
Anti-Gastritis and Anti-Lung Injury Effects of Pine Tree Ethanol Extract Targeting Both NF-κB and AP-1 Pathways


 ,
, 
Abstract
:1. Introduction
2. Results
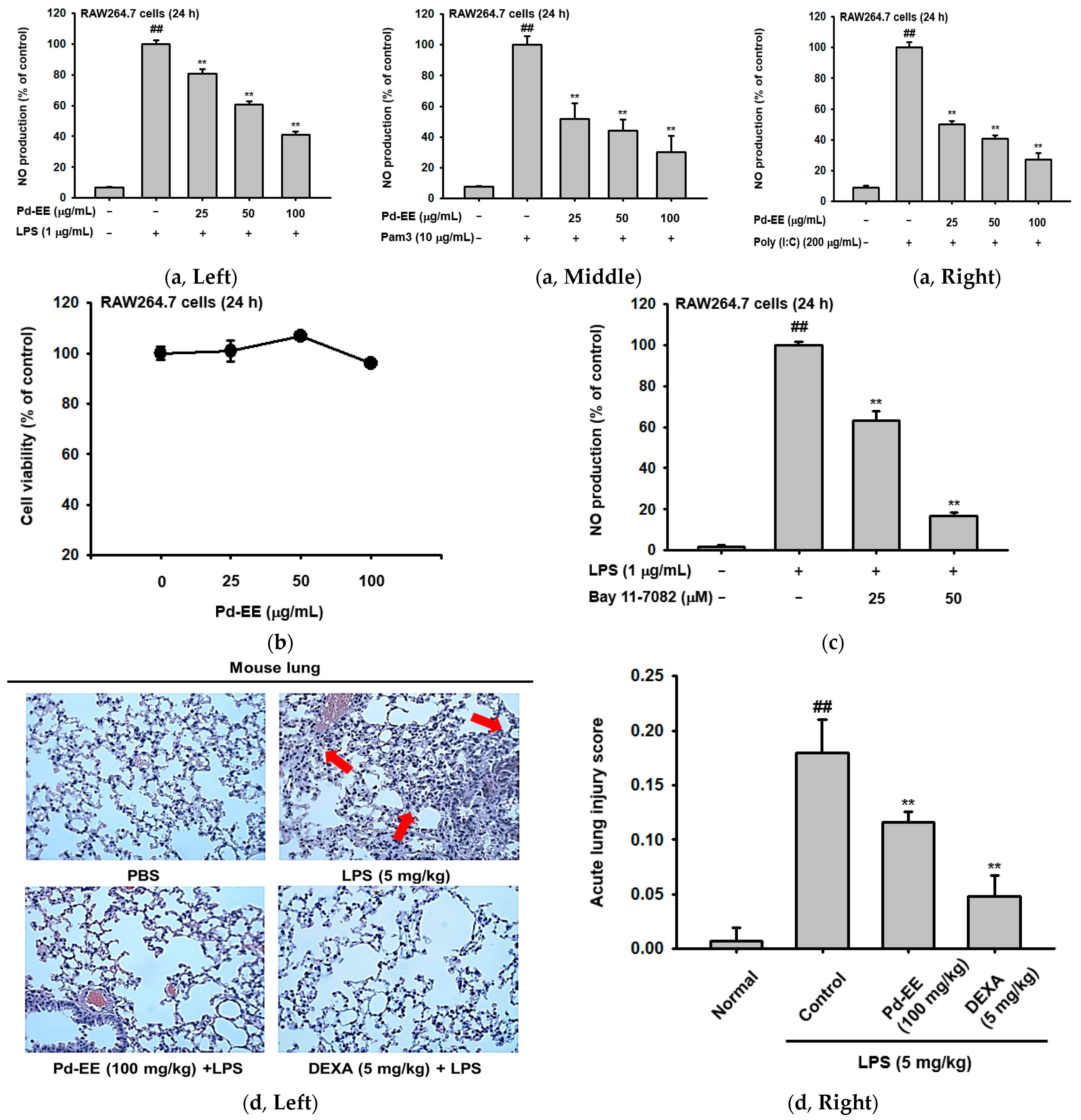
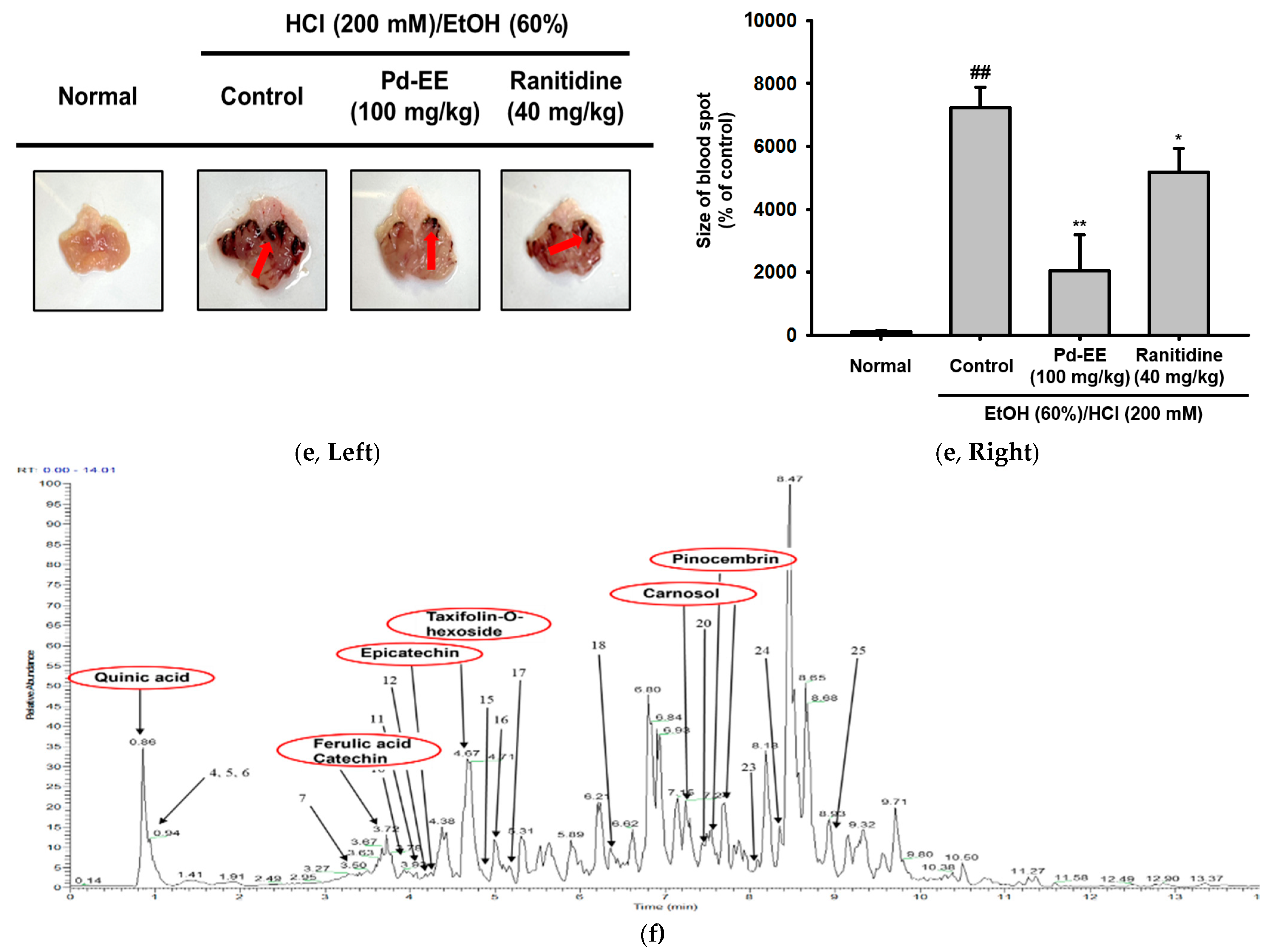
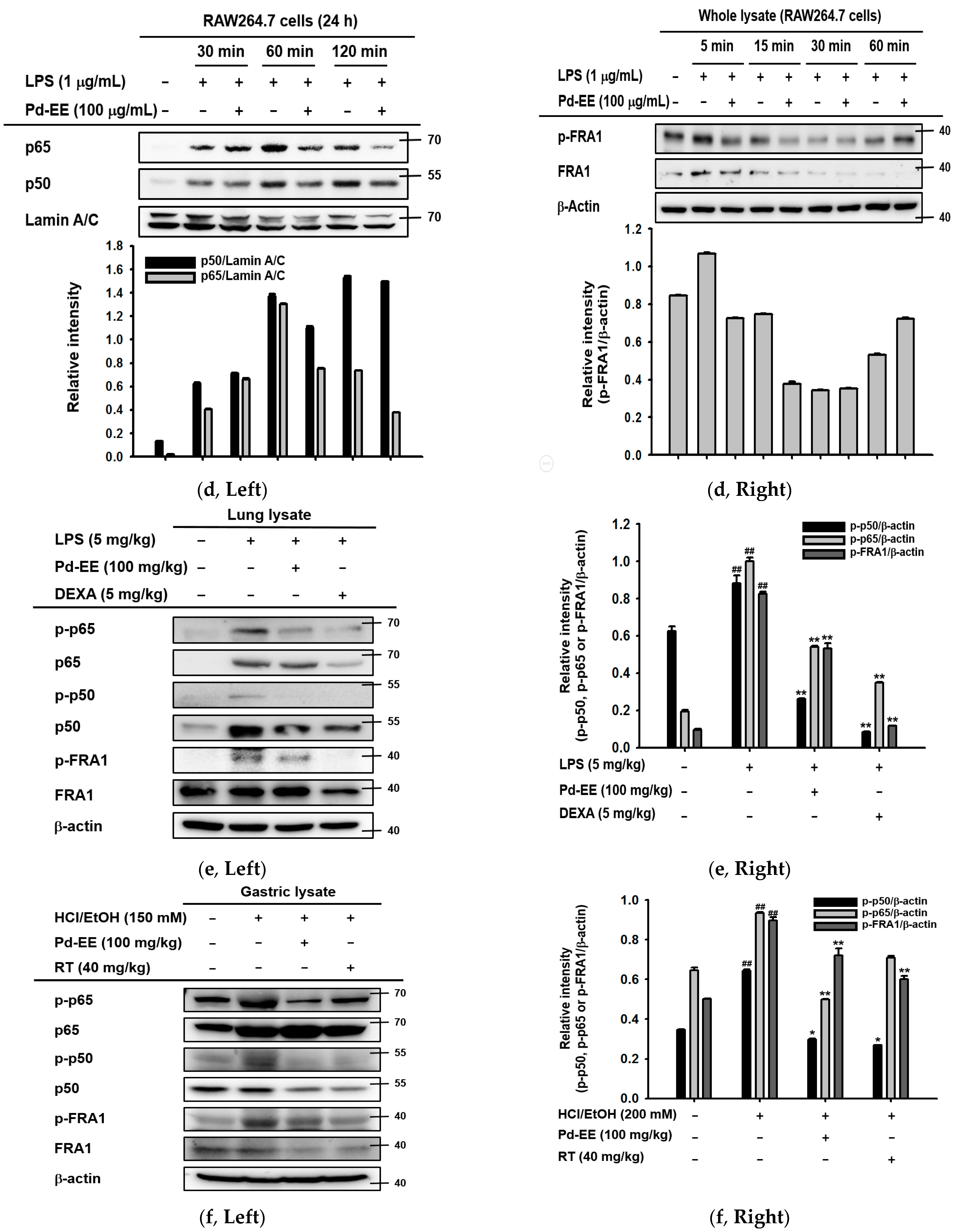
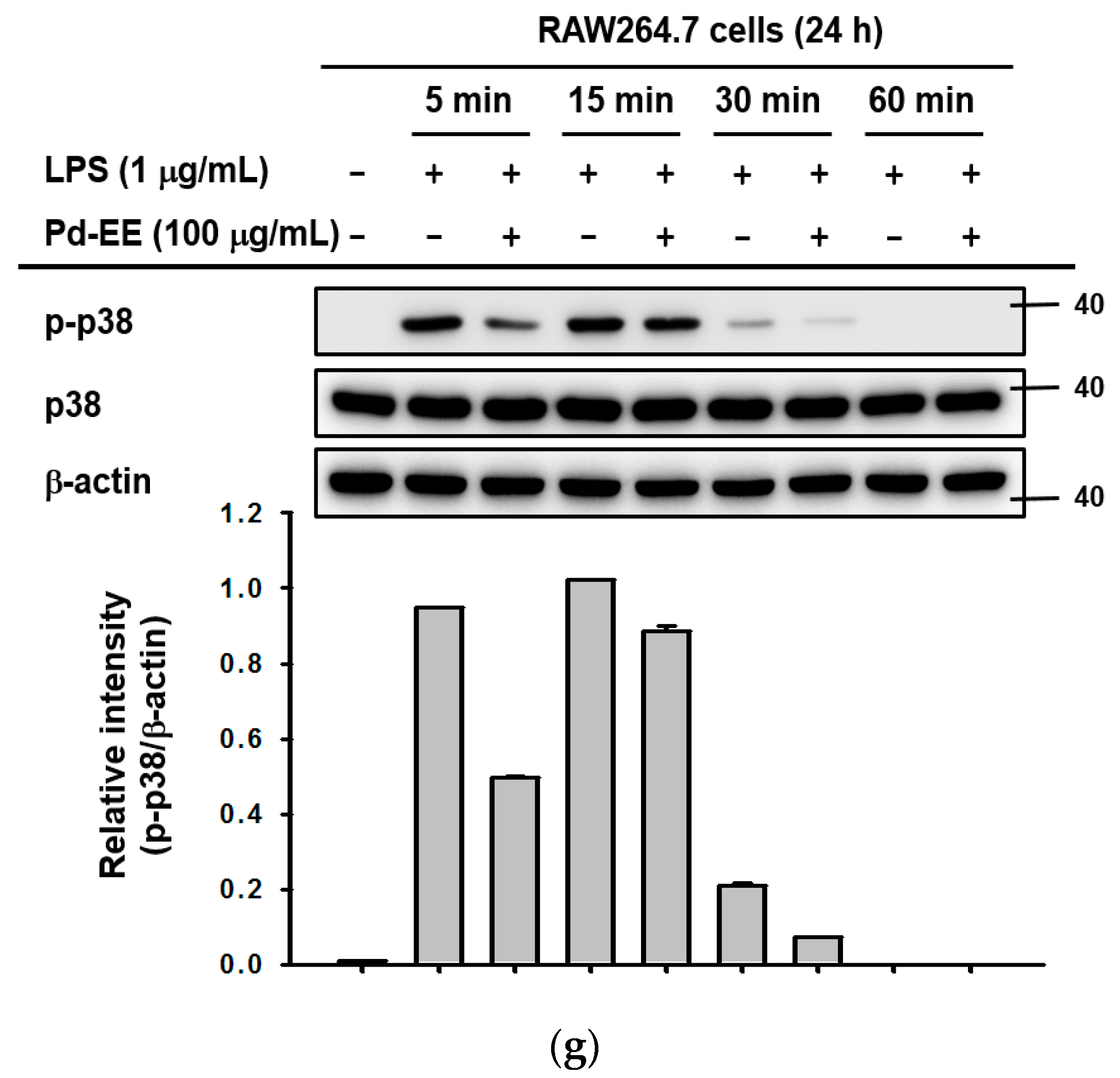
2.1. Pd-EE Alleviates the Inflammatory Response in Both In Vitro and In Vivo Models
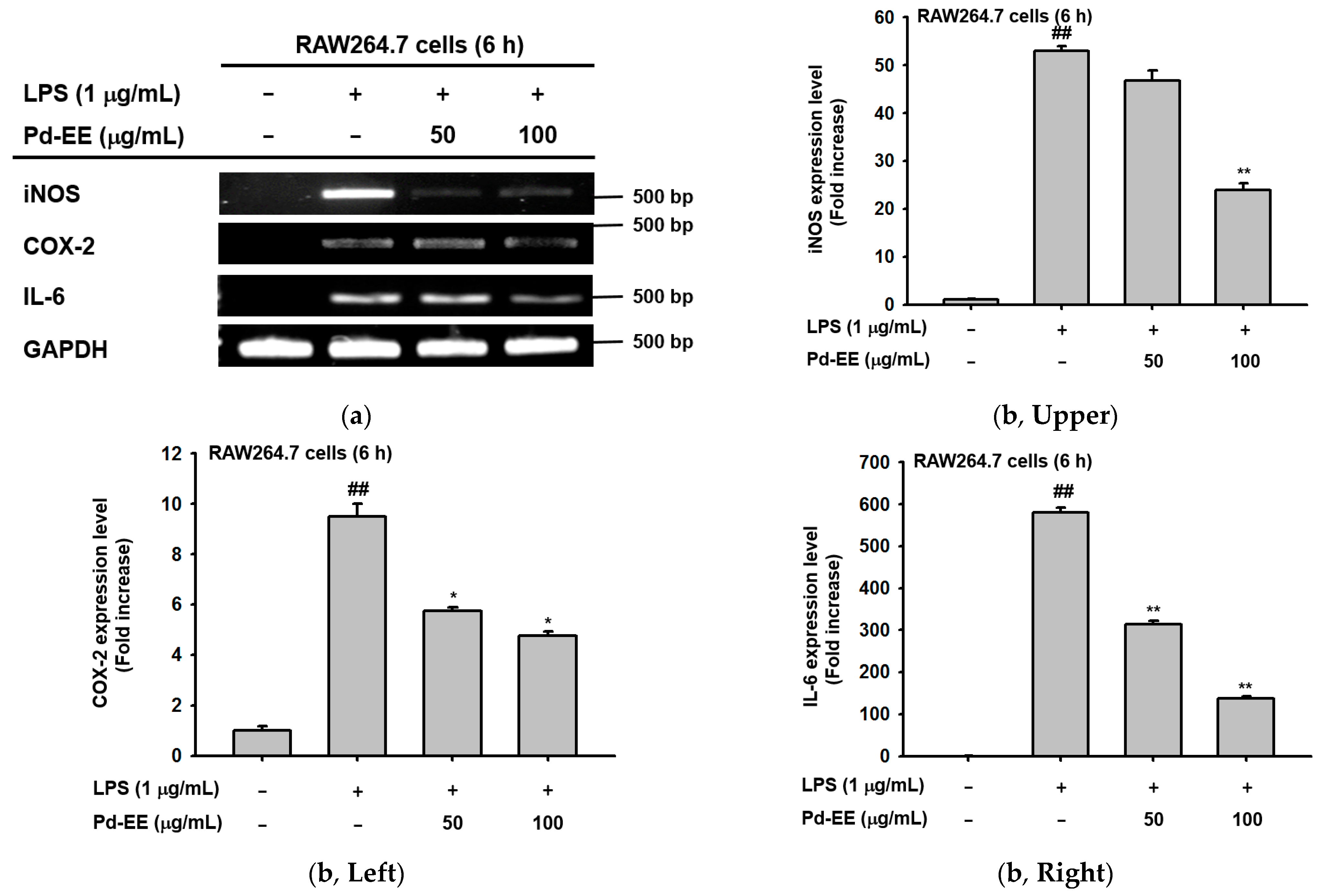
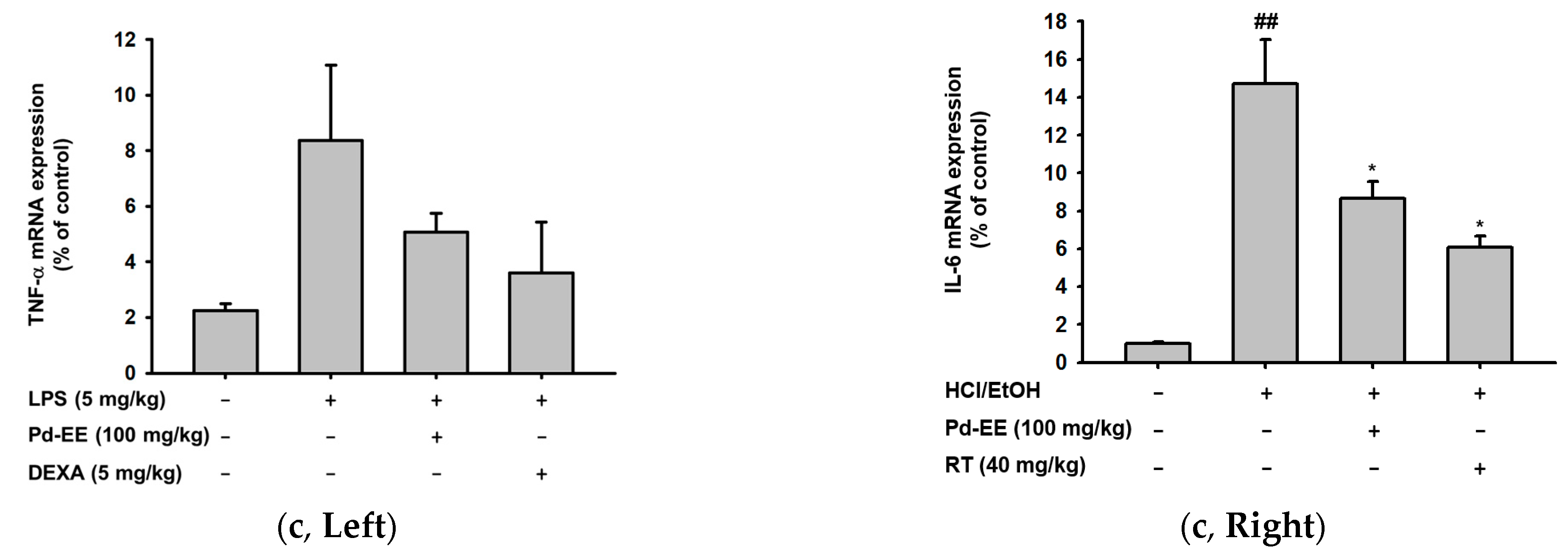
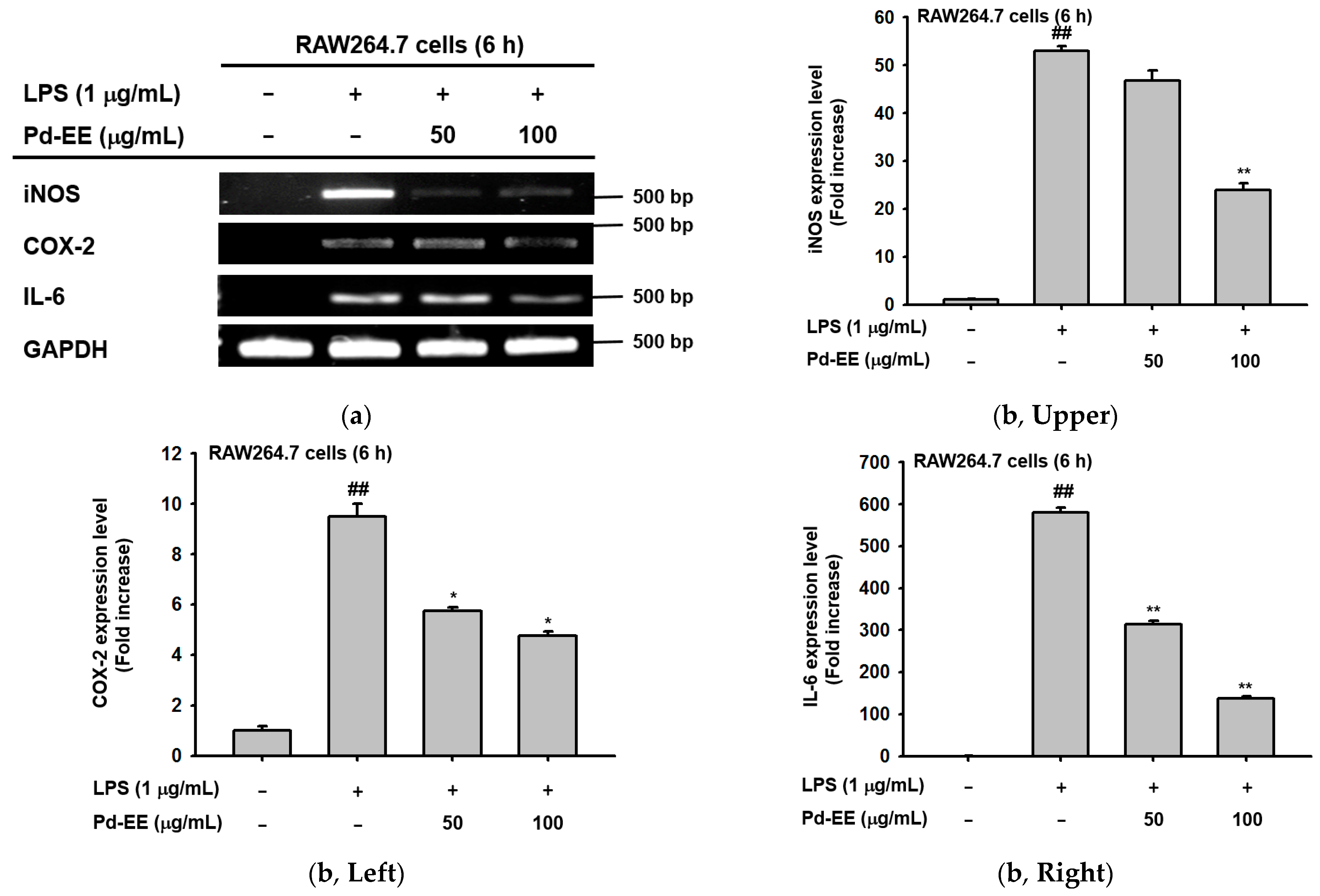
2.2. Anti-Inflammatory Activity of Pd-EE at the Level of mRNA Expression
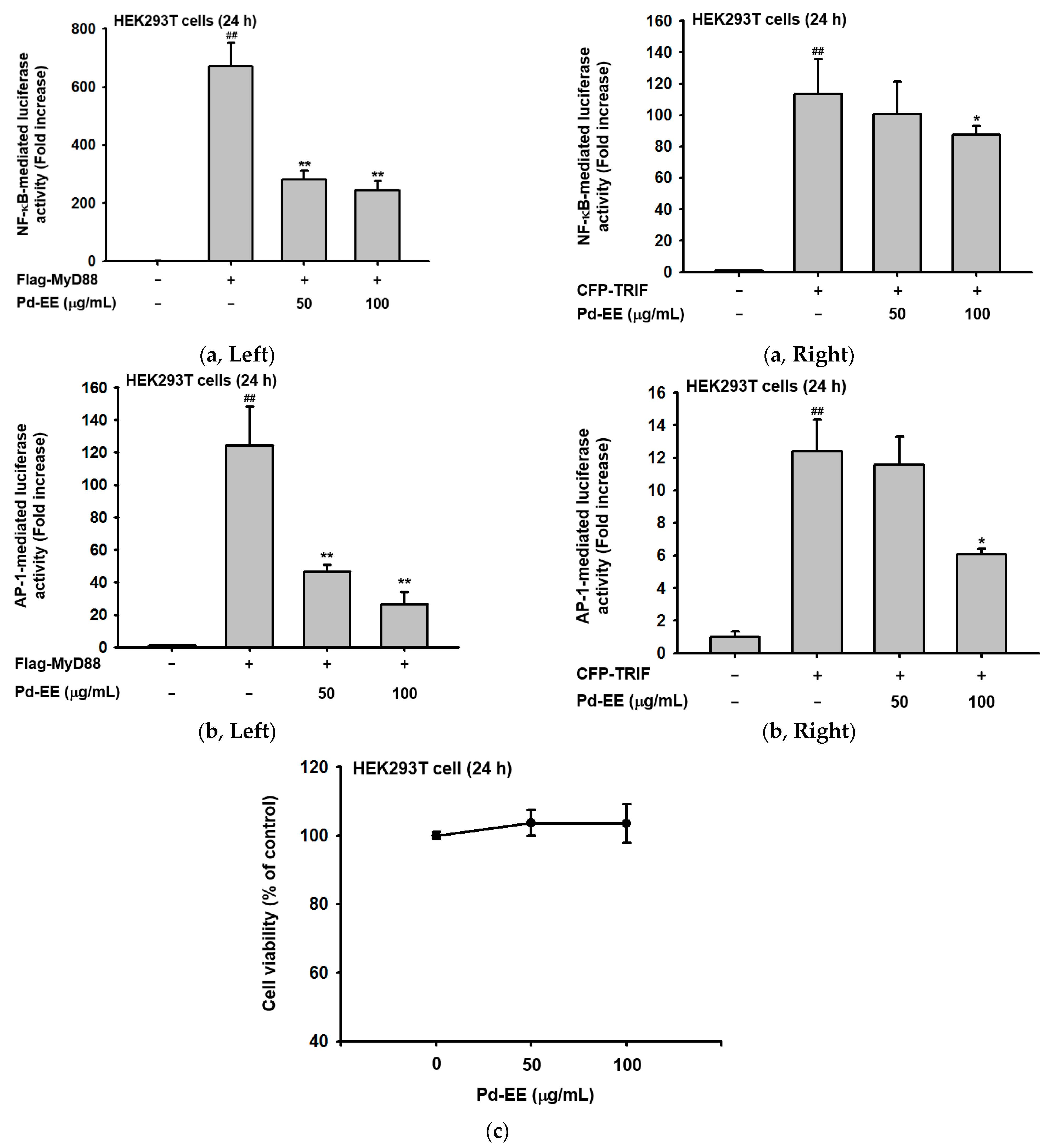
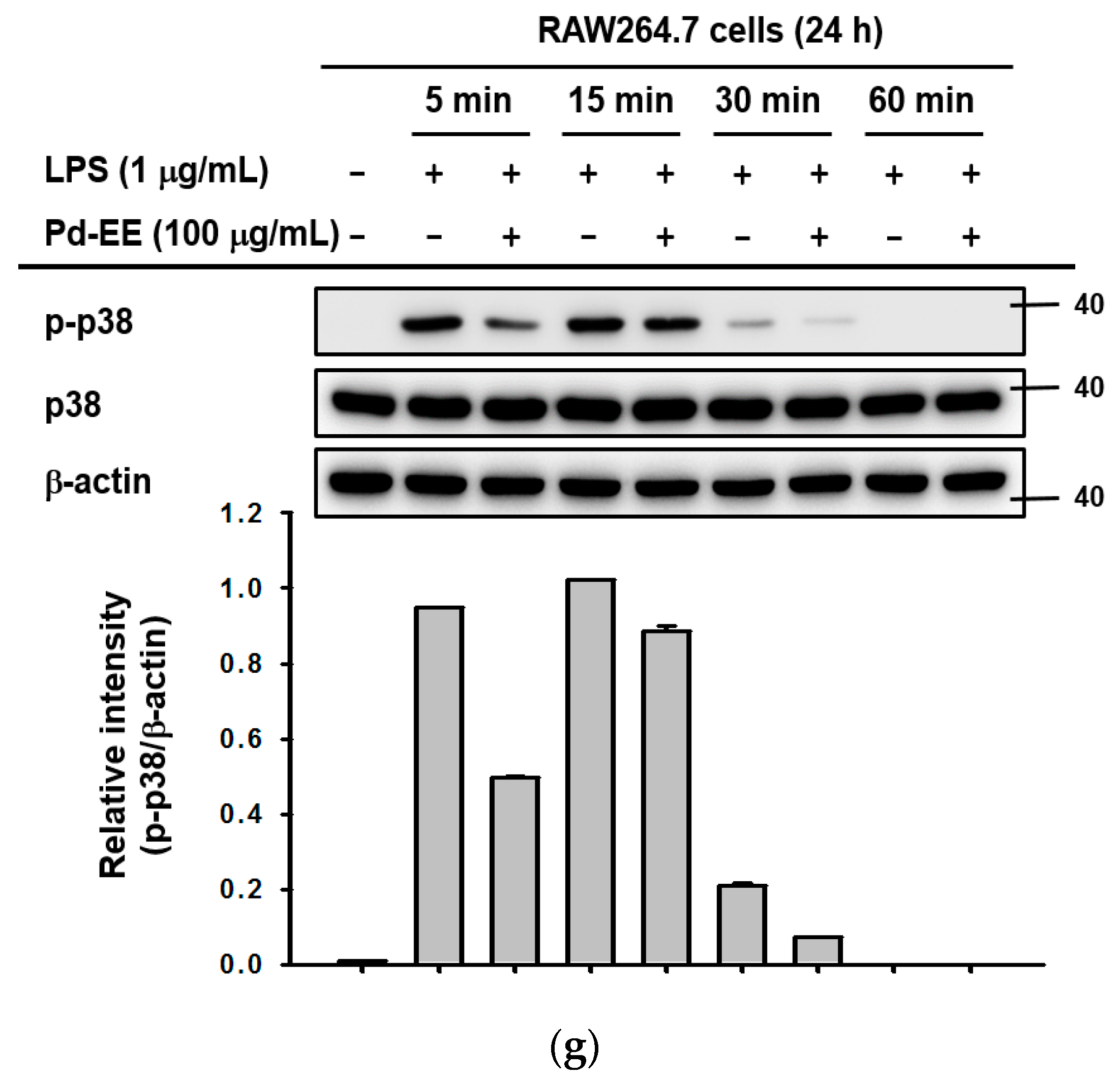
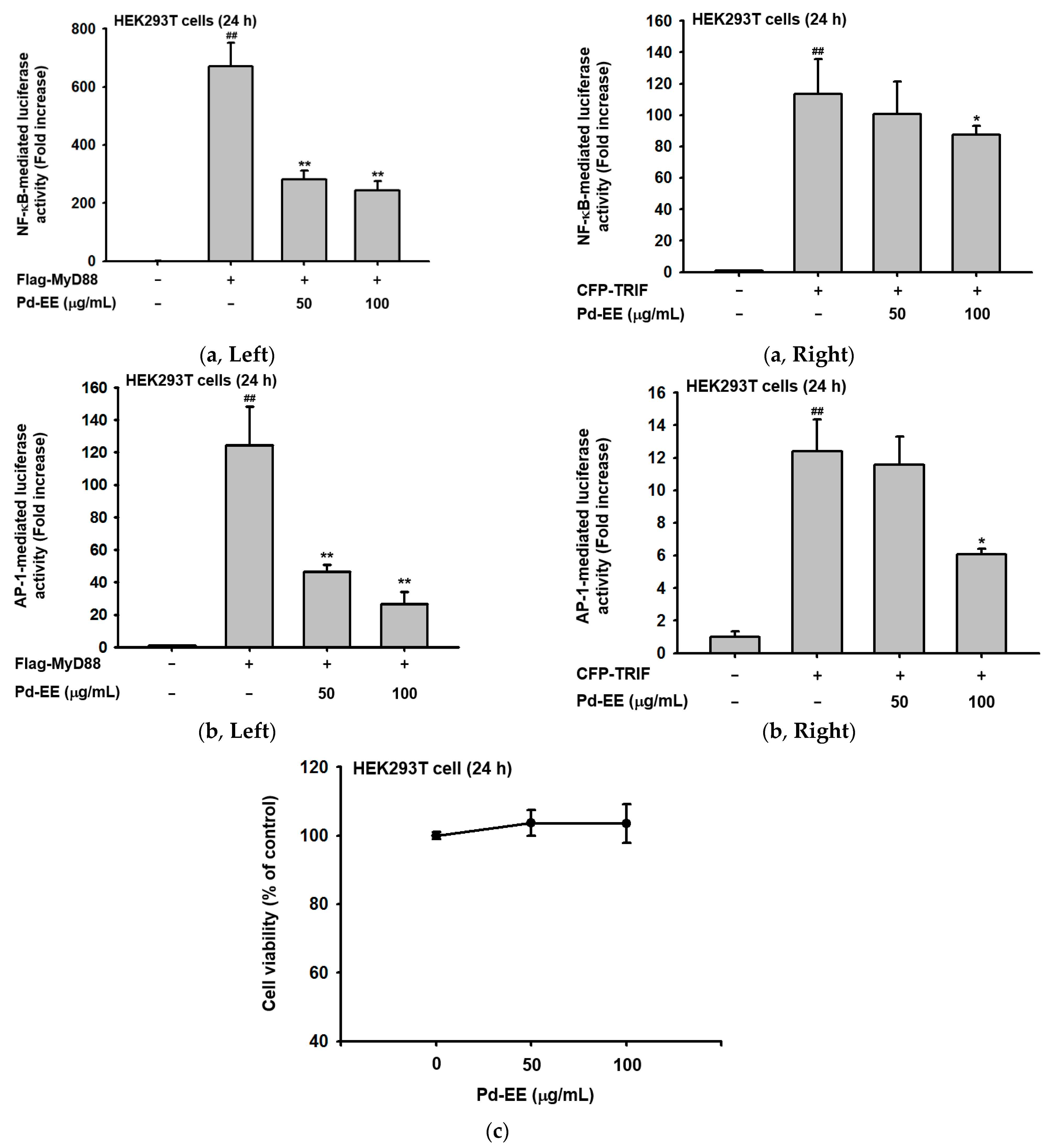
2.3. Pd-EE Affects the Transcriptional Level from the Activity Level of Factors in the Inflammatory Pathway
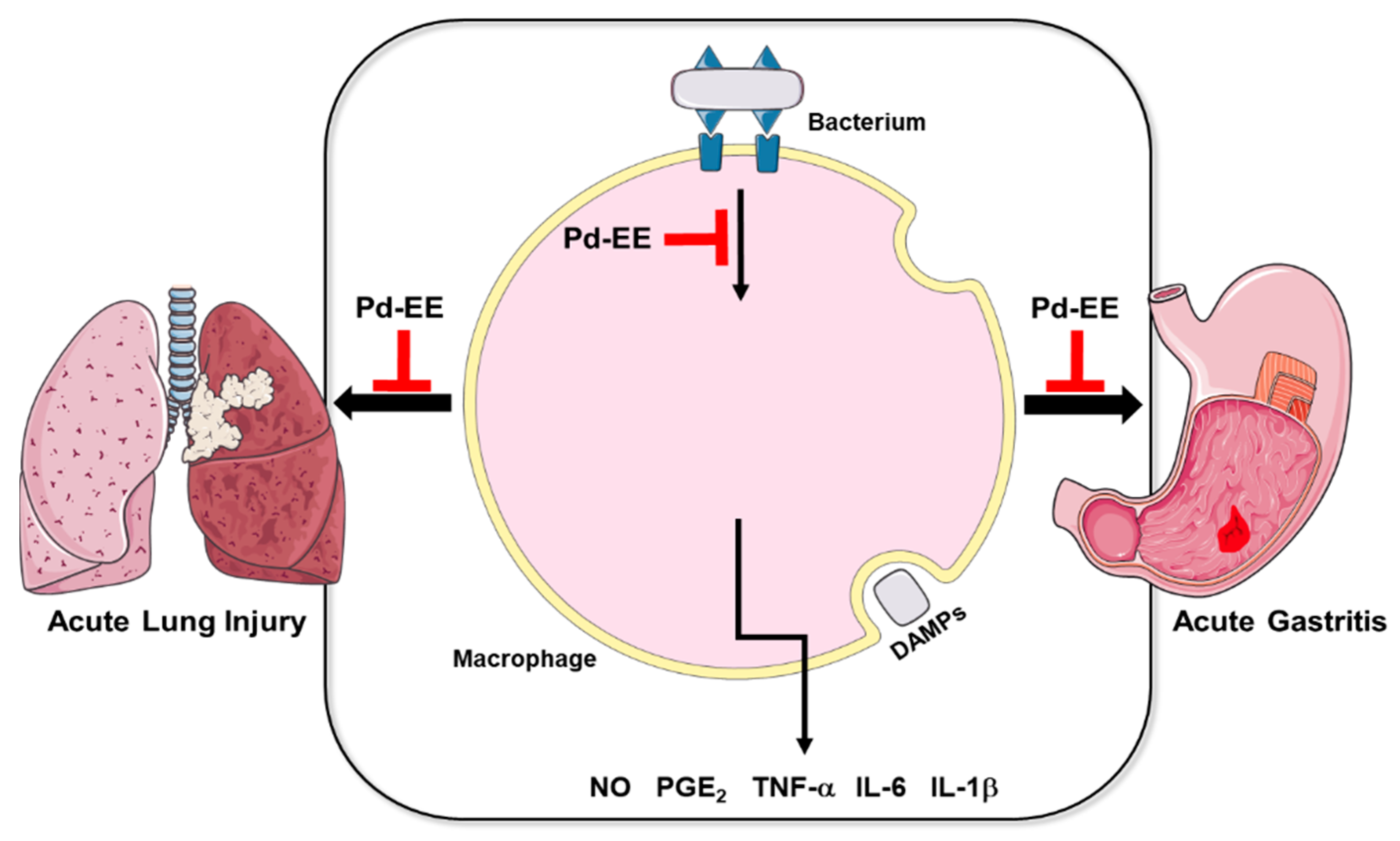
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Cell Culture and Extract Preparation
4.3. Nitric Oxide (NO) Production Assay
4.4. Cell Viability Assay
4.5. mRNA Expression Level Analysis by Semi-Quantitative and Quantitative Reverse Transcription Polymerase Chain Reaction
4.6. Luciferase Reporter Gene Assay
4.7. Whole Lysate Preparation and Western Blotting Assay
4.8. EtOH/HCl-Induced Gastritis Mouse Model
4.9. LPS-Induced Acute Lung Injury Mouse Model
4.10. LC-MS Analysis
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviations
| TAK1 | Transforming growth factor-β-activated kinase 1 |
| NF-κB | Nuclear factor-κB |
| AP-1 | Activator protein-1 |
| MAPKs | Mitogen-activated protein kinases |
| JNK | c-Jun N-terminal kinase |
| PI3K | Phosphoinositide 3-kinase |
| PGE2 | Prostaglandin E2 |
| COX-2 | Cyclooxygenase-2 |
| IL-1β | Interleukin-1β |
| IL-6 | Interleukin 6 |
| TNF-α | Tumor necrosis factor alpha |
| MKK | Mitogen-activated protein kinase |
| iNOS | Inducible nitric oxide synthase |
References
- Choy, E.; Baraliakos, X.; Behrens, F.; D’Angelo, S.; de Vlam, K.; Kirkham, B.W.; Ostergaard, M.; Schett, G.A.; Rissler, M.; Chaouche-Teyara, K.; et al. The need for comparative data in spondyloarthritis. Arthritis. Res. Ther. 2019, 21, A32. [Google Scholar] [CrossRef] [Green Version]
- Bouriche, H.; Kada, S.; Assaf, A.M.; Senator, A.; Gül, F.; Dimertas, I. Phytochemical screening and anti-inflammatory properties of Algerian Hertia cheirifolia methanol extract. Pharm. Biol. 2016, 54, 2584–2590. [Google Scholar] [CrossRef] [Green Version]
