
A Rapid, Reliable RP-UPLC Method for Large-Scale Analysis of Wheat HMW-GS Alleles

Abstract
:1. Introduction
2. Results and Discussion
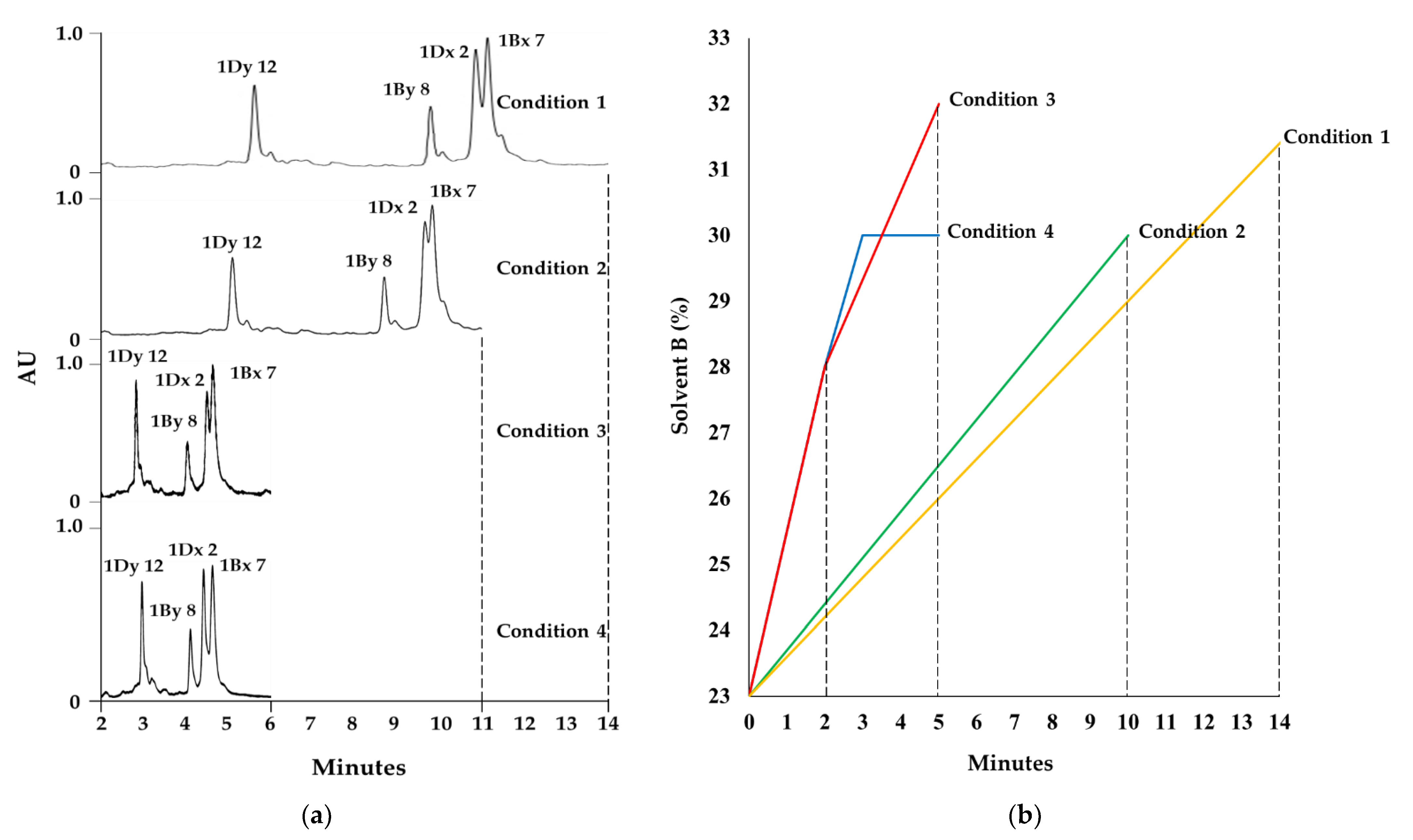
2.1. Optimization of RP-UPLC Conditions
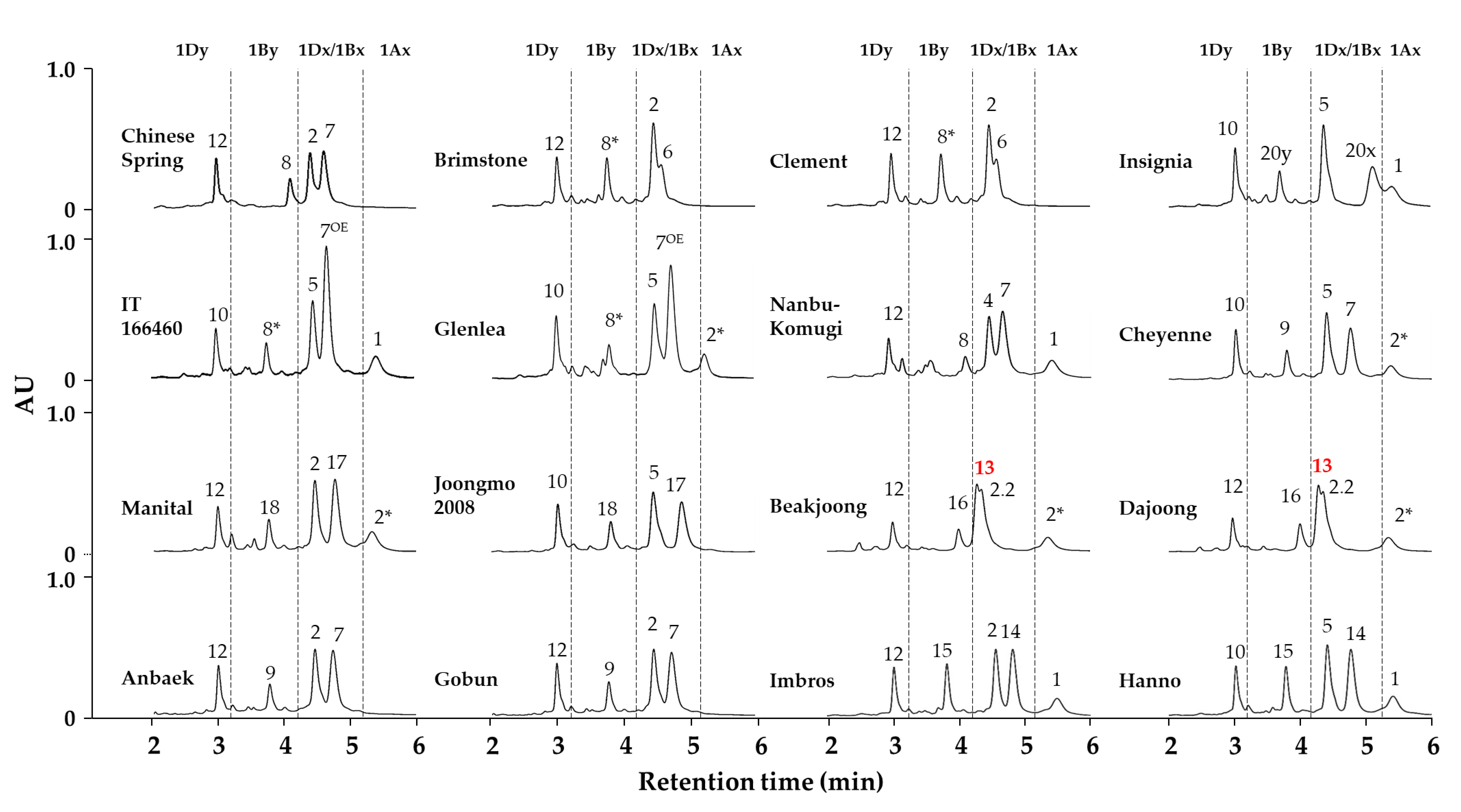
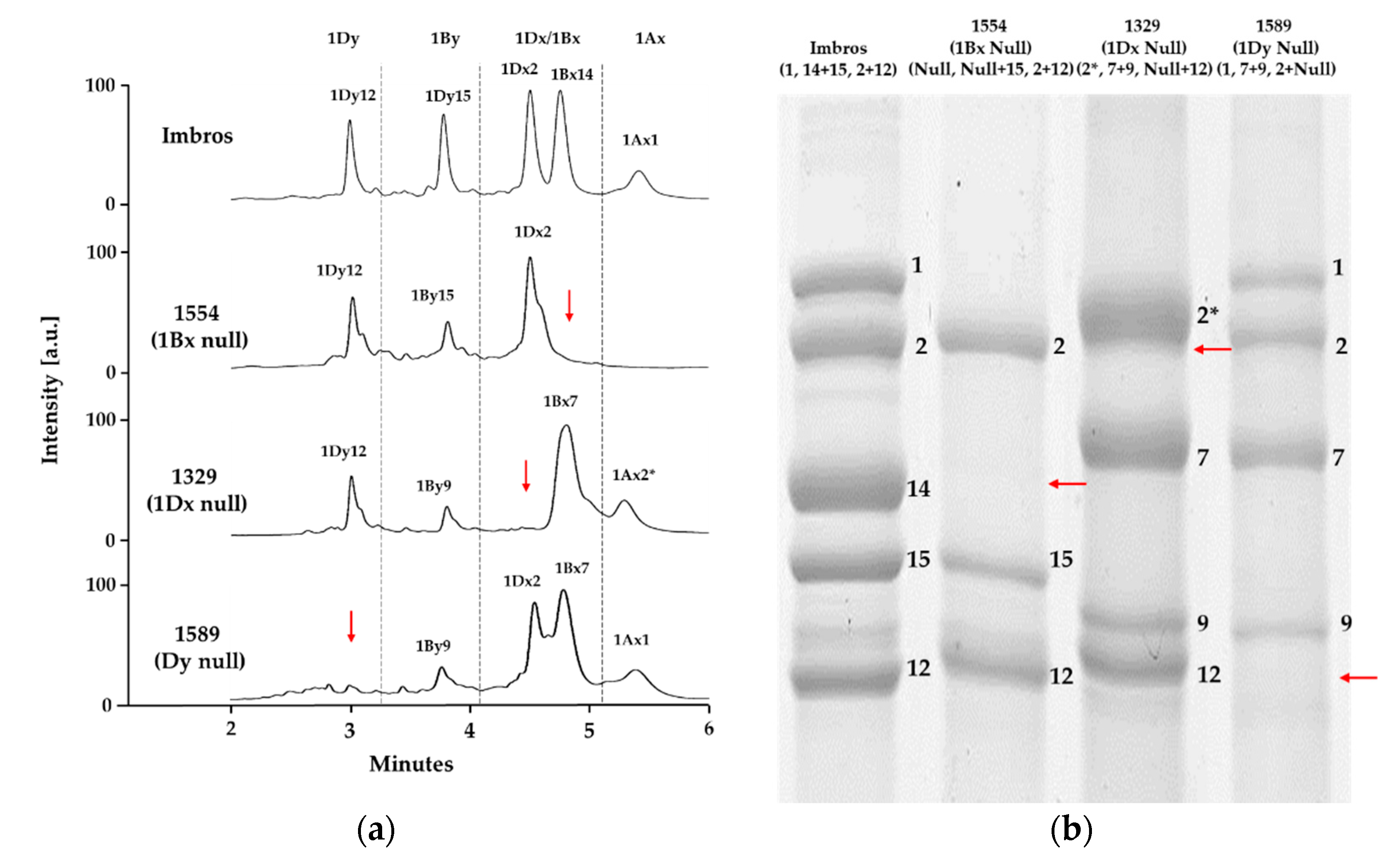
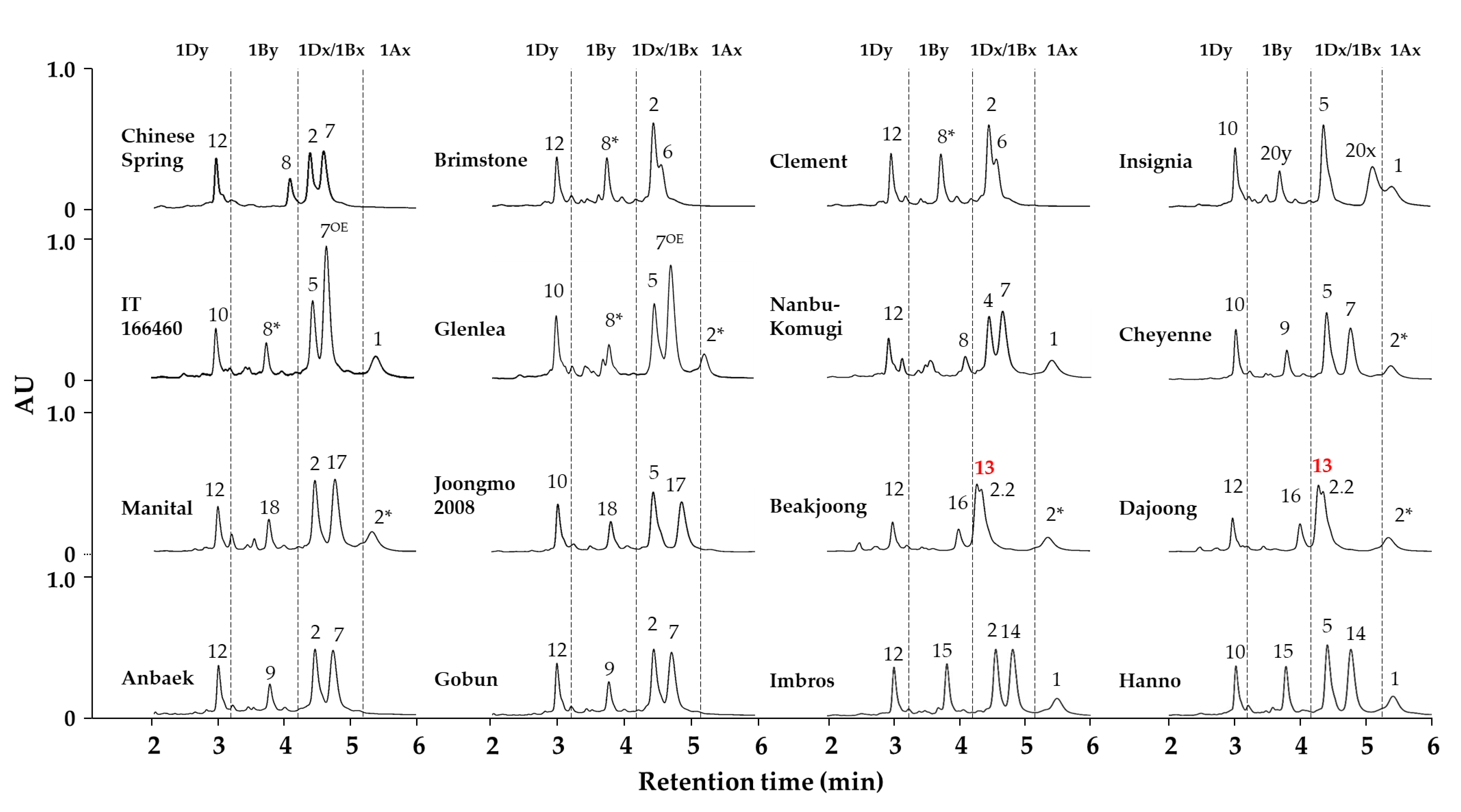
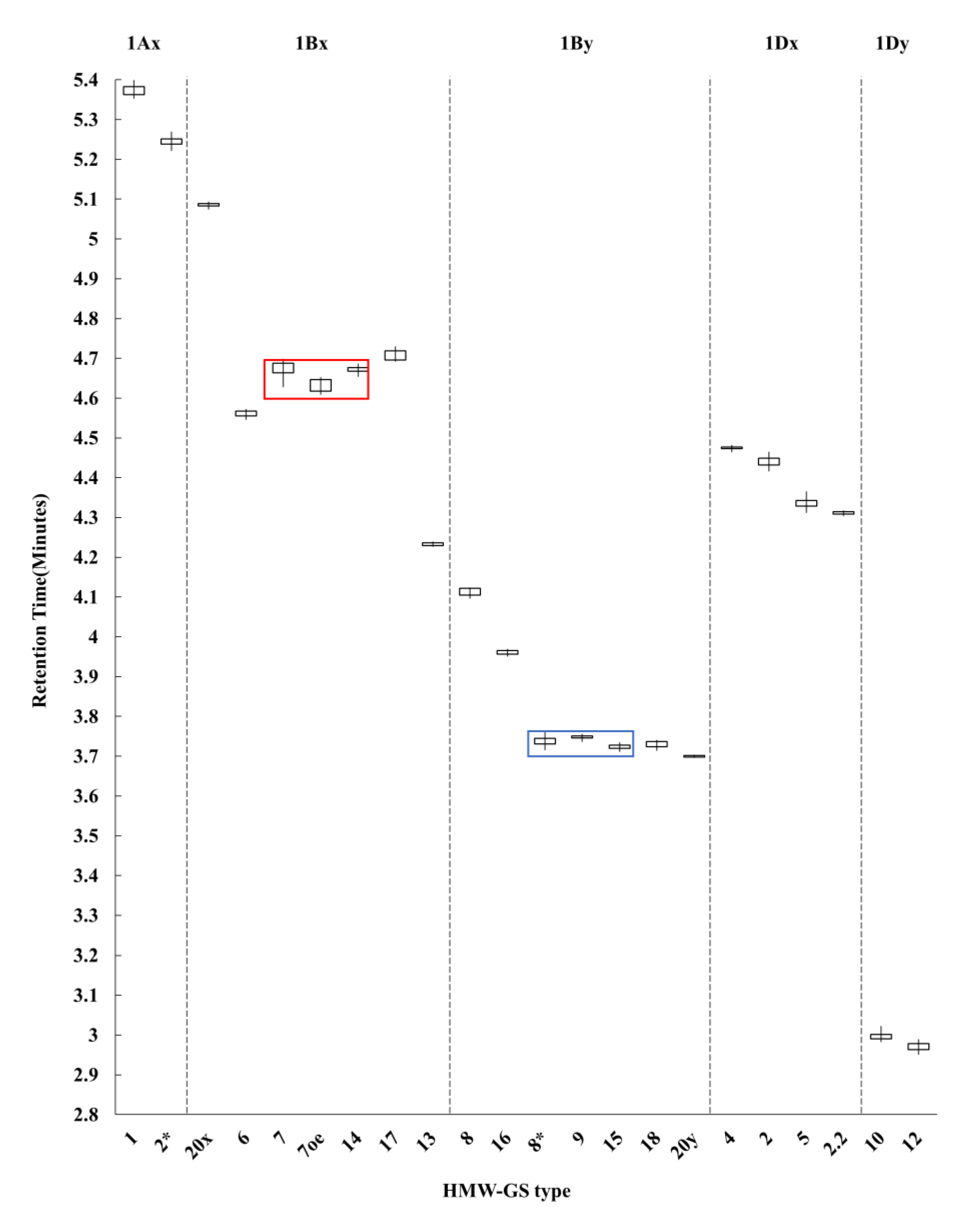
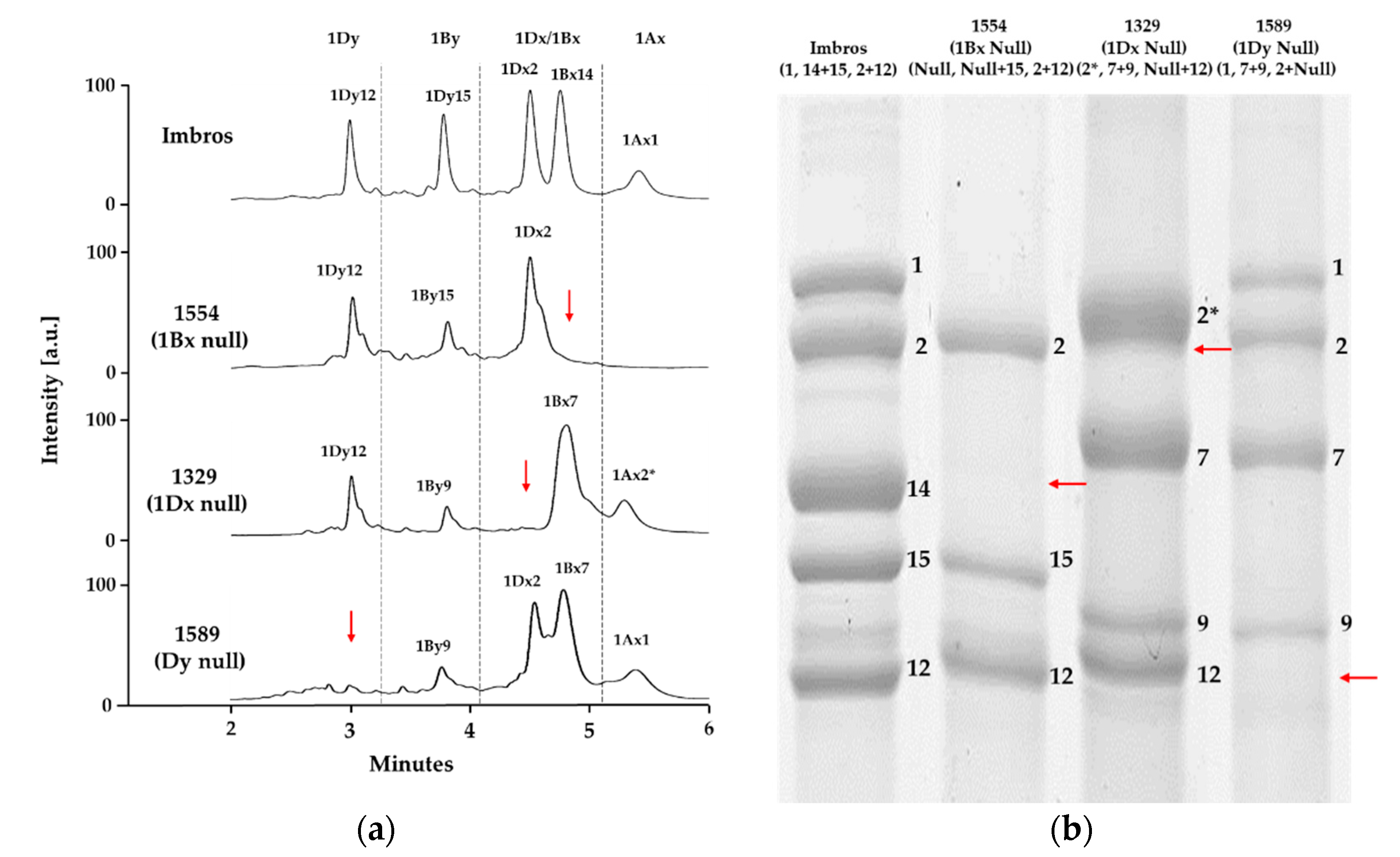
2.2. Identification of HMW-GS Compositions in Standard Wheat Cultivars
2.3. Applying the Optimized Method to a Large Collection of Wheat Germplasm
3. Materials and Methods
3.1. Plant Materials
3.2. Glutenin Extraction and Precipitation
3.3. RP-UPLC
3.4. SDS-PAGE
3.5. Genomic DNA Extraction and PCR Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Payne, P.I. Genetics of wheat storage proteins and the effect of allelic variation on bread-making quality. Annu. Rev. Plant Physiol. 1987, 38, 141–153. [Google Scholar] [CrossRef]
- Macritchie, F. Physicochemical Properties of Wheat Proteins in Relation to Functionality. In Advances in Food and Nutrition Research; Kinsella, J.E., Ed.; Academic Press: Cambridge, MA, USA, 1992; Volume 36, pp. 1–87. [Google Scholar]
- Wieser, H. Chemistry of gluten proteins. Food Microbiol. 2007, 24, 115–119. [Google Scholar] [CrossRef]
- D’Ovidio, R.; Masci, S. The low-molecular-weight glutenin subunits of wheat gluten. J. Cereal Sci. 2004, 39, 321–339. [Google Scholar] [CrossRef]
- Jang, Y.-R.; Cho, K.; Kim, S.W.; Altenbach, S.B.; Lim, S.-H.; Sim, J.-R.; Lee, J.-Y. Development of an Optimized MALDI-TOF-MS Method for High-Throughput Identification of High-Molecular-Weight Glutenin Subunits in Wheat. Molecules 2020, 25, 4347. [Google Scholar] [CrossRef] [PubMed]
- Wrigley, C.W. Giant proteins with flour power. Nature 1996, 381, 738–739. [Google Scholar] [CrossRef]
