1. Introduction
Glioblastomas (GB) are among the most common and malignant brain tumors of the central nervous system, impacting more than 10,000 people in the United States each year with a median survival rate of approximately 15 months in patients receiving treatment [
1]. Treatment typically includes a combination of radiation, neurosurgery, and chemotherapy with temozolomide (TMZ). However, despite great advances in these treatments, the prognosis for GB patients has remained relatively stable over the last three decades, with less than 5% of GB patients living past 5 years, and no reports of cured patients [
2]. Outcomes remain poor, due to resistance to treatment and a lack of other targeted therapies [
3]. Furthermore, translational efficacy of therapeutics moving from preclinical to clinical use has been particularly poor for glioblastoma.
Temozolomide is the current gold standard of treatment for GB, typically given concomitantly with radiation [
4]. However, in nearly 50% of patients this drug is ineffective, largely due to drug resistance linked to a genetic predisposition and methylation of the MGMT gene (O
6-methylguanine methyltransferase), and/or lack of a DNA repair pathway in GB cells [
5]. Due to the highly resistant, infiltrative, and aggressive nature of this disease, new treatments are essential to improve the prognosis of current and future patients.
Curcumin (diferuloylmethane) is a polyphenol isolated from the rhizome of the C. longa plant that has been shown to have anticancer effects, primarily due to its antioxidant and anti-inflammatory properties [
6]. Curcumin has been studied for the treatment of many diseases including Alzheimer’s disease [
7], diabetes [
8], hepatitis, and a variety of different cancers, including pancreatic cancer, breast cancer and glioblastoma [