- Yun, M.; Yi, Y.S. Regulatory roles of ginseng on inflammatory caspases, executioners of inflammasome activation. J. Ginseng Res. 2020, 44, 373–385. [Google Scholar] [CrossRef]
- Ward, P.A.; Lentsch, A.B. The acute inflammatory response and its regulation. Arch. Surg. 1999, 134, 666–669. [Google Scholar] [CrossRef] [Green Version]
- Amarante-Mendes, G.P.; Adjemian, S.; Branco, L.M.; Zanetti, L.C.; Weinlich, R.; Bortoluci, K.R. Pattern recognition receptors and the host cell death molecular machinery. Front. Immunol. 2018, 9, 2379. [Google Scholar] [CrossRef] [Green Version]
- Mogensen, T.H. Pathogen recognition and inflammatory signaling in innate immune defenses. Clin. Microbiol. Rev. 2009, 22, 240–273. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef] [Green Version]
- Sakai, J.; Cammarota, E.; Wright, J.A.; Cicuta, P.; Gottschalk, R.A.; Li, N.; Fraser, I.D.; Bryant, C.E. Lipopolysaccharide-induced NF-κB nuclear translocation is primarily dependent on MyD88, but TNFα expression requires TRIF and MyD88. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef]
- Christian, F.; Smith, E.L.; Carmody, R.J. The regulation of NF-kappaB subunits by phosphorylation. Cells 2016, 5, 12. [Google Scholar] [CrossRef] [Green Version]
- Qiao, Y.; He, H.; Jonsson, P.; Sinha, I.; Zhao, C.; Dahlman-Wright, K. AP-1 is a key regulator of proinflammatory cytokine TNFalpha-mediated triple-negative breast cancer progression. J. Biol. Chem. 2016, 291, 5068–5079. [Google Scholar] [CrossRef] [Green Version]
- Natarajan, K.; Abraham, P.; Kota, R.; Isaac, B. NF-kappaB-iNOS-COX2-TNF alpha inflammatory signaling pathway plays an important role in methotrexate induced small intestinal injury in rats. Food Chem. Toxicol. 2018, 118, 766–783. [Google Scholar] [CrossRef]
- Singh, N.; Baby, D.; Rajguru, J.P.; Patil, P.B.; Thakkannavar, S.S.; Pujari, V.B. Inflammation and cancer. Ann. Afr. Med. 2019, 18, 121. [Google Scholar] [CrossRef]
- Ghasemian, M.; Owlia, S.; Owlia, M.B. Review of anti-inflammatory herbal medicines. Adv. Pharmacol. Sci. 2016, 2016, 9130979. [Google Scholar] [CrossRef] [Green Version]
- Ahn, C.; Lee, J.H.; Kim, J.W.; Park, M.J.; Lee, S.S.; Jeung, E.B. Alleviation effects of natural volatile organic compounds from Pinus densiflora and Chamaecyparis obtusa on systemic and pulmonary inflammation. Biomed. Rep. 2018, 9, 405–414. [Google Scholar] [CrossRef]