- Payne, P.I.; Lawrence, G.J. Catalogue of alleles for the complex gene loci, Glu-A1, Glu-B1, and Glu-D1 which code for high-molecular-weight subunits of glutenin in hexaploid wheat. Cereal Res. Commun. 1983, 11, 29–35. [Google Scholar]
- Shewry, P.; Halford, N.; Tatham, A. High molecular weight subunits of wheat glutenin. J. Cereal Sci. 1992, 15, 105–120. [Google Scholar] [CrossRef]
- Rakszegi, M.; Bekes, F.; Lang, L.; Tamas, L.; Shewry, P.; Bedő, Z. Technological quality of transgenic wheat expressing an increased amount of a HMW glutenin subunit. J. Cereal Sci. 2005, 42, 15–23. [Google Scholar] [CrossRef]
- Barro, F.; Rooke, L.; Békés, F.; Gras, P.; Tatham, A.S.; Fido, R.; Lazzeri, P.A.; Shewry, P.R.; Barceló, P. Transformation of wheat with high molecular weight subunit genes results in improved functional properties. Nat. Biotechnol. 1997, 15, 1295–1299. [Google Scholar] [CrossRef]
- Payne, P.I.; Nightingale, M.A.; Krattiger, A.F.; Holt, L.M. The relationship between HMW glutenin subunit composition and the bread-making quality of British-grown wheat varieties. J. Sci. Food Agric. 1987, 40, 51–65. [Google Scholar] [CrossRef]
- Rogers, W.; Payne, P.; Harinder, K. The HMW glutenin subunit and gliadin compositions of German-grown wheat varieties and their relationship with bread-making quality. Plant Breed. 1989, 103, 89–100. [Google Scholar] [CrossRef]
- Gianibelli, M.; Larroque, O.; MacRitchie, F.; Wrigley, C. Biochemical, genetic, and molecular characterization of wheat endosperm proteins. Cereal Chem. 2001, 78, 635–646. [Google Scholar] [CrossRef]
- Vawser, M.; Cornish, G.B. Over-expression of HMW glutenin subunit Glu-B1 7x in hexaploid wheat varieties (Triticum aestivum). Aust. J. Agric. Res. 2004, 55, 577–588. [Google Scholar] [CrossRef]
