
Prunetin 4′-O-Phosphate, a Novel Compound, in RAW 264.7 Macrophages Exerts Anti-Inflammatory Activity via Suppression of MAP Kinases and the NFκB Pathway

 , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
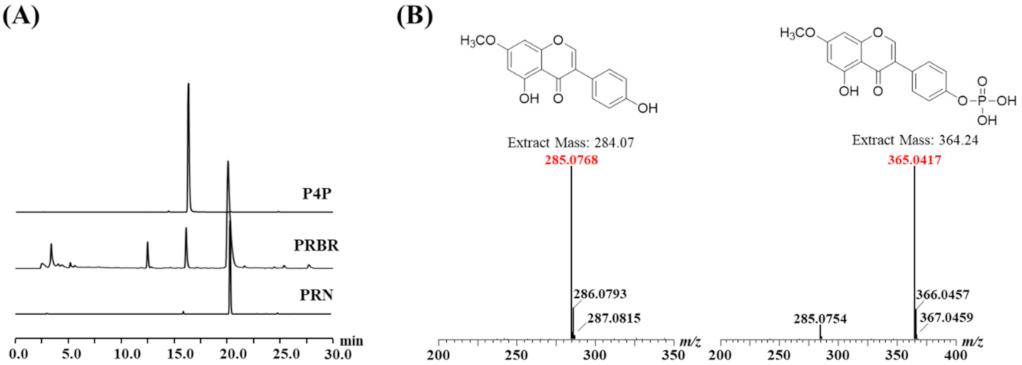
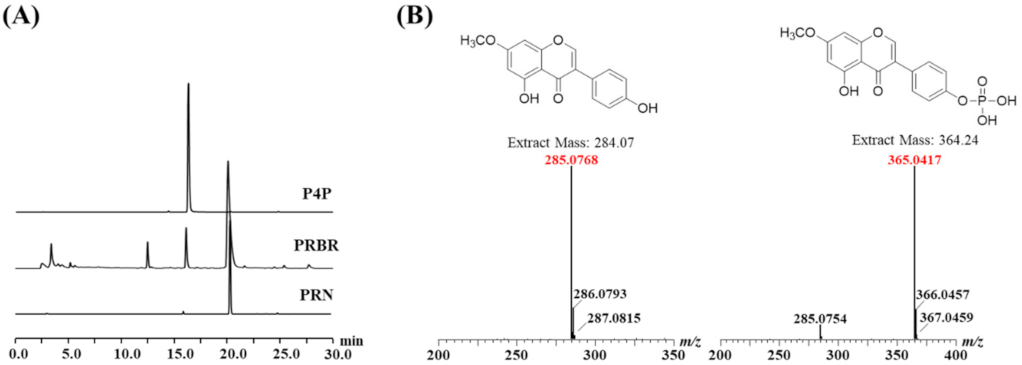
2.1. Chemical Structures of the Biorenovation Products of PRN
2.2. NMR Results
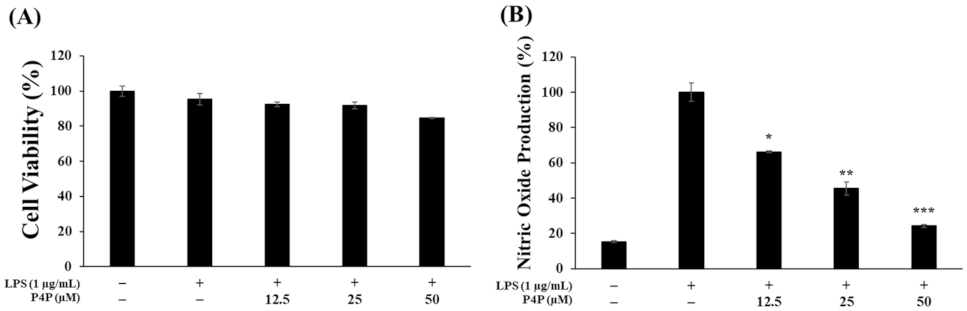
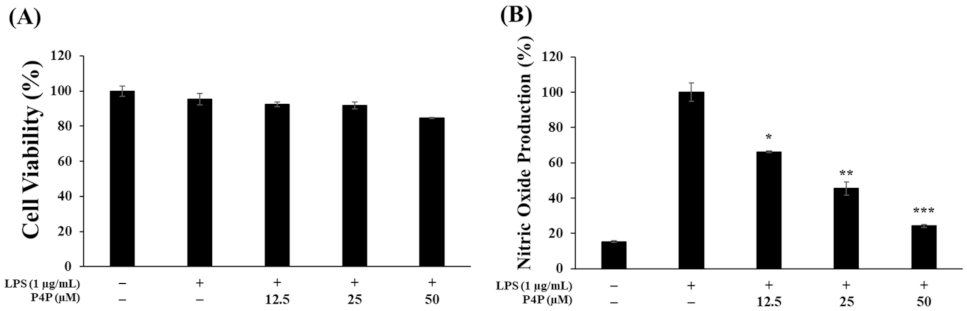
2.3. Effects of P4P on the Viability of RAW 264.7 Cells and NO Production
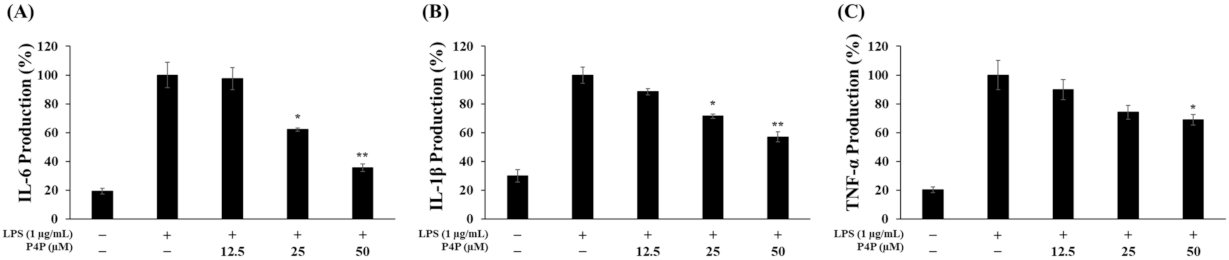
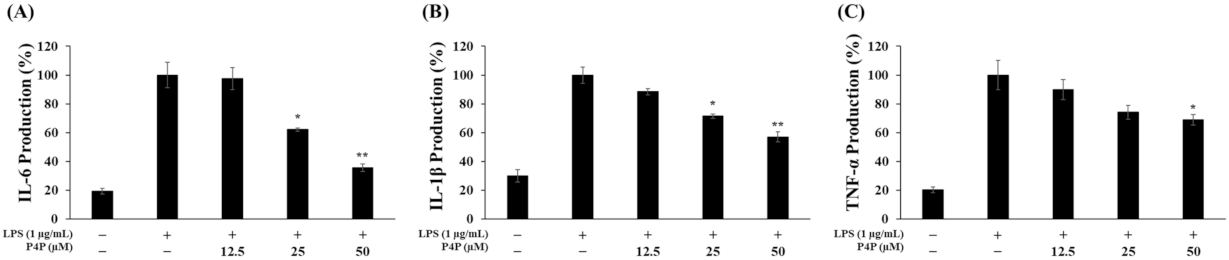
2.4. Suppression of LPS-Induced IL6, IL1β, and TNFα Production
2.5. Suppression of LPS-Induced PGE2 Secretion and Protein Expression of iNOS and COX-2
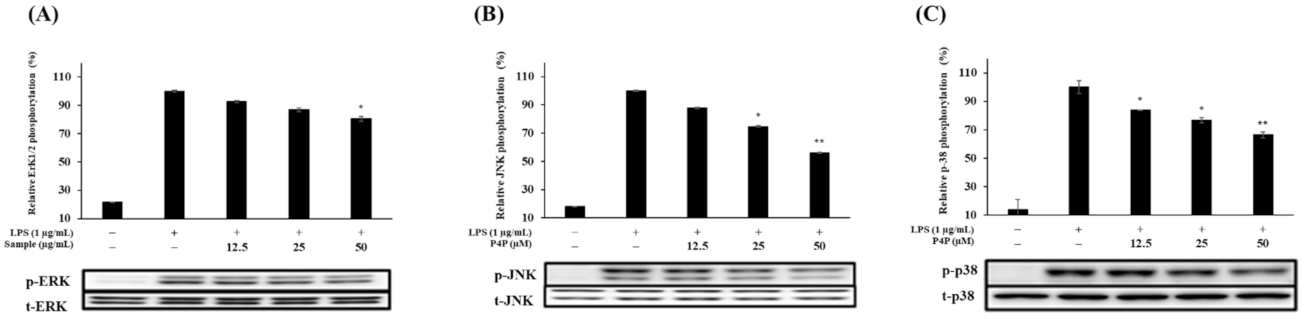
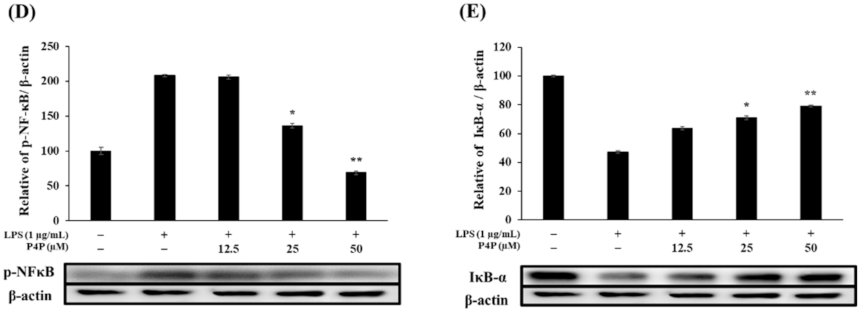
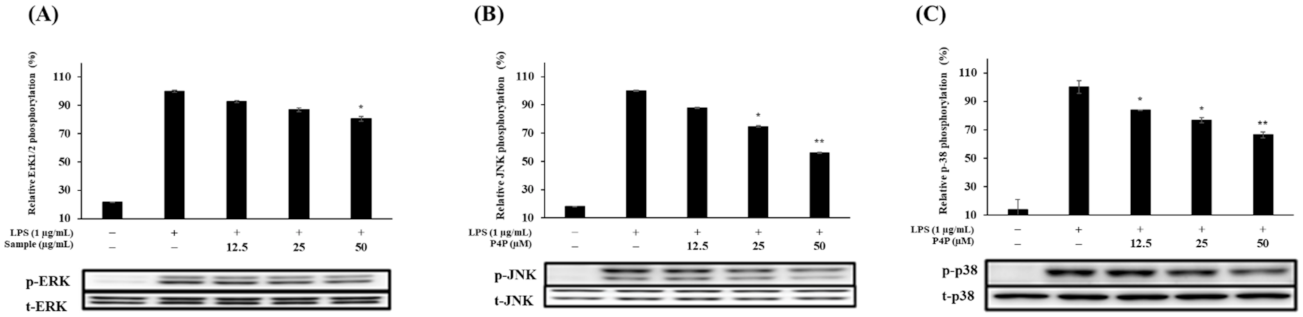
2.6. Expression of Proteins of MAPK and NFκB Pathways
3. Discussion
4. Materials and Methods
4.1. Chemical Preparation
4.2. Biorenovation of PRN
4.3. HPLC Analysis and Purification of PRN Derivatives
4.4. LCMS and NMR Analysis
4.5. Cell Maintenance and Cytotoxicity Assays
4.6. Quantification of NO and PGE2 Secretion
4.7. Determination of IL6, IL1β, and TNFα Production
4.8. Immunoblotting
4.9. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Kim, Y.-J.; Kim, H.-J.; Lee, J.Y.; Kim, D.-H.; Kang, M.S.; Park, W. Anti-Inflammatory Effect of Baicalein on Polyinosinic–Polycytidylic Acid-Induced RAW 264.7 Mouse Macrophages. Viruses 2018, 10, 224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, P.J.; Wynn, T.A. Protective and pathogenic functions of macrophage subsets. Nat. Rev. Immunol. 2011, 11, 723–737. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.W.; Karin, M. A cytokine-mediated link between innate immunity, inflammation, and cancer. J. Clin. Investig. 2007, 117, 1175–1183. [Google Scholar] [CrossRef]
- Ryu, H.W.; Lee, S.U.; Lee, S.; Song, H.-H.; Son, T.H.; Kim, Y.-U.; Yuk, H.J.; Ro, H.; Lee, C.-K.; Hong, S.-T.; et al. 3-Methoxy-catalposide inhibits inflammatory effects in lipopolysaccharide-stimulated RAW264.7 macrophages. Cytokine 2017, 91, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Guha, M.; Mackman, N. LPS induction of gene expression in human monocytes. Cell. Signal. 2001, 13, 85–94. [Google Scholar] [CrossRef]