- Heo, J. Gukyeok Jeungbo Dongeui bogam; Namsandang: Seoul, Korea, 1969. [Google Scholar]
- Li, Y.Y.; Feng, J.; Zhang, X.L.; Cui, Y.Y. Pine bark extracts: Nutraceutical, pharmacological, and toxicological evaluation. J. Pharmacol. Exp. Ther. 2015, 353, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Raza, M.; Bhatt, H. Atrophic Gastritis; StatPearls: Treasure Island, FL, USA, 2021. [Google Scholar]
- Li, D.; Yang, Y.; Wang, S.; He, X.; Liu, M.; Bai, B.; Tian, C.; Sun, R.; Yu, T.; Chu, X. Role of acetylation in doxorubicin-induced cardiotoxicity. Redox Biol. 2021, 46, 102089. [Google Scholar] [CrossRef]
- Kumar, V.; Chhibber, S. Acute lung inflammation in Klebsiella pneumoniae B5055-induced pneumonia and sepsis in BALB/c mice: A comparative study. Inflammation 2011, 34, 452–462. [Google Scholar] [CrossRef]
- Park, B.S.; Lee, J.O. Recognition of lipopolysaccharide pattern by TLR4 complexes. Exp. Mol. Med. 2013, 45, e66. [Google Scholar] [CrossRef] [Green Version]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef]
- Stowell, N.C.; Seideman, J.; Raymond, H.A.; Smalley, K.A.; Lamb, R.J.; Egenolf, D.D.; Bugelski, P.J.; Murray, L.A.; Marsters, P.A.; Bunting, R.A.; et al. Long-term activation of TLR3 by poly(I:C) induces inflammation and impairs lung function in mice. Respir. Res. 2009, 10, A43. [Google Scholar] [CrossRef]
- Matute-Bello, G.; Downey, G.; Moore, B.B.; Groshong, S.D.; Matthay, M.A.; Slutsky, A.S.; Kuebler, W.M. An official American Thoracic Society workshop report: Features and measurements of experimental acute lung injury in animals. Am. J. Respir. Cell. Mol. Biol. 2011, 44, 725–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, S.A.; Park, D.W.; Kwon, J.E.; Song, H.S.; Park, B.; Jeon, H.; Sohn, E.H.; Koo, H.J.; Kang, S.C. Quinic acid inhibits vascular inflammation in TNF-alpha-stimulated vascular smooth muscle cells. Biomed. Pharmacother. 2017, 96, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.N.; Wu, W.J.; Sun, C.Z.; Liu, H.F.; Chen, W.B.; Zhan, Q.P.; Lei, Z.G.; Xin, X.; Ma, J.J.; Yao, K.; et al. Antioxidant and anti-inflammatory capacity of ferulic acid released from wheat bran by solid-state fermentation of Aspergillus niger. Biomed. Environ. Sci. 2019, 32, 11–21. [Google Scholar]