- Butow, B.; Ma, W.; Gale, K.; Cornish, G.; Rampling, L.; Larroque, O.; Morell, M.; Békés, F. Molecular discrimination of Bx7 alleles demonstrates that a highly expressed high-molecular-weight glutenin allele has a major impact on wheat flour dough strength. Theor. Appl. Genet. 2003, 107, 1524–1532. [Google Scholar] [CrossRef]
- Radovanovic, N.; Cloutier, S.; Brown, D.; Humphreys, D.; Lukow, O. Genetic variance for gluten strength contributed by high molecular weight glutenin proteins. Cereal Chem. 2002, 79, 843–849. [Google Scholar] [CrossRef]
- Ragupathy, R.; Naeem, H.A.; Reimer, E.; Lukow, O.M.; Sapirstein, H.D.; Cloutier, S. Evolutionary origin of the segmental duplication encompassing the wheat GLU-B1 locus encoding the overexpressed Bx7 (Bx7 OE) high molecular weight glutenin subunit. Theor. Appl. Genet. 2008, 116, 283–296. [Google Scholar] [CrossRef]
- Bietz, J.A. Separation of cereal proteins by reversed-phase high-performance liquid chromatography. J. Chromatogr. A 1983, 255, 219–238. [Google Scholar] [CrossRef]
- Courcoux, P.; Serot, T.; Larre, C.; Popineau, Y. Characterization and identification of wheat cultivars by multi-dimensional analysis of reversed-phase high-performance liquid chromatograms. J. Chromatogr. A 1992, 596, 225–232. [Google Scholar] [CrossRef]
- Cozzolino, R.; Giorgi, S.D.; Fisichella, S.; Garozzo, D.; Lafiandra, D.; Palermo, A. Matrix-assisted laser desorption/ionization mass spectrometric peptide mapping of high molecular weight glutenin subunits 1Bx7 and 1Dy10 in Cheyenne cultivar. Rapid Commun. Mass Spectrom. 2001, 15, 778–787. [Google Scholar] [CrossRef]
- Dong, K.; Hao, C.; Wang, A.; Cai, M.; Yan, Y. Characterization of HMW glutenin subunits in bread and tetraploid wheats by reversed-phase high-performance liquid chromatography. Cereal Res. Commun. 2009, 37, 65–73. [Google Scholar] [CrossRef]