9,
10,
11,
12]. While initial studies using curcumin
in vitro and with rodent models
in vivo showed great promise, human clinical trials repeatedly failed to show the same therapeutic value even at doses as high as 8 g per day [
13].
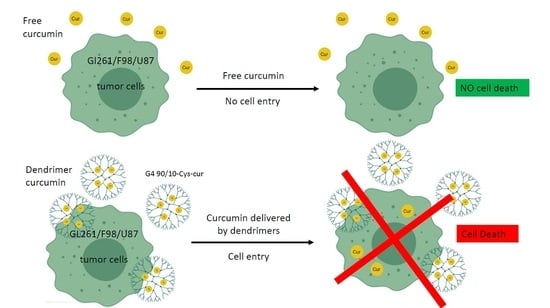
There are three major pitfalls to the use of curcumin that prevent its therapeutic actions: poor bioavailability, rapid clearance from the body, and rapid metabolism [
14]. Previous attempts to facilitate systemic delivery of curcumin, such as packaging in liposomes, have been largely unsuccessful, due to the poor stability of liposomes and the tendency of their external lipid membrane to conjugate to other lipids in the body [
15]. Curcumin alone can cross the blood brain barrier (BBB) in small amounts, but packaging curcumin inside a vehicle such as a liposome has great potential, but their size often prevented them from crossing the intact BBB. While the BBB is disrupted in glioblastoma, the tumor environment and poor vascularization around the tumor prevent even some of the smallest particles from reaching the inner core of a tumor mass [
16]. Other novel vehicles are composed of organic compounds which lack solubility in water. Therefore, vehicles that are capable of delivering therapeutics, while still maintaining a size small enough to penetrate the blood brain tumor barrier (BBTB), are needed.
Nano-molecules have been used for almost a century in industrial fields such as construction, agriculture, and carbon-based fuels; more recently, some have been adapted for use in the biomedical sciences [
17]. First conceptualized in the late 1970s, hyperbranched “starburst polymers” or “dendrimers” were first synthesized in the early 1980s [