- Yoon, B.I.; Ha, U.S.; Sohn, D.W.; Lee, S.J.; Kim, H.W.; Han, C.H.; Lee, C.B.; Cho, Y.H. Anti-inflammatory and antimicrobial effects of nanocatechin in a chronic bacterial prostatitis rat model. J. Infect. Chemother. 2011, 17, 189–194. [Google Scholar] [CrossRef]
- Gupta, M.B.; Bhalla, T.N.; Gupta, G.P.; Mitra, C.R.; Bhargava, K.P. Anti-inflammatory activity of taxifolin. Jpn. J. Pharmacol. 1971, 21, 377–382. [Google Scholar] [CrossRef]
- Johnson, J.J. Carnosol: A promising anti-cancer and anti-inflammatory agent. Cancer Lett. 2011, 305, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Gu, X.; Zhang, Q.; Du, Q.; Shen, H.; Zhu, Z. Pinocembrin attenuates allergic airway inflammation via inhibition of NF-kappaB pathway in mice. Int. Immunopharmacol. 2017, 53, 90–95. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; Hoidal, J.R.; Mukherjee, T.K. Role of TNFalpha in pulmonary pathophysiology. Respir. Res. 2006, 7, A125. [Google Scholar] [CrossRef] [Green Version]
- Giuliani, C.; Bucci, I.; Napolitano, G. The role of the transcription factor nuclear factor-kappa B in thyroid autoimmunity and cancer. Front. Endocrinol. (Lausanne) 2018, 9, 471. [Google Scholar] [CrossRef] [Green Version]
- Zoumpourlis, V.; Papassava, P.; Linardopoulos, S.; Gillespie, D.; Balmain, A.; Pintzas, A. High levels of phosphorylated c-Jun, Fra-1, Fra-2 and ATF-2 proteins correlate with malignant phenotypes in the multistage mouse skin carcinogenesis model. Oncogene 2000, 19, 4011–4021. [Google Scholar] [CrossRef] [Green Version]
- Barguilla, I.; Bach, J.; Peremarti, J.; Marcos, R.; Hernandez, A. FRA1 is essential for the maintenance of the oncogenic phenotype induced by in vitro long-term arsenic exposure. Metallomics 2020, 12, 2161–2173. [Google Scholar]
- Weir, G.M.; Karkada, M.; Hoskin, D.; Stanford, M.M.; MacDonald, L.; Mansour, M.; Liwski, R.S. Combination of poly I:C and Pam3CSK4 enhances activation of B cells in vitro and boosts antibody responses to protein vaccines in vivo. PLoS ONE 2017, 12, e0180073. [Google Scholar] [CrossRef]
- Lee, J.; Rhee, M.H.; Kim, E.; Cho, J.Y. BAY 11-7082 is a broad-spectrum inhibitor with anti-inflammatory activity against multiple targets. Mediat. Inflamm. 2012, 2012, 416036. [Google Scholar] [CrossRef]