- Gao, L.; Ma, W.; Chen, J.; Wang, K.; Li, J.; Wang, S.; Bekes, F.; Appels, R.; Yan, Y. Characterization and comparative analysis of wheat high molecular weight glutenin subunits by SDS-PAGE, RP-HPLC, HPCE, and MALDI-TOF-MS. J. Agric. Food Chem. 2010, 58, 2777–2786. [Google Scholar] [CrossRef]
- Yan, X.; Liu, W.; Yu, Z.; Han, C.; Zeller, F.J.; Hsam, S.L.; Yan, Y. Rapid separation and identification of wheat HMW glutenin subunits by UPLC and comparative analysis with HPLC. Aust. J. Crop Sci. 2014, 8, 140–147. [Google Scholar]
- Zhang, Q.; Dong, Y.; An, X.; Wang, A.; Zhang, Y.; Li, X.; Gao, L.; Xia, X.; He, Z.; Yan, Y. Characterization of HMW glutenin subunits in common wheat and related species by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF-MS). J. Cereal Sci. 2008, 47, 252–261. [Google Scholar] [CrossRef]
- Jang, Y.-R.; Beom, H.-R.; Altenbach, S.B.; Lee, M.-K.; Lim, S.-H.; Lee, J.-Y. Improved method for reliable HMW-GS identification by RP-HPLC and SDS-PAGE in common wheat cultivars. Molecules 2017, 22, 1055. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.-R.; Kim, S.; Sim, J.-R.; Lee, S.-B.; Lim, S.-H.; Kang, C.-S.; Choi, C.; Goo, T.-W.; Lee, J.-Y. High-throughput analysis of high-molecular weight glutenin subunits in 665 wheat genotypes using an optimized MALDI-TOF–MS method. 3 Biotech 2021, 11, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chandra, D.; Timmick, S.; Goodwine, C.; Vecchiarello, N.; Shastry, D.G.; Mullerpatan, A.; Trasatti, J.P.; Cramer, S.; Karande, P. Design of peptide ligands for affinity purification of human growth hormone. J. Chem. Technol. Biotechnol. 2019, 94, 2345–2354. [Google Scholar] [CrossRef]
- Outlaw, V.K.; Kreitler, D.F.; Stelitano, D.; Porotto, M.; Moscona, A.; Gellman, S.H. Effects of Single α-to-β Residue Replacements on Recognition of an Extended Segment in a Viral Fusion Protein. ACS Infect. Dis. 2020, 6, 2017–2022. [Google Scholar] [CrossRef]