18]. Dendrimers differ from other monomers and oligomers in their highly dendritic branching density and functional terminal end groups. Their small size (1–10 nm) and the potential to manipulate and control separate components of a dendrimer, make dendrimers suitable candidates for several biomedical applications, including delivery of genes, encapsulated non-soluble molecules, and drugs conjugated to the surface of dendrimers [
19,
20].
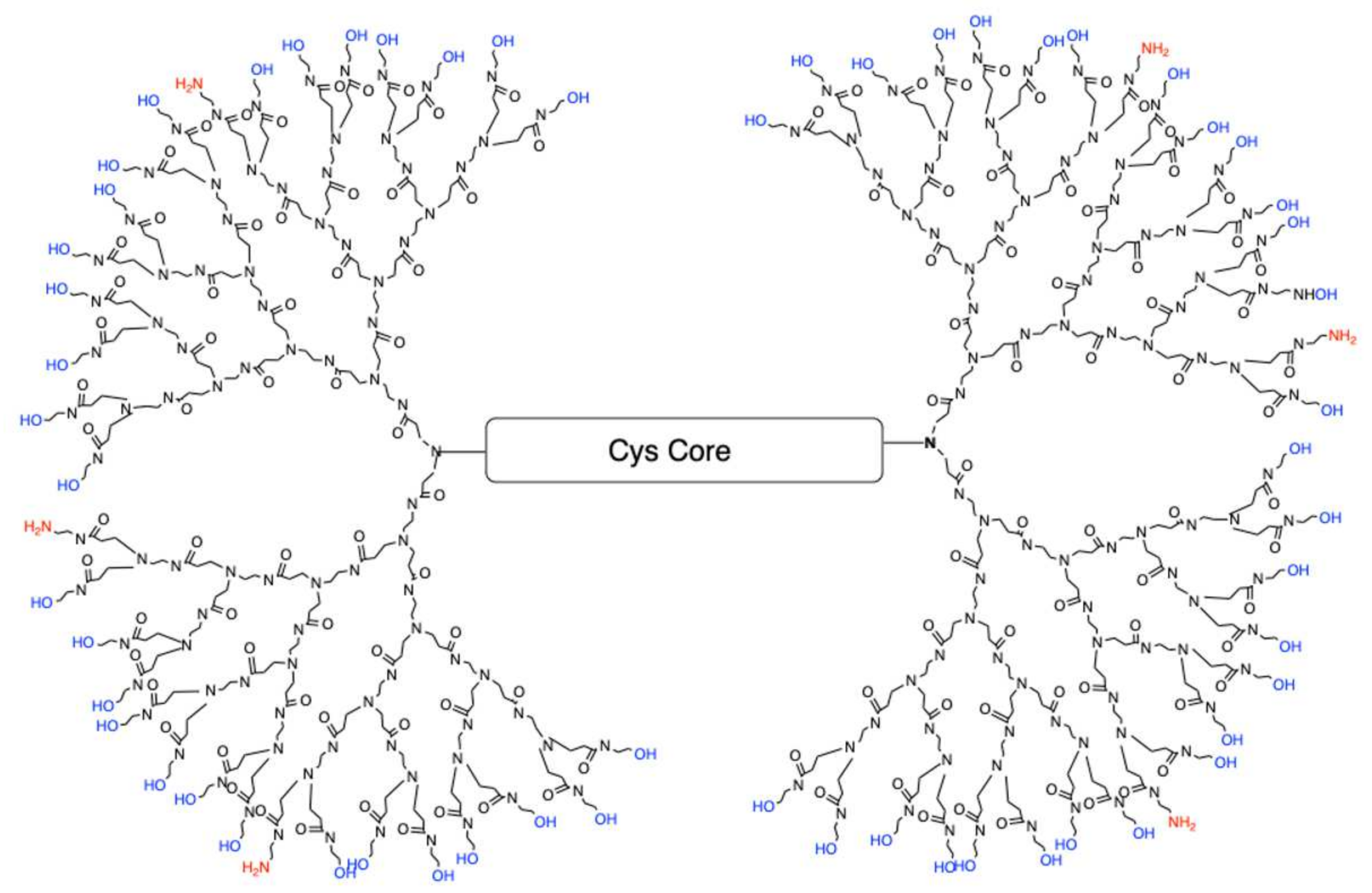
Polyamidoamine (PAMAM) dendrimers are comprised of a core, a branched interior structure, and functional surface groups. Each of these components can be modified to yield dendrimers with a unique phenotype tailored to specific therapeutic actions. Traditional fourth generation PAMAM dendrimer with 100% NH
2 surface, which exist as protonated positively charged amines (NH
3+) at physiological pH, are highly toxic to cells [
21]. Therefore, our lab has developed a reproducible
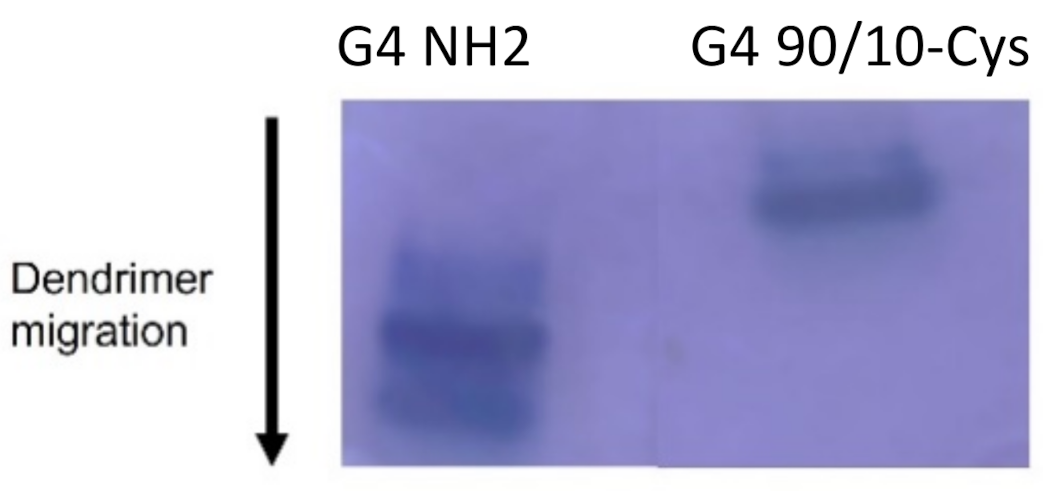
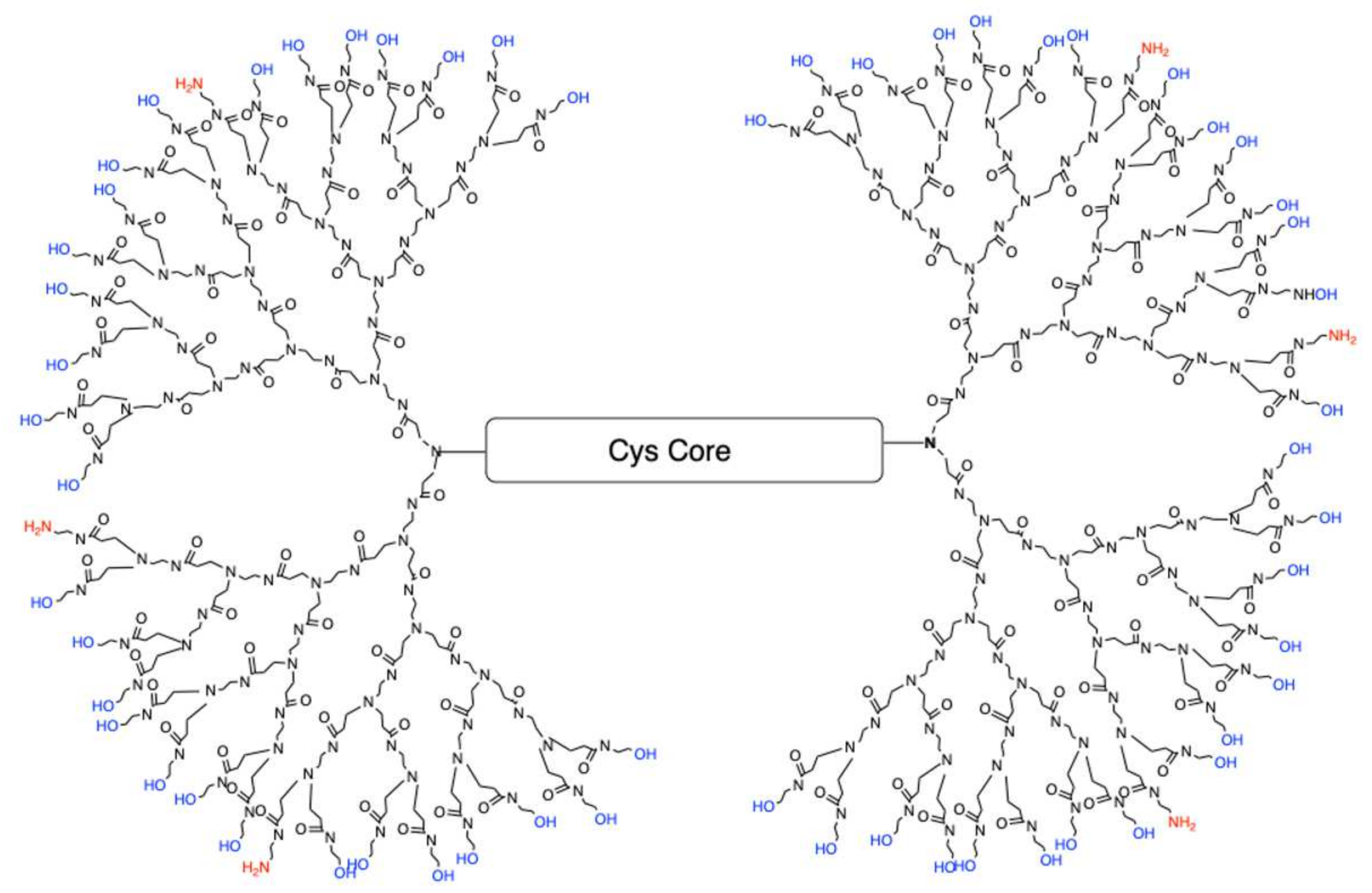
de novo method of synthesizing fourth-generation dendrimers (G4) with a modified surface containing fewer positively charged amines resulting in reduced cellular toxicity. One of the current formulations is a G4 dendrimer with 90% of the amine surface replaced with neutral hydroxyl (OH) groups referred to as G4 90/10-Cys (
Figure 1).
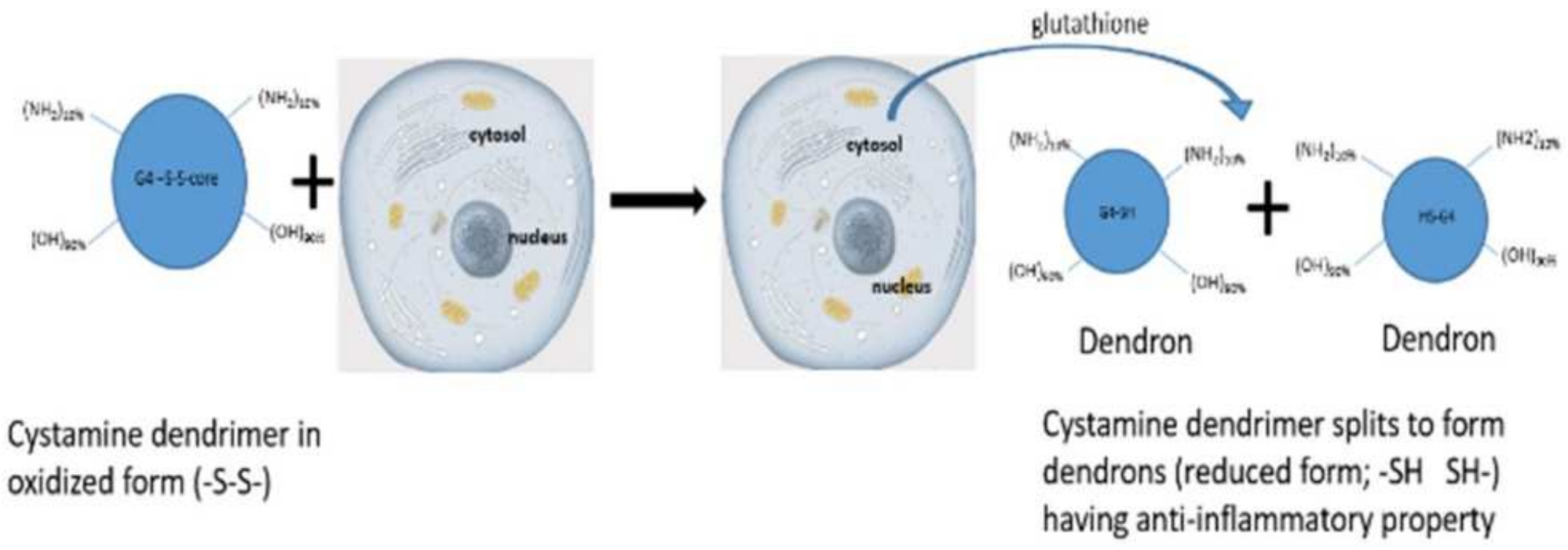
The core of G4 90/10 is composed of cystamine, with a disulfide bond holding together the two halves of the dendrimer, or “dendrons.” As shown in
Figure 2, upon entering a cell, glutathione or other reducing agents reduce and break the disulfide bond, thus splitting the dendrimer into two dendrons [
22], facilitating cargo release (e.g., curcumin) into the interior of the cell.