- Moretta, A.; Marcenaro, E.; Sivori, S.; Della Chiesa, M.; Vitale, M.; Moretta, L. Early liaisons between cells of the innate immune system in inflamed peripheral tissues. Trends Immunol. 2005, 26, 668–675. [Google Scholar] [CrossRef]
- Dai, G.-F.; Zhao, J.; Jiang, Z.-W.; Zhu, L.-P.; Xu, H.-W.; Ma, W.-Y.; Chen, X.-R.; Dong, R.-J.; Li, W.-Y.; Liu, H.-M. Anti-inflammatory effect of novel andrographolide derivatives through inhibition of NO and PGE2 production. Int. Immunopharmacol. 2011, 11, 2144–2149. [Google Scholar] [CrossRef]
- Kaminska, B. MAPK signalling pathways as molecular targets for anti-inflammatory therapy—From molecular mechanisms to therapeutic benefits. Biochim. Biophys. Acta (BBA)-Prot. Proteom. 2005, 1754, 253–262. [Google Scholar] [CrossRef]
- Tsai, T.-C.; Tung, Y.-T.; Kuo, Y.-H.; Liao, J.-W.; Tsai, H.-C.; Chong, K.-Y.; Chen, H.-L.; Chen, C.-M. Anti-inflammatory effects of Antrodia camphorata, a herbal medicine, in a mouse skin ischemia model. J. Ethnopharmacol. 2015, 159, 113–121. [Google Scholar] [CrossRef]
- Wang, W.; Jenkinson, C.; Griscavage, J.; Kern, R.; Arabolos, N.; Byrns, R.; Cederbaum, S.; Ignarro, L. Co-induction of Arginase and Nitric Oxide Synthase in Murine Macrophages Activated by Lipopolysaccharide. Biochem. Biophys. Res. Commun. 1995, 210, 1009–1016. [Google Scholar] [CrossRef]
- Zhu, F.; Du, B.; Xu, B. Anti-inflammatory effects of phytochemicals from fruits, vegetables, and food legumes: A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 1260–1270. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Shibuya, N.; Narukawa, Y.; Oshima, N.; Hada, N.; Kiuchi, F. Synergistic effect of baicalein, wogonin and oroxylin A mixture: multistep inhibition of the NFκB signaling pathway contributes to an anti-inflammatory effect of Scutellaria root flavonoids. J. Nat. Med. 2018, 72, 181–191. [Google Scholar] [CrossRef]
- Pearson, G.; Robinson, F.; Beers Gibson, T.; Xu, B.-E.; Karandikar, M.; Berman, K.; Cobb, M.H. Mitogen-Activated Protein (MAP) Kinase Pathways: Regulation and Physiological Functions*. Endocr. Rev. 2001, 22, 153–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hommes, D.W.; Peppelenbosch, M.P.; van Deventer, S.J. Mitogen activated protein (MAP) kinase signal transduction path-ways and novel anti-inflammatory targets. Gut 2003, 52, 144–151. [Google Scholar] [CrossRef] [Green Version]