- Perdomo-Abúndez, F.C.; Vallejo-Castillo, L.; Vázquez-Leyva, S.; López-Morales, C.A.; Velasco-Velázquez, M.; Pavón, L.; Pérez-Tapia, S.M.; Medina-Rivero, E. Development and validation of a mass spectrometric method to determine the identity of rituximab based on its microheterogeneity profile. J. Chromatogr. B 2020, 1139, 121885. [Google Scholar] [CrossRef]
- Timmick, S.M.; Vecchiarello, N.; Goodwine, C.; Crowell, L.E.; Love, K.R.; Love, J.C.; Cramer, S.M. An impurity characterization based approach for the rapid development of integrated downstream purification processes. Biotechnol. Bioeng. 2018, 115, 2048–2060. [Google Scholar] [CrossRef]
- Zhang, B.; Jeong, J.; Burgess, B.; Jazayri, M.; Tang, Y.; Zhang, Y.T. Development of a rapid RP-UHPLC–MS method for analysis of modifications in therapeutic monoclonal antibodies. J. Chromatogr. B 2016, 1032, 172–181. [Google Scholar] [CrossRef]
- Liu, L.; Ikeda, T.M.; Branlard, G.; Pena, R.J.; Rogers, W.F.; Lerner, S.E.; Kolman, M.A.; Xia, X.; Wang, L.; Ma, W. Comparison of low molecular weight glutenin subunits identified by SDS-PAGE, 2-DE, MALDI-TOF-MS and PCR in common wheat. BMC Plant Biol 2010, 10, 124–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupont, F.; Chan, R.; Lopez, R. Molar fractions of high-molecular-weight glutenin subunits are stable when wheat is grown under various mineral nutrition and temperature regimens. J. Cereal Sci. 2007, 45, 134–139. [Google Scholar] [CrossRef]
- Naeem, H.A.; Sapirstein, H.D. Ultra-fast separation of wheat glutenin subunits by reversed-phase HPLC using a superficially porous silica-based column. J. Cereal Sci. 2007, 46, 157–168. [Google Scholar] [CrossRef]
- Kazman, M.; Lein, V. Cytological and SDS-PAGE characterization of 1994-95-grown European wheat cultivars. Ann. Wheat Newslett. 1996, 42, 86–92. [Google Scholar]