- Song, C.; Jeong, D.; Hong, Y.H.; Li, W.Y.; Lee, S.W.; Hossain, M.A.; Taamalli, A.; Kim, J.H.; Kim, J.H.; Cho, J.Y. Anti-inflammatory and photoaging-protective effects of Olea europaea through inhibition of AP-1 and NF-kappaB pathways. Am. J. Chin. Med. 2020, 48, 1895–1913. [Google Scholar] [CrossRef]
- Hossen, M.J.; Kim, M.Y.; Kim, J.H.; Cho, J.Y. AP-1-targeted inhibition of macrophage function and lipopolysaccharide/D-galactosamine-induced hepatitis by Phyllanthus acidus methanolic extract. Am. J. Chin. Med. 2015, 43, 1137–1158. [Google Scholar] [CrossRef]
- Zhang, J.; Kim, M.Y.; Cho, J.Y. Euodia pasteuriana methanol extract exerts anti-inflammatory effects by targeting TAK1 in the AP-1 signaling pathway. Molecules 2020, 25, 5760. [Google Scholar] [CrossRef] [PubMed]
- Han, S.Y.; Kim, J.; Kim, E.; Kim, S.H.; Seo, D.B.; Kim, J.H.; Shin, S.S.; Cho, J.Y. AKT-targeted anti-inflammatory activity of Panax ginseng calyx ethanolic extract. J. Ginseng Res. 2018, 42, 496–503. [Google Scholar] [CrossRef]
- Kao, K.C.; Hu, H.C.; Chang, C.H.; Hung, C.Y.; Chiu, L.C.; Li, S.H.; Lin, S.W.; Chuang, L.P.; Wang, C.W.; Li, L.F.; et al. Diffuse alveolar damage associated mortality in selected acute respiratory distress syndrome patients with open lung biopsy. Crit. Care. 2015, 19, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.O.; Kim, J.H.; Kim, S.; Kim, M.Y.; Hong, Y.H.; Kim, H.G.; Cho, J.Y. Gastroprotective effects of the nonsaponin fraction of Korean Red Ginseng through cyclooxygenase-1 upregulation. J. Ginseng Res. 2020, 44, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.; Lee, J.S.; Jang, Y.J.; Choung, E.S.; Li, W.Y.; Lee, S.W.; Kim, E.; Kim, J.H.; Cho, J.Y. Sorbaria kirilowii ethanol extract exerts anti-inflammatory effects in vitro and in vivo by targeting Src/Nuclear factor (NF)-kappaB. Biomolecules 2020, 10, 741. [Google Scholar] [CrossRef] [PubMed]
- Won, J.H.; Im, H.T.; Kim, Y.H.; Yun, K.J.; Park, H.J.; Choi, J.W.; Lee, K.T. Anti-inflammatory effect of buddlejasaponin IV through the inhibition of iNOS and COX-2 expression in RAW 264.7 macrophages via the NF-kappaB inactivation. Br. J. Pharmacol. 2006, 148, 216–225. [Google Scholar] [CrossRef] [Green Version]
- Fujioka, S.; Niu, J.; Schmidt, C.; Sclabas, G.M.; Peng, B.; Uwagawa, T.; Li, Z.; Evans, D.B.; Abbruzzese, J.L.; Chiao, P.J. NF-kappaB and AP-1 connection: Mechanism of NF-kappaB-dependent regulation of AP-1 activity. Mol. Cell Biol. 2004, 24, 7806–7819. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.F.; Liao, K.C.; Chen, C.H. 2-Phenylnaphthalene derivatives inhibit lipopolysaccharide-induced pro-inflammatory mediators by downregulating of MAPK/NF-kappaB pathways in RAW 264.7 macrophage cells. PLoS ONE 2017, 12, e0168945. [Google Scholar]
