
Discovery of Active Ingredients Targeted TREM2 by SPR Biosensor-UPLC/MS Recognition System, and Investigating the Mechanism of Anti-Neuroinflammatory Activity on the Lignin-Amides from Datura metel Seeds

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
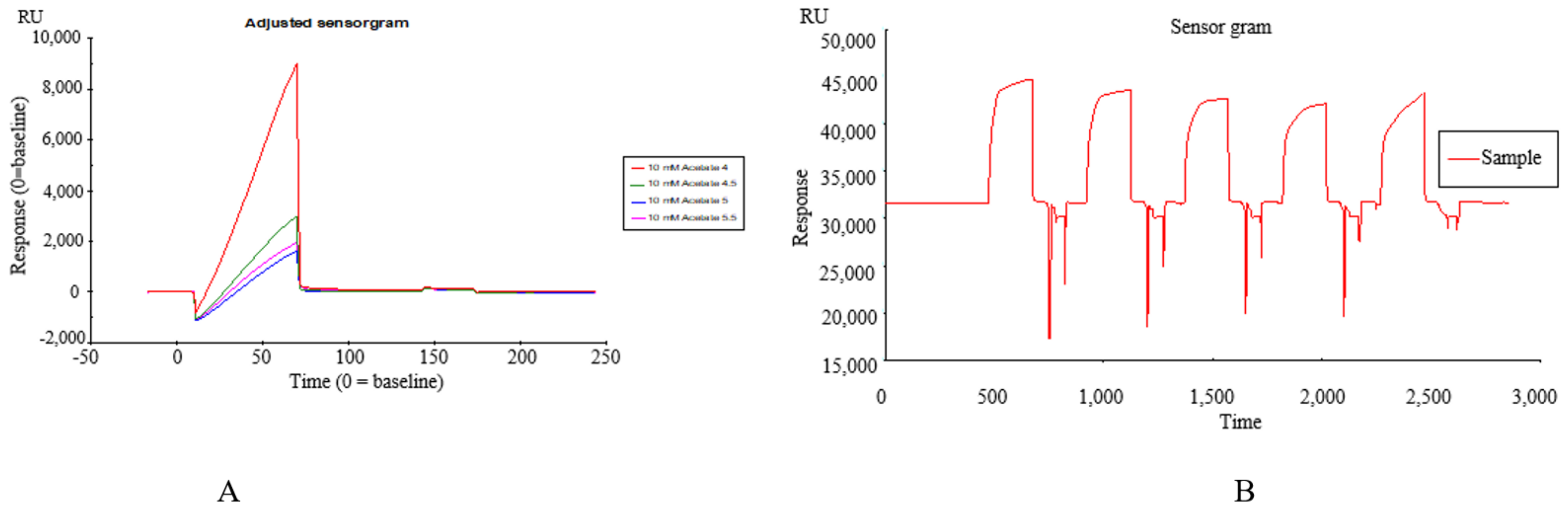
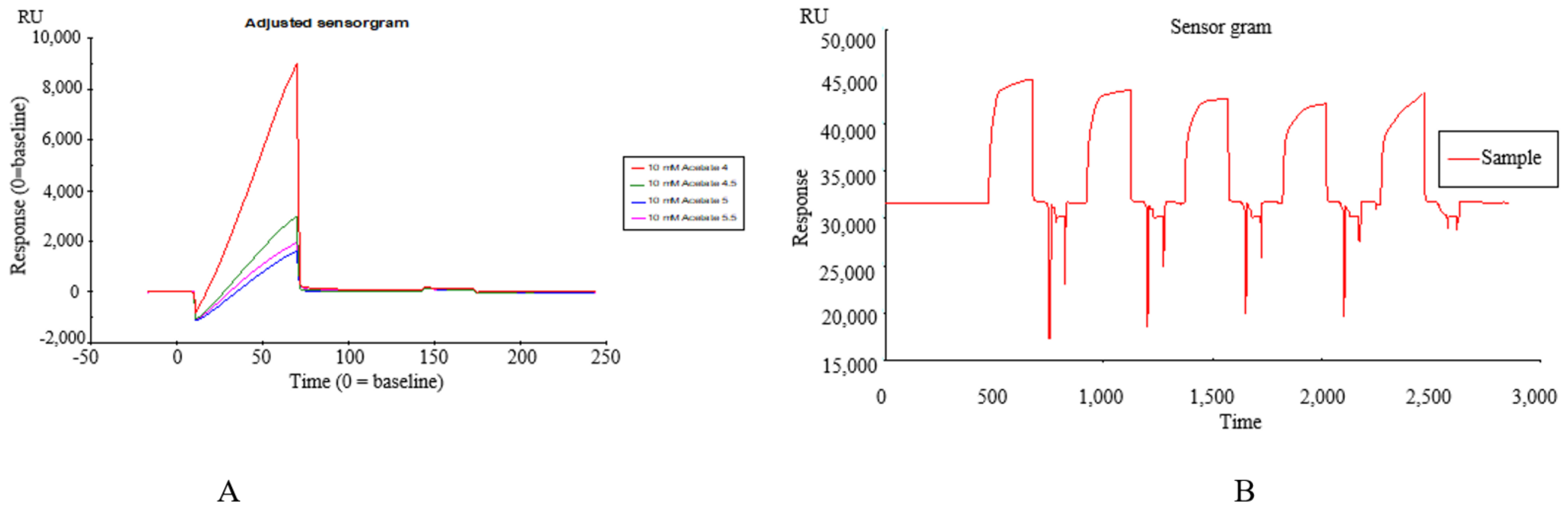