- Branlard, G.; Dardevet, M.; Amiour, N.; Igrejas, G. Allelic diversity of HMW and LMW glutenin subunits and omega-gliadins in French bread wheat (Triticum aestivum L.). Genet. Resour. Crop Evol. 2003, 50, 669–679. [Google Scholar] [CrossRef]
- Cho, S.-W.; Roy, S.K.; Chun, J.-B.; Cho, K.; Park, C.S. Overexpression of the Bx7 high molecular weight glutenin subunit on the Glu-B1 locus in a Korean wheat landrace. Plant Biotechnol. Rep. 2017, 11, 97–105. [Google Scholar] [CrossRef]
- Lerner, S.; Ponzio, N.; Rogers, W. Relationship of Over-Expression of High Molecular Weight Glutenin Subunit Bx7 with Gluten Strength. In Proceedings of the 10th International Wheat Genetics Symposium, Paestum, Italy, 1–6 September 2003; pp. 1360–1362. [Google Scholar]
- Zheng, W.; Peng, Y.; Ma, J.; Appels, R.; Sun, D.; Ma, W. High frequency of abnormal high molecular weight glutenin alleles in Chinese wheat landraces of the Yangtze-River region. J. Cereal Sci. 2011, 54, 401–408. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.; Shepherd, K.; Cornish, G. A Simplified SDS-PAGE Procedure for Separating. J. Cereal Sci. 1991, 14, 203–208. [Google Scholar] [CrossRef]
- Melas, V.; Morel, M.-H.; Autran, J.-C.; Feillet, P. Simple and rapid method for purifying low molecular weight subunits of glutenin from wheat. Cereal Chem. 1994, 71, 234–237. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | HMW-GS | References | ||
|---|---|---|---|---|
| Glu-A1 | Glu-B1 | Glu-D1 | ||
| Anbaek | Null | 7 + 9 | 2 + 12 | Jang et al. [5,25] |
| Baekjoong | 2* | 13 + 16 | 2.2 + 12 | Jang et al. [5,25] |
| Brimstone | Null | 6 + 8* | 2 + 12 | Liu et al. [32] and Jang et al. [5,25] |
| Cheyenne | 2* | 7 + 9 | 5 + 10 | Dupont et al. [33] and Jang et al. [5,25] |