Recent studies suggest that the polyphenol curcumin, in addition to its many anticancer effects, may also function as a “drug resistance preventer” in certain types of cancers, including glioblastoma [
23]. Due to its potential therapeutic value, we created a novel dendrimer/curcumin formulation (G4 90/10-Cys-Cur) for use in this study. The G4 90/10-Cys-Cur has a ratio of 10:1 (
w/
w) dendrimer to curcumin. On a mole basis, the ratio is four curcumin per dendrimer. The curcumin-encapsulated dendrimer may be used as a treatment, themselves, or possibly as an adjunctive treatment to those resistant to TMZ or other chemotherapy drugs.
A common practice in drug development is to investigate the effectiveness of treatments in lower forms of mammals before moving to non-human primates and humans as test subjects. The guidelines for multiple dosing toxicity assessment were developed in 1940, which included multispecies testing as a basic requirement for new drugs, prior to clinical applications [
24]. These multispecies preclinical drug testing approaches were further encouraged by study reports about various pharmacological responses in different species to the same drug. Therefore, to account for multispecies variability, we tested our dendrimer treatments on U87-human, F98-rat, and GL261-mouse glioblastoma cell lines, and healthy control cell lines using HEK 293, rat mesenchymal stem cells (MSCs), and mouse-MSCs.
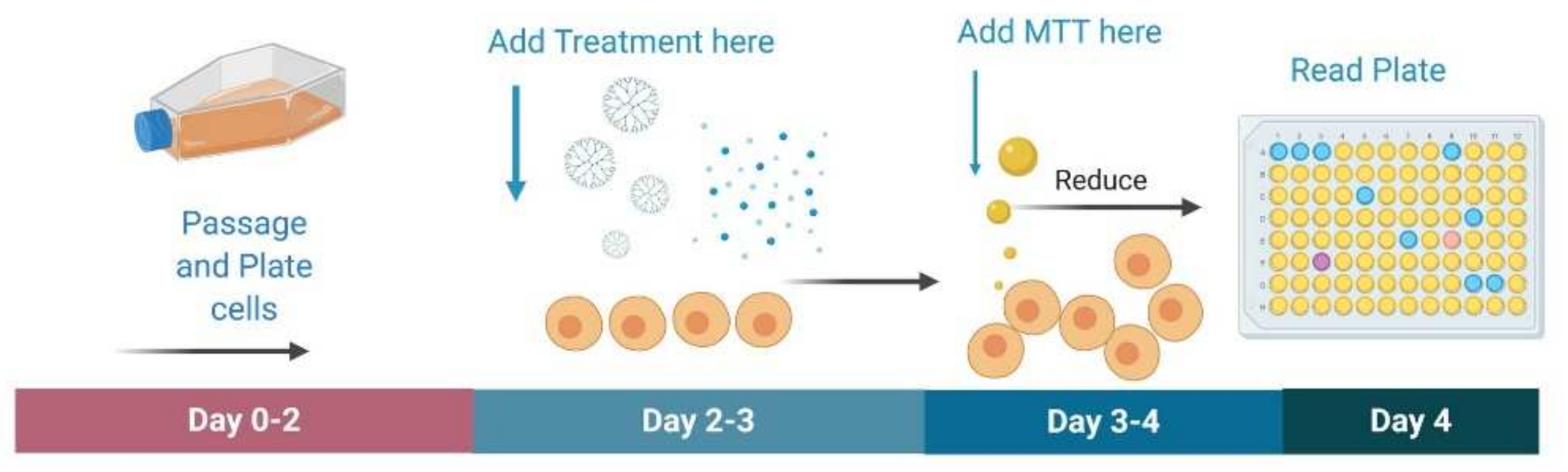
The goal of the study was to develop modified PAMAM dendrimers of different formulations and encapsulating curcumin as potential treatment for glioblastoma in rodent and non-rodent cell lines. Efficacy and specificity of treatment toward cancer cells was assessed using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) toxicity assays. Although qualitative assessments of cell death included observations of floating cells, accumulation of cell fragments and precipitated material, change in media color, etc., the MTT assay was used to quantify the cell death. The MTT is commonly used to determine general toxicity profiles for new pharmacological therapies.