- Lieb, K.; Engels, S.; Fiebich, B.L. Inhibition of LPS-induced iNOS and NO synthesis in primary rat microglial cells. Neurochem. Int. 2003, 42, 131–137. [Google Scholar] [CrossRef]
- Makarov, S.S. NF-κB as a therapeutic target in chronic inflammation: recent advances. Mol. Med. Today 2000, 6, 441–448. [Google Scholar] [CrossRef]
- Koo, H.J.; Yoon, W.J.; Sohn, E.H.; Ham, Y.M.; Jang, S.A.; Kwon, J.E.; Jeong, Y.J.; Kwak, J.H.; Sohn, E.; Park, S.Y.; et al. The analgesic and anti-inflammatory effects of Litsea japonica fruit are mediated via suppression of NFκB and JNK/p38 MAPK ac-tivation. Int. Immunopharmacol. 2014, 22, 84–97. [Google Scholar] [CrossRef]
- Kiemer, A.K.; Hartung, T.; Huber, C.; Vollmar, A.M. Phyllanthus amarus has anti-inflammatory potential by inhibition of iNOS, COX-2, and cytokines via the NFκB pathway. J. Hepatol. 2003, 38, 289–297. [Google Scholar] [CrossRef]
- Wang, D.; DuBois, R.N. The role of COX-2 in intestinal inflammation and colorectal cancer. Oncogene 2009, 29, 781–788. [Google Scholar] [CrossRef] [Green Version]
- Park, K.E.; Qin, Y.; Bavry, A. Nonsteroidal anti-inflammatory drugs and their effects in the elderly. Aging Health 2012, 8, 167–177. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-S.; Ahn, C.-B.; Je, J.-Y. Anti-inflammatory action of high molecular weight Mytilus edulis hydrolysates fraction in LPS-induced RAW264.7 macrophage via NF-κB and MAPK pathways. Food Chem. 2016, 202, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Shalini, V.; Pushpan, C.K.; Sindhu, G.; Jayalekshmy, A.; Helen, A. Tricin, flavonoid from Njavara reduces inflammatory responses in hPBMCs by modulating the p38MAPK and PI3K/Akt pathways and prevents inflammation associated endothelial dysfunction in HUVECs. Immunobiology 2016, 221, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.-T.; Wang, M.; Yang, L.; Jiang, J.-G.; Zhao, J.-W.; Zhu, W. Flavonoid glycosides from Rubus chingii Hu fruits display anti-inflammatory activity through suppressing MAPKs activation in macrophages. J. Funct. Foods 2015, 18, 235–243. [Google Scholar] [CrossRef]
