
New Kendomycin Derivative Isolated from Streptomyces sp. Cl 58-27

{kind=link}
{kind=link}
Abstract
:1. Introduction
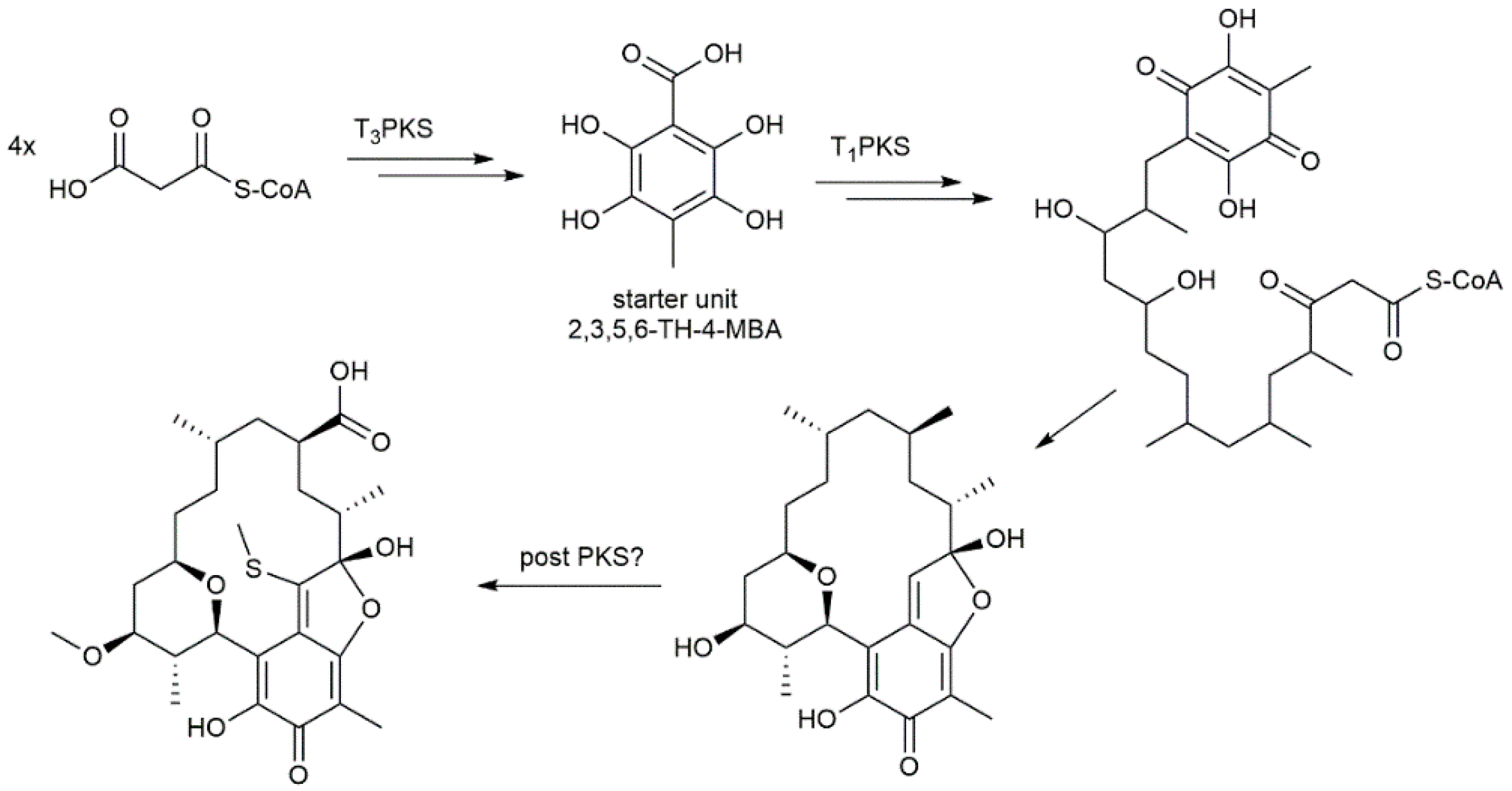
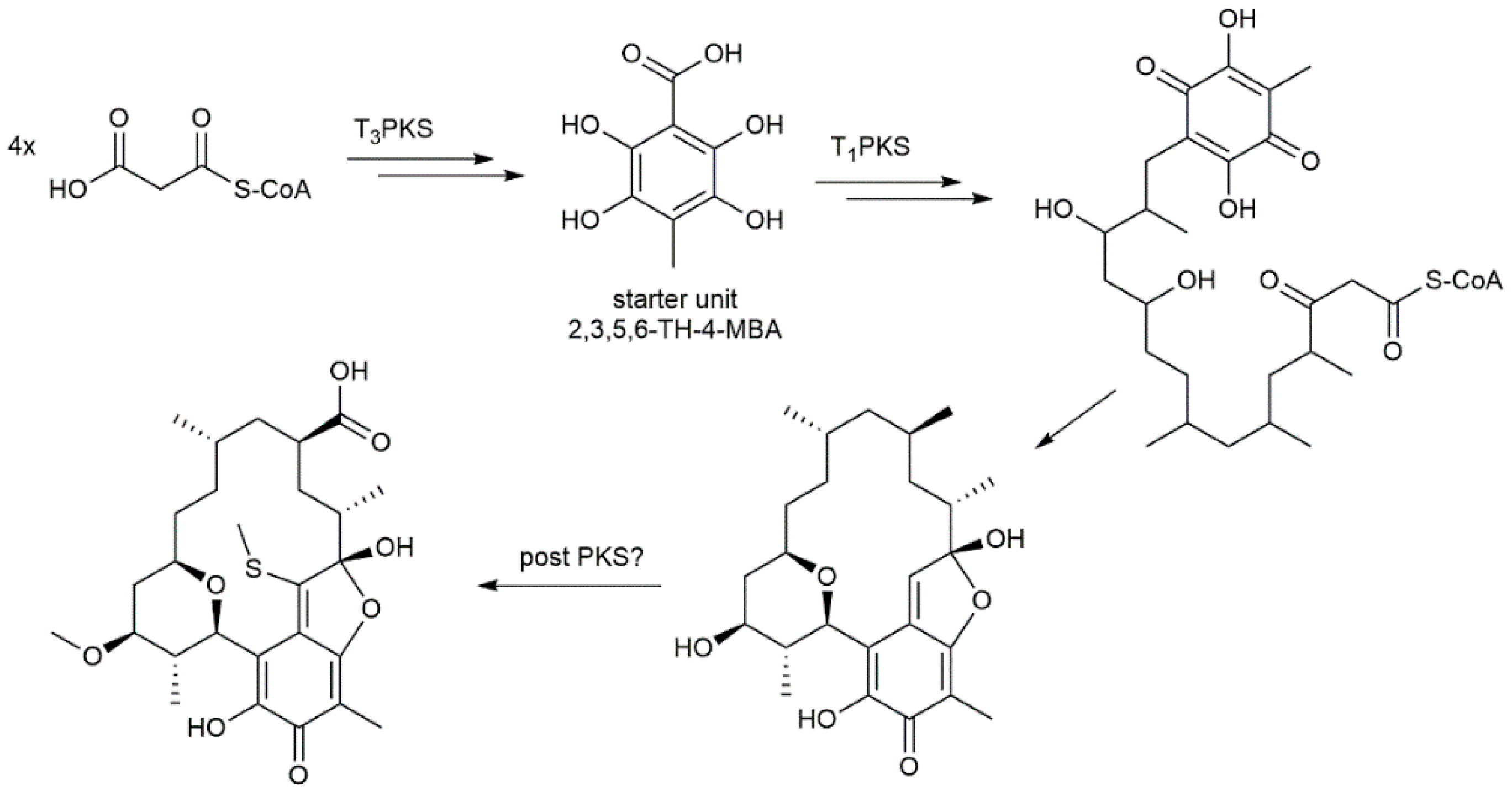
2. Results and Discussion
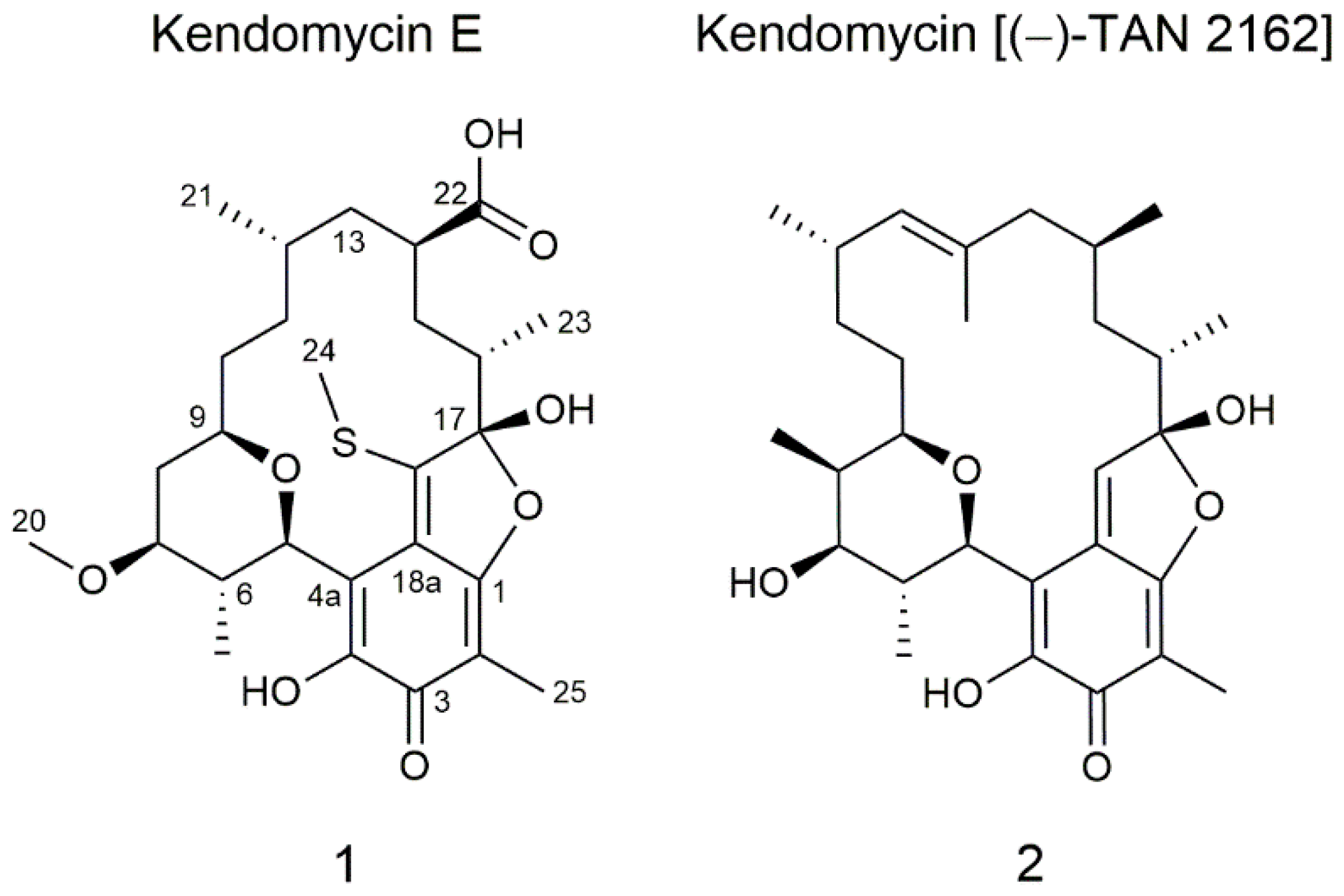
2.1. Isolation and Structure Elucidation of Kendomycin E
2.2. Genome Sequencing of Streptomyces sp. Cl 58-27 and Identification of Kendomycin E Gene Cluster
3. Materials and Methods
3.1. Cultivation, Metabolite Extraction and Dereplication
3.2. Isolation, Purification and Structure Elucidation
3.3. Antimicrobial Susceptibility Test
3.4. Genome Sequencing and Bioinformatics
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Chopra, I.; Roberts, M. Tetracycline antibiotics: Mode of action, applications, molecular biology, and epidemiology of bacterial resistance. Microbiol. Mol. Biol. Rev. 2001, 65, 232–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Risdian, C.; Mozef, T.; Wink, J. Biosynthesis of Polyketides in Streptomyces. Microorganisms 2019, 7, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, B. Polyketide biosynthesis beyond the type I, II and III polyketide synthase paradigms. Curr. Opin. Chem. Biol. 2003, 7, 285–295. [Google Scholar] [CrossRef]