3. Discussion
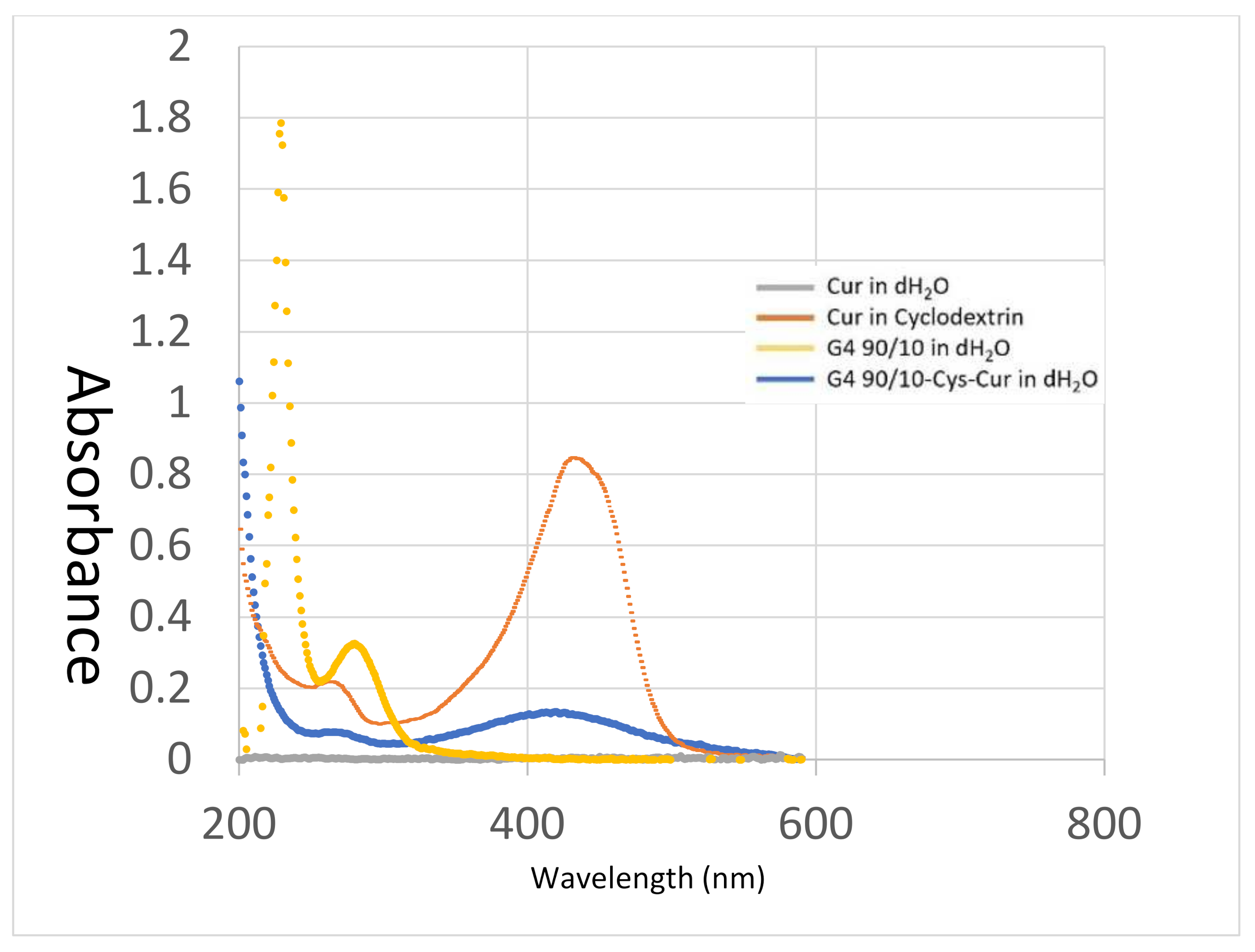
Dendrimer nano-molecules are increasingly used in nanotherapeutic research, particularly for the treatment of cancers, due to their small size and ability to encapsulate and deliver non-soluble treatments deep into the tumor. Over the past few years, our lab has focused on the delivery of the polyphenol curcumin, and has found that, by encapsulating this nutraceutical inside fourth generation PAMAM dendrimers, we were able to encapsulate the curcumin, thereby increasing curcumin bioavailability.
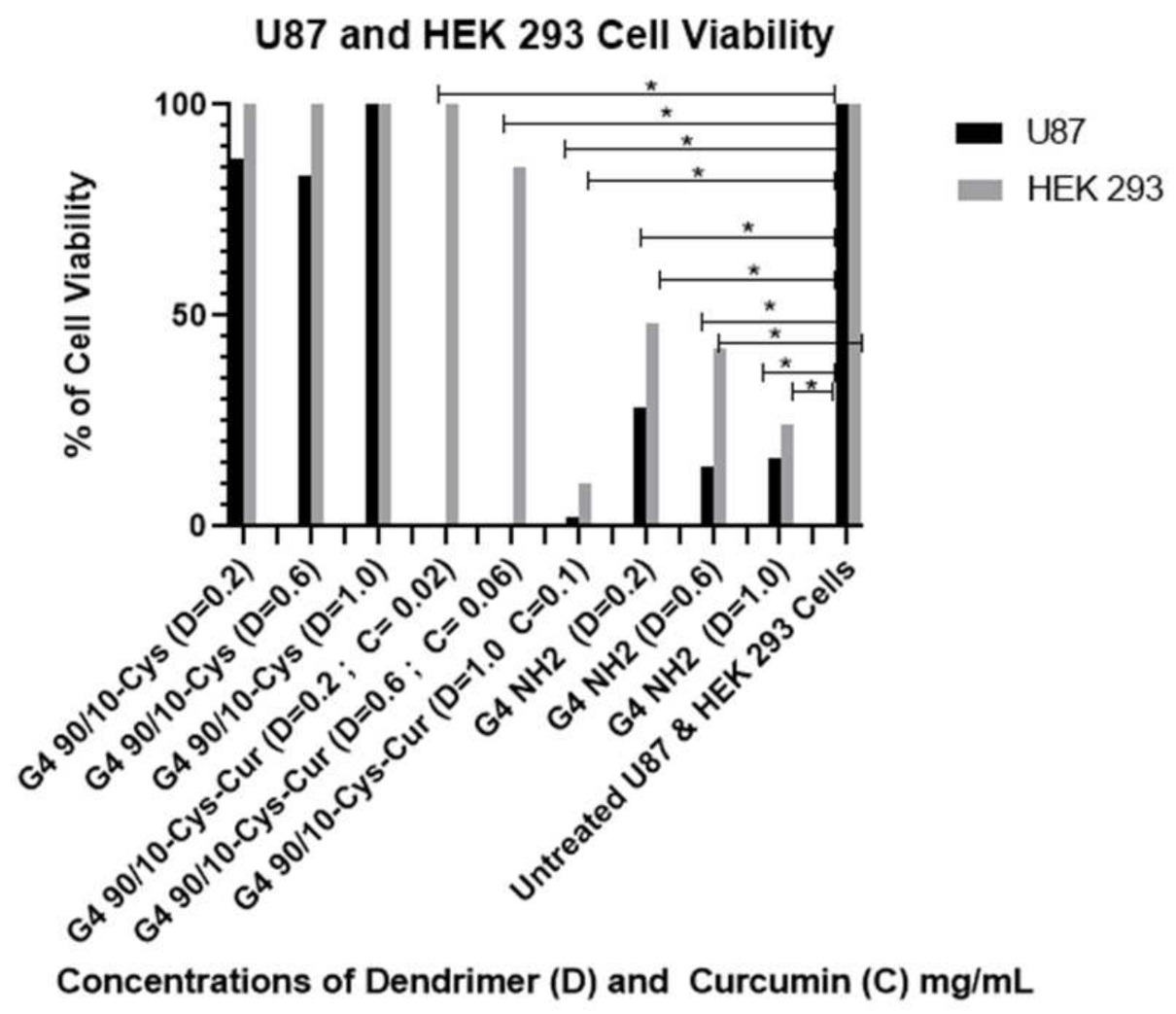
Under our experimental conditions, curcumin by itself was ineffective at killing any of the cancer cell lines (human, rat or mouse). This may be related to its extremely poor solubility in water, thereby decreasing its bioavailability. Encapsulating curcumin into the mixed surface dendrimer (G4 90/10-Cys) showed a remarkable increase in its anti-cancer properties. In the case of the human cancer cell line (U87), even the lowest dose tested (0.02 mg/mL) killed 