- Li, D.; Chen, J.; Ye, J.; Zhai, X.; Song, J.; Jiang, C.; Wang, J.; Zhang, H.; Jia, X.; Zhu, F. Anti-inflammatory effect of the six compounds isolated from Nauclea officinalis Pierrc ex Pitard, and molecular mechanism of strictosamide via suppressing the NFκB and MAPK signaling path-way in LPS-induced RAW 264.7 macrophages. J. Ethnopharmacol. 2017, 196, 66–74. [Google Scholar] [CrossRef]
- Pateras, I.; Giaginis, C.; Tsigris, C.; Patsouris, E.; Theocharis, S.; Madonna, R.; De Caterina, R. NF-κB signaling at the crossroads of inflammation and atherogenesis: searching for new therapeutic links relevance of new drug discovery to reduce NF-κB activation in cardiovascular disease. Expert Opin. Ther. Targets 2014, 18, 1089–1101. [Google Scholar] [CrossRef]
- Maulik, N.; Sato, M.; Price, B.D.; Das, D.K. An essential role of NF-κB in tyrosine kinase signaling of p38 MAP kinase regulation of myocardial adaptation to ischemia. FEBS Lett. 1998, 429, 365–369. [Google Scholar] [CrossRef] [Green Version]
- Bonizzi, G.; Karin, M. The two NF-κB activation pathways and their role in innate and adaptive immunity. Trends Immunol. 2004, 25, 280–288. [Google Scholar] [CrossRef]
- Castañeda-Ovando, A.; de Lourdes Pacheco-Hernández, M.; Páez-Hernández, M.E.; Rodríguez, J.A.; Galán-Vidal, C.A. Chemical studies of anthocyanins: a review. Food Chem. 2009, 113, 859–871. [Google Scholar] [CrossRef]
- Burak, M.; Imen, Y. Antioxidant properties of flavonoids. Med. J. Indones. 1999, 19, 296–304. [Google Scholar]
- Lee, Y.K.; Yuk, D.Y.; Lee, J.W.; Lee, S.Y.; Ha, T.Y.; Oh, K.W.; Yun, Y.P.; Hong, J.T. Epigallocatechin-3-gallate prevents lipopolysaccharide-induced elevation of β-amyloid genera-tion and memory deficiency. Brain Res. 2009, 1250, 164–174. [Google Scholar] [CrossRef]
- Walker, E.H.; Pacold, M.E.; Perisic, O.; Stephens, L.; Hawkins, P.; Wymann, M.; Williams, R.L. Structural Determinants of Phosphoinositide 3-Kinase Inhibition by Wortmannin, LY294002, Quercetin, Myricetin, and Staurosporine. Mol. Cell 2000, 6, 909–919. [Google Scholar] [CrossRef]