- Tacar, O.; Sriamornsak, P.; Dass, C.R. Doxorubicin: An update on anticancer molecular action, toxicity and novel drug delivery systems. J. Pharm. Pharmacol. 2013, 65, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Whicher, J.R.; Hansen, D.A.; Hale, W.A.; Chemler, J.A.; Congdon, G.R.; Narayan, A.R.H.; Håkansson, K.; Sherman, D.H.; Smith, J.L.; et al. Structure of a modular polyketide synthase. Nature 2014, 510, 512–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, Y.; Ogata, H.; Goto, S. Type III Polyketide Synthases: Functional Classification and Phylogenomics. Chembiochem 2017, 18, 50–65. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Deng, Z.; Liu, T. Streptomyces species: Ideal chassis for natural product discovery and overproduction. Metab. Eng. 2018, 50, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Bode, H.B.; Zeeck, A. Structure and biosynthesis of kendomycin, a carbocyclic ansa-compound from Streptomyces. J. Chem. Soc. Perkin Trans. 1 2000, 31, 323–328. [Google Scholar] [CrossRef]
- Bode, H.B.; Zeeck, A. Biosynthesis of kendomycin: Origin of the oxygen atoms and further investigations. J. Chem. Soc. Perkin Trans. 1 2000, 2665–2670. [Google Scholar] [CrossRef]
- Funahashi, Y.; Kawamura, N.; Ishimaru, T. Jap. Pat. 08231552 [A2 960 010]. Chem. Abstr. 1996, 125, 326–518. [Google Scholar]
- Funahashi, Y.; Kawamura, N.; Ishimaru, T. Jap. Pat. 08231551 [A2 960 910]. Chem. Abstr. 1997, 126, 6553. [Google Scholar]