100% of the cancer cells while showing no effect on the healthy HEK 293 control cells (0% killing). A similar trend was noted with the mid-dose of 0.06 mg/mL encapsulated curcumin. However, at the highest dose (0.1 mg/mL), 90% of healthy HEK 293 cells were also killed. These results suggest that there is an optimum dose range of encapsulated curcumin that would be effective for glioblastoma treatment (
Figure 8).
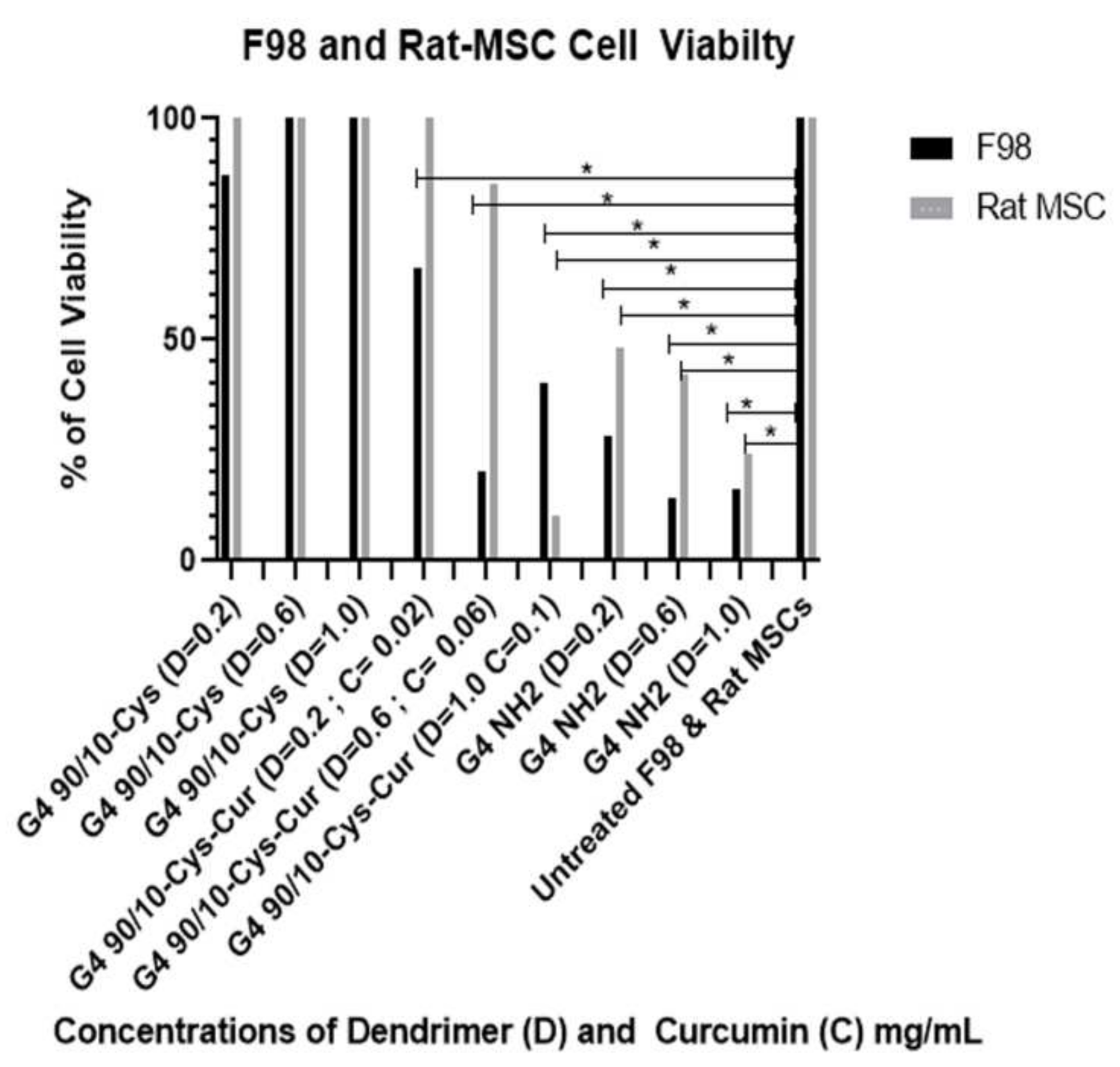
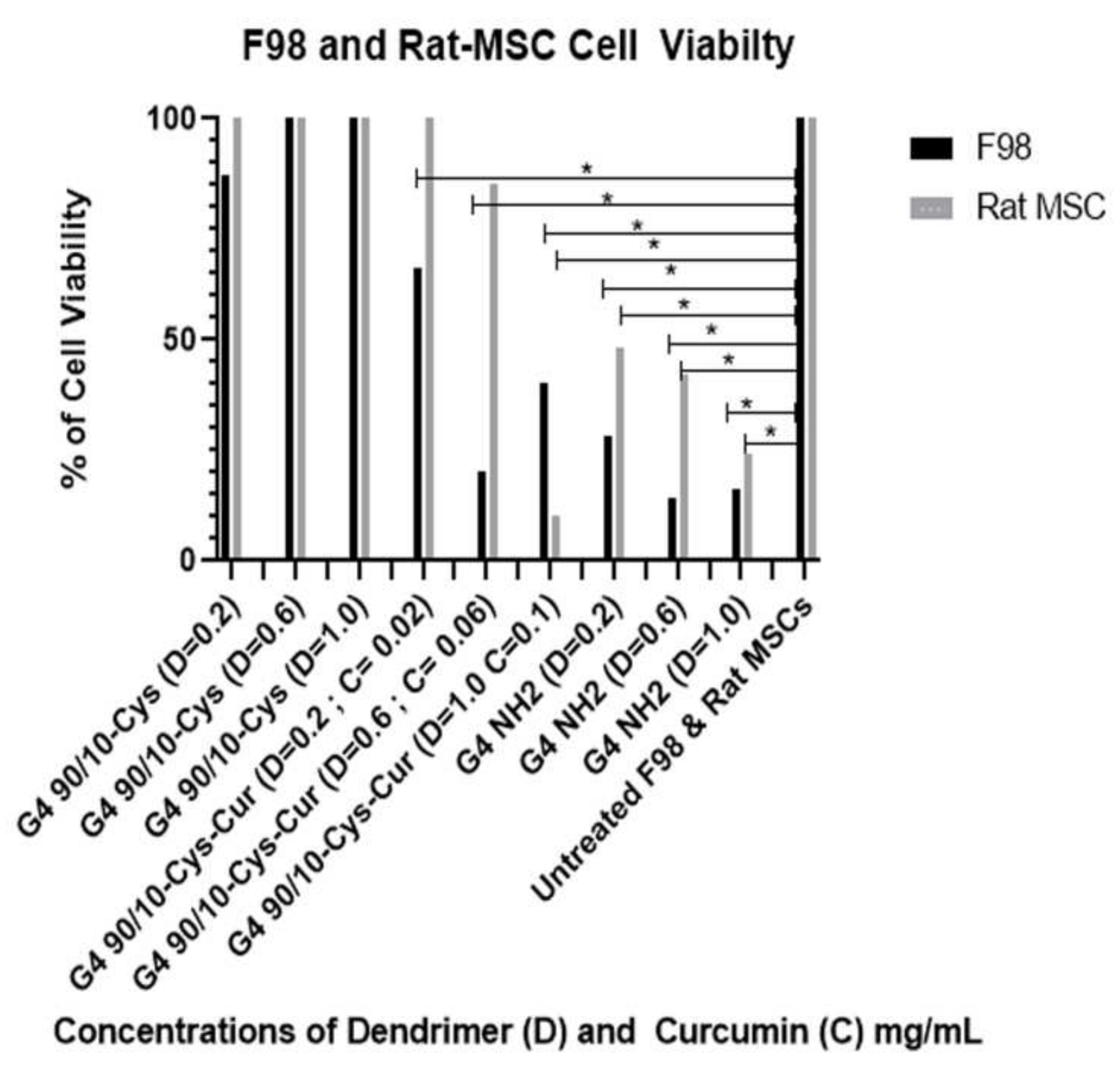
Encapsulated curcumin was less effective in killing mouse (GL 261) or rat (F98) cancer cells. In the case of mouse cells, the lowest dose (0.02 mg/mL) showed about 30% killing while the highest dose (0.1 mg/mL) gave about 80% killing of the cancer cells. None of the doses tested showed any deleterious effects on the control rat or mouse (healthy) cells (
Figure 9 and
Figure 10).
Concordant with previous research, G4 NH2 was found to be extremely toxic, reducing cell viability in all 6 cell types, even at low doses. However, these experiments did support our hypothesis that by reducing the number of surface amines, we reduced dendrimer toxicity to both healthy and cancerous cells. The mixed surface G4 90/10-Cys showed no significant reduction in cell viability in all 3 of the cancer lines, or any of the control lines. We conclude that mixed surface dendrimers are safe and effective candidates for vehicle delivery systems, due to their small (~4 nm) size and ability to encapsulate cargo (curcumin). Additionally, by encapsulating curcumin inside our G4 90/10-Cys we were able to increase the effectiveness of the cargo compared to free curcumin.
When compared to our G4 NH2 toxicity control, we found that our G4 90/10-Cys-Cur was actually more efficient at killing U87 cells than the G4 NH2, at all three doses. While this was not reproduced in the rodent cell lines, it shows that the human glioblastoma cell line, in particular, may be more sensitive to treatment with G4 90/10-Cys-Cur. These results will be used to help guide treatment options and concentration doses for a glioblastoma tumor model in vivo.