- Metodiewa, D.; Kochman, A.; Karolczak, S. Evidence for antiradical and antioxidant properties of four biologically active N, N, diethylaminoethyl ethers of flavanone oximes: a comparison with natural polyphenolic flavonoid (rutin) action. Biochem. Mol. Biol. Int. 1997, 41, 1067–1075. [Google Scholar] [PubMed]
- Hayashi, T.; Sawa, K.; Kawasaki, M.; Arisawa, M.; Shimizu, M.; Morita, N. Inhibition of Cow’s Milk Xanthine Oxidase by Flavonoids. J. Nat. Prod. 1988, 51, 345–348. [Google Scholar] [CrossRef]
- Silver, L.L.; Bostian, K. Screening of natural products for antimicrobial agents. Eur. J. Clin. Microbiol. Infect. Dis. 1990, 9, 455–461. [Google Scholar] [CrossRef]
- Grange, J.M.; Davey, R.W. Antibacterial properties of propolis (bee glue). J. R. Soc. Med. 1990, 83, 159–160. [Google Scholar] [CrossRef] [PubMed]
- Shriner, R.L.; Hull, C.J. Isoflavones. III. The Structure of Prunetin and a New Synthesis of Genistein1. J. Org. Chem. 1945, 10, 288–291. [Google Scholar] [CrossRef]
- Middleton, E., Jr.; Kandaswami, C.; Theoharides, T.C. The effects of plant flavonoids on mammalian cells: implications for inflammation, heart disease, and cancer. Pharmacol. Rev. 2000, 52, 673–751. [Google Scholar]
- Barnes, S. Soy isoflavones–phytoestrogens and what else? J. Nutr. 2004, 134, 1225S–1228S. [Google Scholar] [CrossRef] [Green Version]
- Nam, D.C.; Kim, B.K.; Lee, H.J.; Shin, H.D.; Lee, C.J.; Hwang, S.C. Effects of prunetin on the proteolytic activity, secretion and gene expression of MMP-3 in vitro and pro- duction of MMP-3 in vivo. Korean J. Physiol. Pharmacol. 2016, 20, 221–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, T.-G.; Yang, G.; Lee, H.-M.; Kim, M.-D.; Choi, H.-Y.; Park, K.-S.; Lee, S.-D.; Kook, Y.-B.; An, H.-J. Molecular mechanisms underlying the anti-obesity potential of prunetin, an O-methylated isoflavone. Biochem. Pharmacol. 2013, 85, 1525–1533. [Google Scholar] [CrossRef] [PubMed]
- Koirala, M.; Lee, Y.K.; Kim, M.S.; Chung, Y.C.; Park, J.S.; Kim, S.Y. Biotransformation of Naringenin by Bacillus amylolique-faciens Into Three Naringenin Derivatives. Nat. Prod. Commun. 2019, 14, 1934578–19851971. [Google Scholar]
- Kim, K.-M.; Park, J.-S.; Choi, H.; Kim, M.-S.; Seo, J.-H.; Pandey, R.P.; Kim, J.W.; Hyun, C.-G.; Kim, S.-Y. Biosynthesis of novel daidzein derivatives using Bacillus amyloliquefaciens whole cells. Biocatal. Biotransform. 2018, 36, 469–475. [Google Scholar] [CrossRef]