- Su, M.H.; Hosken, M.I.; Hotovec, B.J.; Johnston, T.L. US Pat. 5728727 [A980317]. Chem. Abstr. 1998, 128, 239–489. [Google Scholar]
- Zhang, S.; Xie, Q.; Sun, C.; Tian, X.-P.; Gui, C.; Qin, X.; Zhang, H.; Ju, J. Cytotoxic Kendomycins Containing the Carbacylic Ansa Scaffold from the Marine-Derived Verrucosispora sp. SCSIO 07399. J. Nat. Prod. 2019, 82, 3366–3371. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, S.C.; Bode, H.B.; Kochems, I.; Müller, R. A type I/type III polyketide synthase hybrid biosynthetic pathway for the structurally unique ansa compound kendomycin. Chembiochem 2008, 9, 2711–2721. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; van Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef] [PubMed]
- Buckingham, J. Dictionary of Natural Products on CD-ROM; Version 13:2; Chapman & Hall: London, UK, 2005; ISBN 9780412491504. [Google Scholar]
- Mo, X.; Shi, C.; Gui, C.; Zhang, Y.; Ju, J.; Wang, Q. Identification of nocamycin biosynthetic gene cluster from Saccharothrix syringae NRRL B-16468 and generation of new nocamycin derivatives by manipulating gene cluster. Microb. Cell Fact. 2017, 16, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kautsar, S.A.; Blin, K.; Shaw, S.; Navarro-Muñoz, J.C.; Terlouw, B.R.; van der Hooft, J.J.J.; van