Potential new pharmacotherapeutics emerge from biological sources, chemical molecules, vaccines, and other untapped or re-purposed biological sources. Medicines developed from different sources can respond to genetic predisposition, risk factors, and comorbid conditions, producing potential adverse effects in humans, which makes selection of suitable candidates for clinical trials difficult. This difficulty is mitigated with the help of multi-species drug testing studies [
26]. Based on the complexity of using “designer therapies”, it has become a necessary requisite of global regulatory bodies to test new therapeutics in animals prior to human trials (U.S. FDA document:
https://www.fda.gov/media/72028/download) (accessed on 25 May 2021). The multispecies preclinical studies provide us with safety and tolerability data of new drugs to support the initial therapeutic dose and long-term drug dosing calculations for clinical trials. Taken together, our treatments with encapsulated curcumin have reduced cellular viability across 3 species while leaving control cells relatively healthy at all but one of those same doses. Additionally, our G4 90/10-Cys and G4 90/10-Cys-Cur have proven to be safe when tested on control tissue at lower doses and, therefore, could be used as an adjunctive treatment or potential vehicle in the treatment of GB.
Temozolomide is the current gold standard of treatment for GB. However, in nearly 50% of patients this drug is ineffective, largely due to a natural drug resistance resulting from overexpression of the MGMT gene. Recent data suggest that curcumin, in addition to its many anticancer effects, may function to prevent drug resistance in certain types of cancer [
23]. Other labs have already shown promising results when combining curcumin containing treatments with Temozolomide [
27,
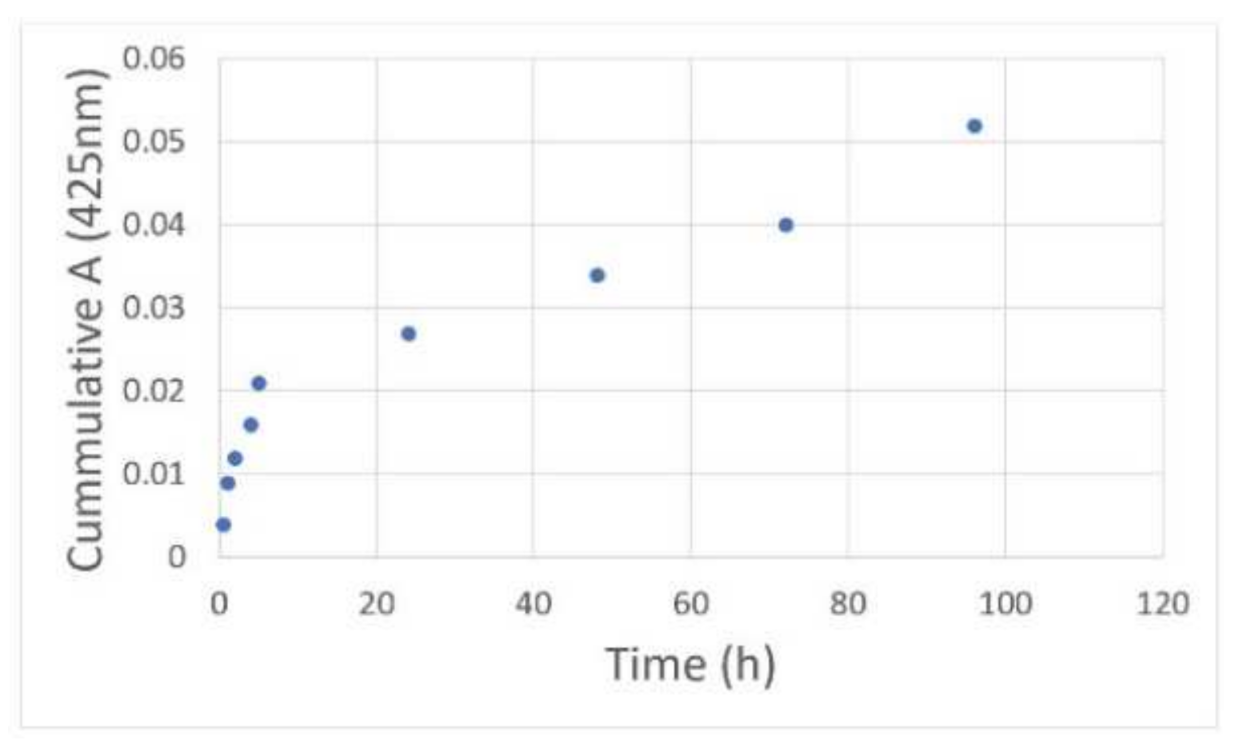
28]. Treatments with just curcumin have largely failed in clinical trials due to its poor solubility, quick degradation, and elimination from the body before it can deliver its full therapeutic effect. By encapsulating curcumin inside a dendrimer nano-molecule, we increased its therapeutic efficacy by improving bioavailability, its release kinetics over a period of time, reducing the rate at which it is released and broken down and eliminated from the body.
Our findings are limited due to the lack of defined mechanisms underlying the MTT data. Curcumin works on multiple cell cycle pathways and tumor suppressor proteins [
29]. It appears that the curcumin is responsible for the reduction in cellular viability of cancer cells, and not the increase in amines on the surface. More investigations are needed to determine if the importance of the curcumin concentration encapsulated in our dendrimers plays a role in the cell viability. However, delineating which specific genes are responsible for these effects is important in tailoring the therapy to individuals or for combined adjunctive treatment. Future
in vitro studies in this lab will include testing hybrid treatments with G4 90/10-Cys-Cur in addition to a chemotherapeutic, such as Temozolomide, to assess their effects on cancerous and healthy tissue cell lines. Based on the results of the
in vitro work, future
in vivo studies will include delivering these treatments into rodent models of glioblastoma to assess their potential where other treatments have failed.


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}