- Zhu, T.; Chen, J.; Zhao, Y.; Zhang, J.; Peng, Q.; Huang, J.; Luo, J.; Zhang, W. Neuromedin B mediates IL-6 and COX-2 expression through NF-kappaB/P65 and AP-1/C-JUN activation in human primary myometrial cells. Biosci. Rep. 2019, 39, 20192139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Wan, Y.; Huang, C. The biological functions of NF-kappaB1 (p50) and its potential as an anti-cancer target. Curr. Cancer Drug Targets. 2009, 9, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Grotsch, B.; Brachs, S.; Lang, C.; Luther, J.; Derer, A.; Schlotzer-Schrehardt, U.; Bozec, A.; Fillatreau, S.; Berberich, I.; Hobeika, E.; et al. The AP-1 transcription factor Fra1 inhibits follicular B cell differentiation into plasma cells. J. Exp. Med. 2014, 211, 2199–2212. [Google Scholar] [CrossRef] [PubMed]
- Whitmarsh, A.J.; Davis, R.J. Transcription factor AP-1 regulation by mitogen-activated protein kinase signal transduction pathways. J. Mol. Med. (Berl) 1996, 74, 589–607. [Google Scholar] [CrossRef]
- Zhang, J.; Rho, Y.; Kim, M.Y.; Cho, J.Y. TAK1 in the AP-1 pathway is a critical target of Saururus chinensis (Lour.) Baill in its anti-inflammatory action. J. Ethnopharmacol. 2021, 279, 114400. [Google Scholar] [CrossRef]
- Ha, A.T.; Kim, M.Y.; Cho, J.Y. TAK1/AP-1-Targeted anti-inflammatory effects of Barringtonia augusta methanol extract. Molecules 2021, 26, 3053. [Google Scholar] [CrossRef]
- Yi, Y.S.; Cho, J.Y.; Kim, D. Cerbera manghas methanol extract exerts anti-inflammatory activity by targeting c-Jun N-terminal kinase in the AP-1 pathway. J. Ethnopharmacol. 2016, 193, 387–396. [Google Scholar] [CrossRef]
- Song, C.; Hong, Y.H.; Park, J.G.; Kim, H.G.; Jeong, D.; Oh, J.; Sung, G.H.; Hossain, M.A.; Taamalli, A.; Kim, J.H.; et al. Suppression of Src and Syk in the NF-kappaB signaling pathway by Olea europaea methanol extract is leading to its anti-inflammatory effects. J. Ethnopharmacol. 2019, 235, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.G.; Kim, S.; Kim, H.G.; Kim, E.; Jeong, D.; Kim, J.H.; Yang, W.S.; Oh, J.; Sung, G.H.; Hossain, M.A.; et al. Mycetia cauliflora methanol extract exerts anti-inflammatory activity by directly targeting PDK1 in the NF-kappaB pathway. J. Ethnopharmacol. 