Santen, J.A.; Tracanna, V.; Suarez Duran, H.G.; Pascal Andreu, V.; et al. MIBiG 2.0: A repository for biosynthetic gene clusters of known function. Nucleic Acids Res. 2020, 48, D454–D458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989; p. 1659. [Google Scholar]
- Meyer, F.; Goesmann, A.; McHardy, A.C.; Bartels, D.; Bekel, T.; Clausen, J.; Kalinowski, J.; Linke, B.; Rupp, O.; Giegerich, R.; et al. GenDB—An open source genome annotation system for prokaryote genomes. Nucleic Acids Res. 2003, 31, 2187–2195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paulus, C.; Gromyko, O.; Luzhetskyy, A. New Kendomycin Derivative Isolated from Streptomyces sp. Cl 58-27. Molecules 2021, 26, 6834. https://doi.org/10.3390/molecules26226834
Paulus C, Gromyko O, Luzhetskyy A. New Kendomycin Derivative Isolated from Streptomyces sp. Cl 58-27. Molecules. 2021; 26(22):6834. https://doi.org/10.3390/molecules26226834
Chicago/Turabian StylePaulus, Constanze, Oleksandr Gromyko, and Andriy Luzhetskyy. 2021. "New Kendomycin Derivative Isolated from Streptomyces sp. Cl 58-27" Molecules 26, no. 22: 6834. https://doi.org/10.3390/molecules26226834
APA StylePaulus, C., Gromyko, O., & Luzhetskyy, A. (2021). New Kendomycin Derivative Isolated from Streptomyces sp. Cl 58-27. Molecules, 26(22), 6834. https://doi.org/10.3390/molecules26226834