- Choi, H.; Park, J.-S.; Kim, K.-M.; Kim, M.; Ko, K.-W.; Hyun, C.-G.; Ahn, J.W.; Seo, J.-H.; Kim, S.-Y. Enhancing the antimicrobial effect of genistein by biotransformation in microbial system. J. Ind. Eng. Chem. 2018, 63, 255–261. [Google Scholar] [CrossRef]
- Kim, M.S.; Park, J.S.; Chung, Y.C.; Jang, S.C.; Hyun, C.G.; Kim, S.Y. Anti-Inflammatory Effects of Formononetin 7-O-phosphate, a Novel Biorenovation Product, on LPS-Stimulated RAW 264.7 Macrophage Cells. Molecules 2019, 24, 3910. [Google Scholar] [CrossRef] [Green Version]
- Dong, N.; Li, X.; Xue, C.; Zhang, L.; Wang, C.; Xu, X.; Shan, A.; Dong, N.; Li, X.; Xue, C.; et al. Astragalus polysaccharides alleviates LPS-induced inflammation via the NFκB/MAPK signaling pathway. J. Cell Physiol. 2020, 235, 5525–5540. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.K.; Choi, E.-J. Compromised MAPK signaling in human diseases: an update. Arch. Toxicol. 2015, 89, 867–882. [Google Scholar] [CrossRef]
- Dhillon, A.S.; Hagan, S.; Rath, O.; Kolch, W. MAP kinase signalling pathways in cancer. Oncogene 2007, 26, 3279–3290. [Google Scholar] [CrossRef] [Green Version]
- Fuentes, E.; Rojas, A.; Palomo, I. NF-κB signaling pathway as target for antiplatelet activity. Blood Rev. 2016, 30, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Anti-inflammatory Agents: Present and Future. Cell 2010, 140, 935–950. [Google Scholar] [CrossRef] [Green Version]
- Mazumder MA, R.; Hongsprabhas, P. Genistein as antioxidant and antibrowning agents in in vivo and in vitro: A review. Biomed. Pharmacother. 2016, 82, 379–392. [Google Scholar] [CrossRef]
- Li, Q.-S.; Li, C.-Y.; Li, Z.-L.; Zhu, H.-L. Genistein and its Synthetic Analogs as Anticancer Agents. Anti-Cancer Agents Med. Chem. 2012, 12, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Zhou, J.; Zhao, H.; Fan, M.; Gao, W. The anti-inflammatory effects of formononetin and ononin on lipopolysaccharide-induced zebrafish models based on lipidomics and targeted transcriptomics. Metabolomics 2019, 15, 153. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Van Valkenburgh, J.; Hong, X.; Conti, P.S.; Zhang, X.; Chen, K. Small molecules as theranostic agents in cancer immunology. Theranostics 2019, 9, 7849–7871. [Google Scholar] [CrossRef]