2019, 231, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Han, S.Y.; Yi, Y.S.; Jeong, S.G.; Hong, Y.H.; Choi, K.J.; Hossain, M.A.; Hwang, H.; Rho, H.S.; Lee, J.; Kim, J.H.; et al. Ethanol extract of Lilium bulbs plays an anti-inflammatory role by targeting the IKKalpha/beta-mediated NF-kappaB pathway in macrophages. Am. J. Chin. Med. 2018, 46, 1281–1296. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.S.; Kim, D.; Yi, Y.S.; Kim, J.H.; Jeong, H.Y.; Hwang, K.; Kim, J.H.; Park, J.; Cho, J.Y. AKT-targeted anti-inflammatory activity of the methanol extract of Chrysanthemum indicum var. albescens. J. Ethnopharmacol. 2017, 201, 82–90. [Google Scholar] [CrossRef]
- Baik, J.S.; Seo, Y.N.; Yi, J.M.; Rhee, M.H.; Park, M.T.; Kim, S.D. Ginsenoside-Rp1 inhibits radiation-induced effects in lipopolysaccharide-stimulated J774A.1 macrophages and suppresses phenotypic variation in CT26 colon cancer cells. J. Ginseng Res. 2020, 44, 843–848. [Google Scholar] [CrossRef]
- Lee, C.Y.; Kim, H.G.; Park, S.H.; Jang, S.G.; Park, K.J.; Kim, D.S.; Kim, J.H.; Cho, J.Y. Anti-inflammatory functions of alverine via targeting Src in the NF-κB pathway. Biomolecules 2020, 10, 611. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.S.; Zhu, H.; Li, H.; Li, Y.; Zhao, B.; Jin, Y.H. Ginsenoside compound K inhibits nuclear factor-kappa B by targeting annexin A2. J. Ginseng Res. 2019, 43, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhong, S.; Hou, L.; Wang, Y.; Xing, X.; Guan, T.; Zhang, J.; Li, T. Computational and experimental characterization of estrogenic activities of 20(S, R)-protopanaxadiol and 20(S, R)-protopanaxatriol. J. Ginseng Res. 2020, 44, 690–696. [Google Scholar] [CrossRef]
- Howe, J.G.; Hershey, J.W. A sensitive immunoblotting method for measuring protein synthesis initiation factor levels in lysates of Escherichia coli. J. Biol. Chem. 1981, 256, 12836–12839. [Google Scholar] [CrossRef]
- Lorz, L.R.; Kim, D.; Kim, M.Y.; Cho, J.Y. Panax ginseng-derived fraction BIOGF1K reduces atopic dermatitis responses via suppression of mitogen-activated protein kinase signaling pathway. J. Ginseng Res. 2020, 44, 453–460. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Score per Field | ||
|---|---|---|---|
| 0 | 1 | 2 | |