| Chinese Spring | Null | 7 + 8 | 2 + 12 | Liu et al. [32] and Jang et al. [5,25] |
| Clement | Null | 6 + 8* | 2 + 12 | Liu et al. [32] and Jang et al. [5,25] |
| Dajoong | 2* | 13 + 16 | 2.2 + 12 | Jang et al. [5,25] |
| Glenlea | 2* | 7OE + 8* | 5 + 10 | Naeem and Sapirstein [34] and Jang et al. [5,25] |
| Gobun | Null | 7 + 9 | 2 + 12 | Jang et al. [5,25] |
| Hanno | 1 | 14 + 15 | 5 + 10 | Gao et al. [22] and Jang et al. [5] |
| Imbros | 1 | 14 + 15 | 2 + 12 | Kazman et al. [35] and Jang et al. [5] |
| Insignia | 1 | 20 + 20 | 5 + 10 | Branlard et al. [36] and Jang et al. [5,25] |
| IT166460 | 1 | 7OE + 8* | 2 + 12 | Cho et al. [37] and Jang et al. [5,25] |
| Joongmo 2008 | Null | 17 + 18 | 5 + 10 | Jang et al. [5,25] |
| Manital | 2* | 17 + 18 | 2 + 12 | Liu et al. [32] and Jang et al. [5] |
| Nanbu-komugi | 1 | 7 + 8 | 4 + 12 | Liu et al. [32] and Jang et al. [5,25] |
| Type | HMW-GS | Retention Time (min) 1 | RSD (%) 2 | Number of Cultivars 3 |
|---|---|---|---|---|
| Ax | 1 | 5.37 ± 0.01 | 0.25 | 5 |
| 2* | 5.24 ± 0.01 | 0.22 | 5 | |
| Bx | 6 | 4.56 ± 0.01 | 0.19 | 2 |
| 7 | 4.67 ± 0.02 | 0.42 | 5 | |
| 7OE | 4.63 ± 0.02 | 0.35 | 2 | |
| 13 | 4.23 ± 0.01 | 0.10 | 2 | |
| 14 | 4.67 ± 0.01 | 0.16 | 2 | |
| 17 | 4.71 ± 0.01 | 0.27 | 2 | |
| 20x | 5.09 ± 0.01 | 0.11 | 1 | |
| By | 8 | 4.11 ± 0.01 | 0.24 | 2 |
| 8* | 3.74 ± 0.01 | 0.31 | 4 | |
| 9 | 3.75 ± 0.01 | 0.12 | 3 | |
| 15 | 3.72 ± 0.01 | 0.17 | 2 | |
| 16 | 3.96 ± 0.01 | 0.14 | 2 | |
| 18 | 3.73 ± 0.01 | 0.23 | 2 | |
| 20y | 3.70 ± 0.01 | 0.07 | 1 | |
| Dx | 2 | 4.44 ± 0.01 | 0.26 | 8 |
| 2.2 | 4.31 ± 0.01 | 0.11 | 2 | |
| 4 | 4.48 ± 0.01 | 0.12 | 1 | |
| 5 | 4.34 ± 0.01 | 0.32 | 5 | |
| Dy | 10 | 3.00 ± 0.01 | 0.39 | 5 |
| 12 | 2.97 ± 0.01 | 0.33 | 11 |
| Locus | HMW-GS | Allele * | Number of Cultivars | Frequency (%) |
|---|---|---|---|---|
| Glu-A1 | 1 | Glu-A1a | 418 | 23.39 |