- Tang, L.; Singh, R.; Liu, Z.; Hu, M. Structure and Concentration Changes Affect Characterization of UGT Isoform-Specific Metabolism of Isoflavones. Mol. Pharm. 2009, 6, 1466–1482. [Google Scholar] [CrossRef]
- Zhao, J.; Yang, J.; Xie, Y. Improvement strategies for the oral bioavailability of poorly water-soluble flavonoids: An overview. Int. J. Pharm. 2019, 570, 118642. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Li, D.D.; Zeng, F.; Zhu, Z.H.; Song, P.; Zhao, M.; Duan, J.A. Efficient biosynthesis, analysis, solubility and an-ti-bacterial activities of succinylglycosylated naringenin. Nat. Prod. Res. 2019, 33, 1756–1760. [Google Scholar] [CrossRef]
- Byun, S.H.; Yang, C.H.; Kim, S.C. Inhibitory effect of Sctophulariae Radix extract on TNFα, IL1β, IL6 and Nitric Oxide pro-duction in lipoposaccharideactivated Raw 264.7 cells. Korea J. Herbol. 2005, 20, 7–16. [Google Scholar]
- Hyun, M.S.; Woo, W.H.; Hur, J.M.; Kim, D.H.; Moon, Y.J. The role of ROS and p38 MAP kinase in berberineinduced apoptosis on human hepatoma HepG2 cells. J. Korean Soc. Appl. Biol. Chem. 2008, 51, 129–135. [Google Scholar]
- Ahn, J.Y.; Lee, E.R.; Kim, J.Y.; Cho, S.G. Protein phosphorylation as a regulatory mechanism of various cellular function. Cancer Prev. Res. 2006, 11, 1–8. [Google Scholar]
- Chun, J.H.; Kang, S.; Varani, J.; Lin, J.; Fisher, G.J.; Voorhees, J.J. Decreased extracellular signal regulated kinase and increased stress activated MAP kinase activities in aged human skin in vitro. J. Investig. Dermatol. 2000, 15, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Liden, J.; Rafter, I.; Truss, M.; Gustafsson, J.A.; Okret, S. Glucocorticoid effects on NF-κB binding in the transcription of the ICAM-1 gene. Biochem. Biophys. Res. Commun. 2000, 273, 1008–1014. [Google Scholar] [CrossRef]
- Pruett, S.B.; Fan, R.; Zheng, Q. Characterization of glucocorticoid receptor translocation, cytoplasmic IkB, nuclear NF-κB, and activation of NF-κB in T lymphocytes exposed to stressinducible concentrations of corticosterone in vivo. Int. Immunopharmacol. 2003, 3, 1–16. [Google Scholar] [CrossRef]
