. J. Pathol. 2008, 214, 199–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meindl-Beinker, N.M.; Dooley, S. Transforming growth factor-beta and hepatocyte transdifferentiation in liver fibrogenesis. J. Gastroenterol. Hepatol. 2008, 23, S122–S227. [Google Scholar] [CrossRef]
- Faouzi, S.; Le Bail, B.; Neaud, V.; Boussarie, L.; Saric, J.; Bioulac-Sage, P.; Balabaud, C.; Rosenbaum, J. Myofibroblasts are responsible for collagen synthesis in the stroma of human hepatocellular carcinoma: An in vivo and in vitro study. J. Hepatol. 1999, 30, 275–284. [Google Scholar] [CrossRef]
- Benyon, R.C.; Arthur, M.J. Extracellular matrix degradation and the role of hepatic stellate cells. Semin. Liver Dis. 2001, 21, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Rippe, R.A. Life or death: The fate of the hepatic stellate cell following hepatic injury. Hepatology 1998, 27, 1447–1448. [Google Scholar] [CrossRef]
- Cao, X.H.; Zhao, S.S.; Liu, D.Y.; Wang, Z.; Niu, L.L.; Hou, L.H.; Wang, C.L. ROS-Ca2+ is associated with mitochondria permeability transition pore involved in surfactin-induced MCF-7 cells apoptosis. Chem. Biol. Interact. 2011, 190, 1627. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Youle, R.J. The role of mitochondria in apoptosis. Annu. Rev. Genet. 2009, 43, 95–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desagher, S.; Martinou, J.C. Mitochondria as the central control point of apoptosis. Trends Cell Biol. 2000, 10, 369–377. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Zhao, L.; Xia, Y.; Chen, J.; Hua, M.; Sun, Y. Schisandrin B Attenuates Hepatic Stellate Cell Activation and Promotes Apoptosis to Protect against Liver Fibrosis. Molecules 2021, 26, 6882. https://doi.org/10.3390/molecules26226882
Li Z, Zhao L, Xia Y, Chen J, Hua M, Sun Y. Schisandrin B Attenuates Hepatic Stellate Cell Activation and Promotes Apoptosis to Protect against Liver Fibrosis. Molecules. 2021; 26(22):6882. https://doi.org/10.3390/molecules26226882
Chicago/Turabian StyleLi, Zhiman, Lijuan Zhao, Yunshi Xia, Jianbo Chen, Mei Hua, and Yinshi Sun. 2021. "Schisandrin B Attenuates Hepatic Stellate Cell Activation and Promotes Apoptosis to Protect against Liver Fibrosis" Molecules 26, no. 22: 6882. https://doi.org/10.3390/molecules26226882
APA StyleLi, Z., Zhao, L., Xia, Y., Chen, J., Hua, M., & Sun, Y. (2021). Schisandrin B Attenuates Hepatic Stellate Cell Activation and Promotes Apoptosis to Protect against Liver Fibrosis. Molecules, 26(22), 6882. https://doi.org/10.3390/molecules26226882