| 2* | Glu-A1b | 774 | 43.31 | |
| Null | Glu-A1c | 595 | 33.30 | |
| Glu-B1 | 6 | Glu-B1-1d | 8 | 0.45 |
| 6 + 8* | Glu-B1ca | 65 | 3.64 | |
| 7 | Glu-B1a | 64 | 3.58 | |
| 7 + 8 | Glu-B1b | 473 | 26.47 | |
| 7 + 8* | Glu-B1ce | 219 | 12.26 | |
| 7 + 9 | Glu-B1c | 503 | 28.15 | |
| 7OE + 8* | Glu-B1al | 33 | 1.85 | |
| 7OE + 9 | unknown | 2 | 0.11 | |
| 8 | Glu-B1aj | 1 | 0.06 | |
| 8* | Glu-B1-2o | 1 | 0.06 | |
| 9 | Glu-B1-2b | 1 | 0.06 | |
| 13 + 16 | Glu-B1f | 67 | 3.75 | |
| 14 + 15 | Glu-B1h | 2 | 0.11 | |
| 15 | Glu-B1-2e | 7 | 0.39 | |
| 17 + 18 | Glu-B1i | 219 | 12.26 | |
| 20y | Glu-B1-2z | 2 | 0.11 | |
| 20 + 20 | Glu-B1e | 120 | 6.72 | |
| Glu-D1 | 2 + 12 | Glu-D1a | 758 | 42.42 |
| 2 | Glu-D1k | 1 | 0.06 | |
| 2.2 + 12 | Glu-D1f | 150 | 8.39 | |
| 4 + 12 | Glu-D1c | 38 | 2.13 | |
| 5 | Glu-D1-1d | 2 | 0.11 | |
| 5 + 10 | Glu-D1d | 837 | 46.84 | |
| 12 | Glu-D1l | 1 | 0.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-B.; Yang, Y.-J.; Lim, S.-H.; Gu, Y.Q.; Lee, J.-Y. A Rapid, Reliable RP-UPLC Method for Large-Scale Analysis of Wheat HMW-GS Alleles. Molecules 2021, 26, 6174. https://doi.org/10.3390/molecules26206174
Lee S-B, Yang Y-J, Lim S-H, Gu YQ, Lee J-Y. A Rapid, Reliable RP-UPLC Method for Large-Scale Analysis of Wheat HMW-GS Alleles. Molecules. 2021; 26(20):6174. https://doi.org/10.3390/molecules26206174
Chicago/Turabian StyleLee, Su-Bin, Yu-Jeong Yang, Sun-Hyung Lim, Yong Q. Gu, and Jong-Yeol Lee. 2021. "A Rapid, Reliable RP-UPLC Method for Large-Scale Analysis of Wheat HMW-GS Alleles" Molecules 26, no. 20: 6174. https://doi.org/10.3390/molecules26206174
APA StyleLee, S.-B., Yang, Y.-J., Lim, S.-H., Gu, Y. Q., & Lee, J.-Y. (2021). A Rapid, Reliable RP-UPLC Method for Large-Scale Analysis of Wheat HMW-GS Alleles. Molecules, 26(20), 6174. https://doi.org/10.3390/molecules26206174