- Kumar, P.; Nagarajan, A.; Uchil, P. Analysis of Cell Viability by the MTT Assay. Cold Spring Harb. Protoc. 2018, 2018, pdb-prot095505. [Google Scholar] [CrossRef] [PubMed]
- Kamada, T.; Kang, M.-C.; Phan, C.-S.; Zanil, I.I.; Jeon, Y.-J.; Vairappan, C.S. Bioactive Cembranoids from the Soft Coral Genus Sinularia sp. in Borneo. Mar. Drugs 2018, 16, 99. [Google Scholar] [CrossRef] [Green Version]
- Rho, H.S.; Ghimeray, A.K.; Yoo, D.S.; Ahn, S.M.; Kwon, S.S.; Lee, K.H.; Cho, D.H.; Cho, J.Y. Kaempferol and Kaempferol Rhamnosides with Depigmenting and Anti-Inflammatory Properties. Molecules 2011, 16, 3338–3344. [Google Scholar] [CrossRef] [Green Version]
- Kocić, J.; Santibañez, J.F.; Krstić, A.; Mojsilović, S.; Ilić, V.; Bugarski, D. Interleukin-17 modulates myoblast cell migration by inhibiting urokinase type plasminogen activator expression through p38 mitogen-activated protein kinase. Int. J. Biochem. Cell Biol. 2013, 45, 464–475. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, T.-J.; Hong, H.; Kim, M.-S.; Park, J.-S.; Chi, W.-J.; Kim, S.-Y. Prunetin 4′-O-Phosphate, a Novel Compound, in RAW 264.7 Macrophages Exerts Anti-Inflammatory Activity via Suppression of MAP Kinases and the NFκB Pathway. Molecules 2021, 26, 6841. https://doi.org/10.3390/molecules26226841
Park T-J, Hong H, Kim M-S, Park J-S, Chi W-J, Kim S-Y. Prunetin 4′-O-Phosphate, a Novel Compound, in RAW 264.7 Macrophages Exerts Anti-Inflammatory Activity via Suppression of MAP Kinases and the NFκB Pathway. Molecules. 2021; 26(22):6841. https://doi.org/10.3390/molecules26226841
Chicago/Turabian StylePark, Tae-Jin, Hyehyun Hong, Min-Seon Kim, Jin-Soo Park, Won-Jae Chi, and Seung-Young Kim. 2021. "Prunetin 4′-O-Phosphate, a Novel Compound, in RAW 264.7 Macrophages Exerts Anti-Inflammatory Activity via Suppression of MAP Kinases and the NFκB Pathway" Molecules 26, no. 22: 6841. https://doi.org/10.3390/molecules26226841
APA StylePark, T.-J., Hong, H., Kim, M.-S., Park, J.-S., Chi, W.-J., & Kim, S.-Y. (2021). Prunetin 4′-O-Phosphate, a Novel Compound, in RAW 264.7 Macrophages Exerts Anti-Inflammatory Activity via Suppression of MAP Kinases and the NFκB Pathway. Molecules, 26(22), 6841. https://doi.org/10.3390/molecules26226841