| A. Neutrophils in the alveolar space | None | 1–5 | >5 |
| B. Neutrophils in the interstitial space | None | 1–5 | >5 |
| C. Hyaline membranes | None | 3 | >3 |
| D. Alveolar septal thickening | < 2× | 2–4 × | >4× |
| Score = [(20 × A) + (14 × B) + (7 × C) + (2 × D)] / (number of fields × 100) | |||
| Gene | Direction | Sequences (5′ to 3′) |
|---|---|---|
| iNOS (semi-RT-PCR) | Forward Reverse | TGCCAGGGTCACAACTTTACA ACCCCAAGCAAGACTTGGAC |
| iNOS (qPCR) | Forward Reverse | GCCACCAACAATGGCAACAT TCGATGCACAACTGGGTGAA |
| COX-2 (semi-RT-PCR) | Forward Reverse | TCACGTGGAGTCCGCTTTAC TTCGACAGGAAGGGGATGTT |
| COX-2 (qPCR) | Forward Reverse | TTGGAGGCGAAGTGGGTTTT TGGCTGTTTTGGTAGGCTGT |
| IL-6 (semi-RT-PCR) | Forward Reverse | GCCTTCTTGGGACTGATGG TGGAAATTGGGGTAGGAAGGAC |
| IL-6 (qPCR) | Forward Reverse | AGCCAGAGTCCTTCAGAGAGA AGGAGAGCATTGGAAATTGGGG |
| TNF-α (semi-RT-PCR) | Forward Reverse | TGCCTATGTCTCAGCCTCTT GAGGCCATTTGGGAACTTCT |
| GAPDH (semi-RT-PCR) | Forward Reverse | GAAGGTCGGTGTGAACGGAT AGTGATGGCATGGACTGTGG |
| GAPDH (qPCR) | Forward Reverse | TGTTGAACGGATTTGGCCGTA ACTGTGCCGTTGAATTTGCC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.A.; Oh, J.; Choi, S.R.; Lee, C.H.; Lee, B.-H.; Lee, M.-N.; Hossain, M.A.; Kim, J.-H.; Lee, S.; Cho, J.Y. Anti-Gastritis and Anti-Lung Injury Effects of Pine Tree Ethanol Extract Targeting Both NF-κB and AP-1 Pathways. Molecules 2021, 26, 6275. https://doi.org/10.3390/molecules26206275
Kim SA, Oh J, Choi SR, Lee CH, Lee B-H, Lee M-N, Hossain MA, Kim J-H, Lee S, Cho JY. Anti-Gastritis and Anti-Lung Injury Effects of Pine Tree Ethanol Extract Targeting Both NF-κB and AP-1 Pathways. Molecules. 2021; 26(20):6275. https://doi.org/10.3390/molecules26206275
Chicago/Turabian StyleKim, Seung A, Jieun Oh, Se Rin Choi, Choong Hwan Lee, Byoung-Hee Lee, Mi-Nam Lee, Mohammad Amjad Hossain, Jong-Hoon Kim, Sarah Lee, and Jae Youl Cho. 2021. "Anti-Gastritis and Anti-Lung Injury Effects of Pine Tree Ethanol Extract Targeting Both NF-κB and AP-1 Pathways" Molecules 26, no. 20: 6275. https://doi.org/10.3390/molecules26206275
APA StyleKim, S. A., Oh, J., Choi, S. R., Lee, C. H., Lee, B.-H., Lee, M.-N., Hossain, M. A., Kim, J.-H., Lee, S., & Cho, J. Y. (2021). Anti-Gastritis and Anti-Lung Injury Effects of Pine Tree Ethanol Extract Targeting Both NF-κB and AP-1 Pathways. Molecules, 26(20), 6275. https://doi.org/10.3390/molecules26206275