2.1. Construction of SPR Biosensor-UPLC/MS Active Ingredient Recognition System for TREM2 Target
Immobilization of TREM2 on SPR Active Ingredients Recognition System
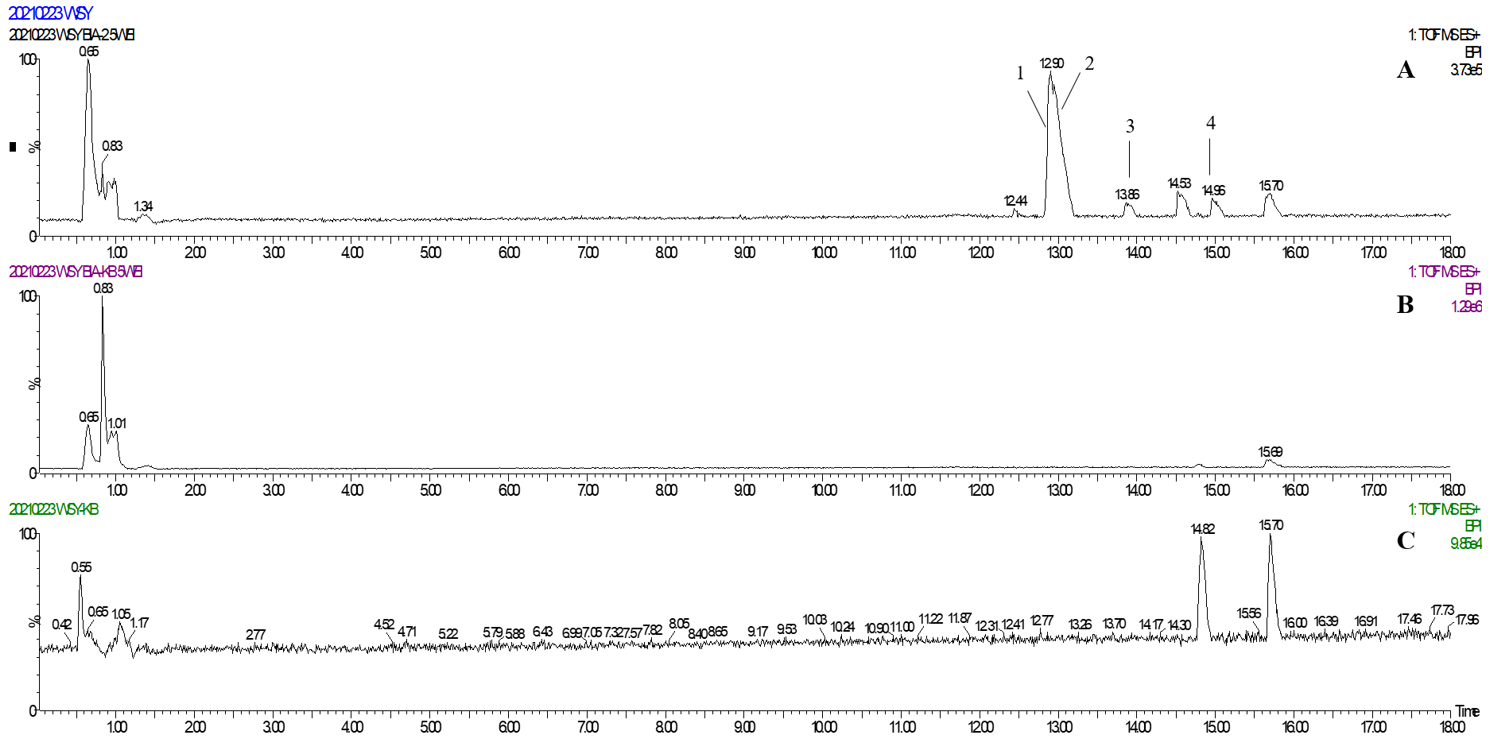
2.2. Identification of TREM2 Target Direct Binding Active Ingredients
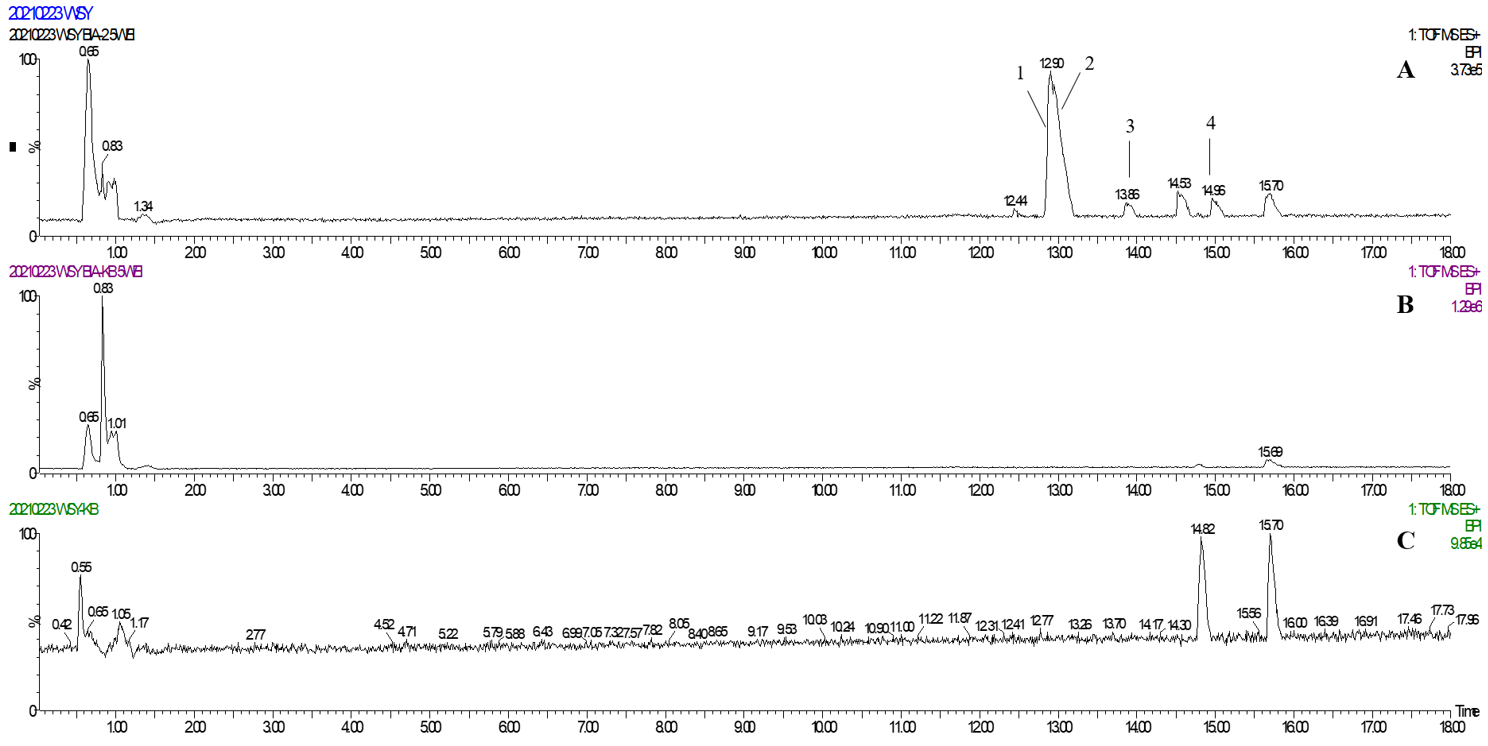
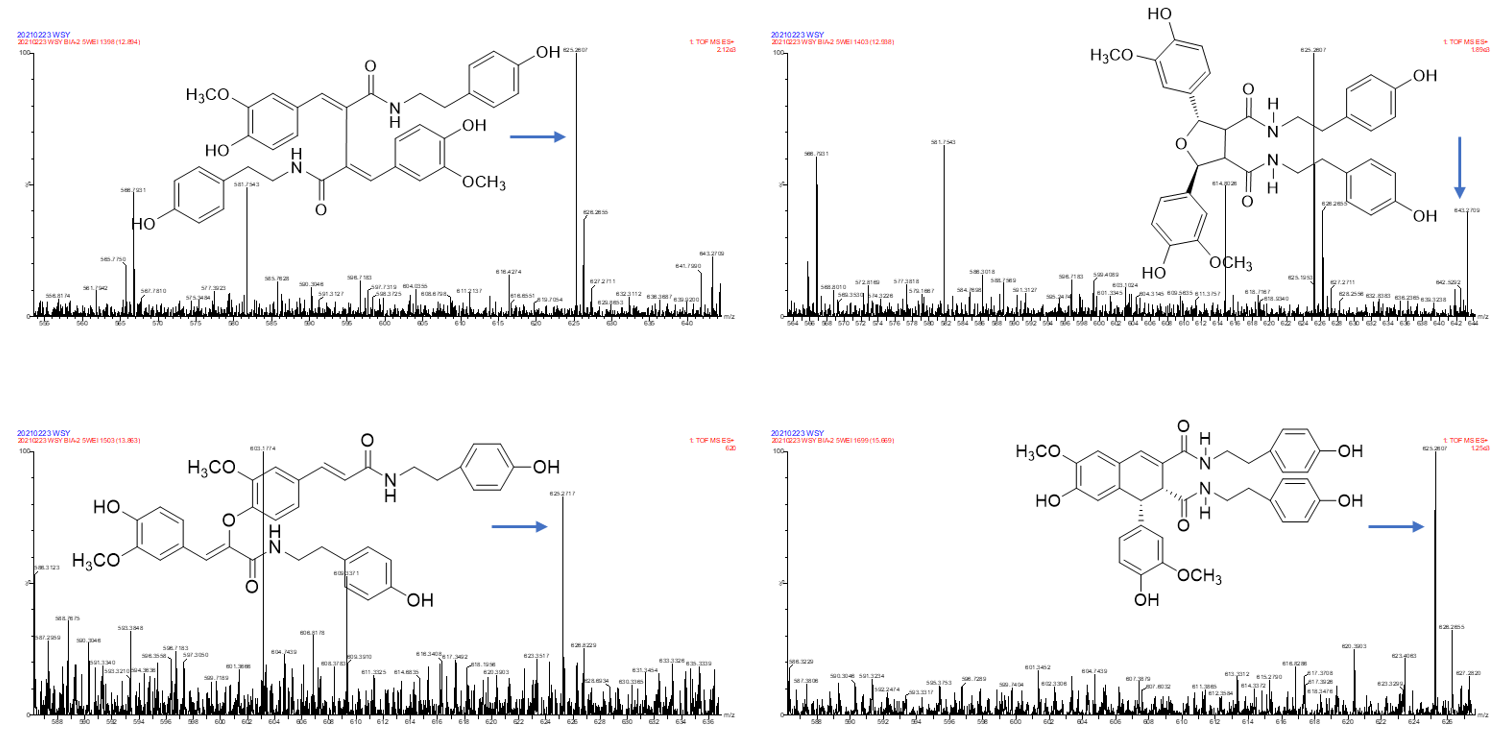
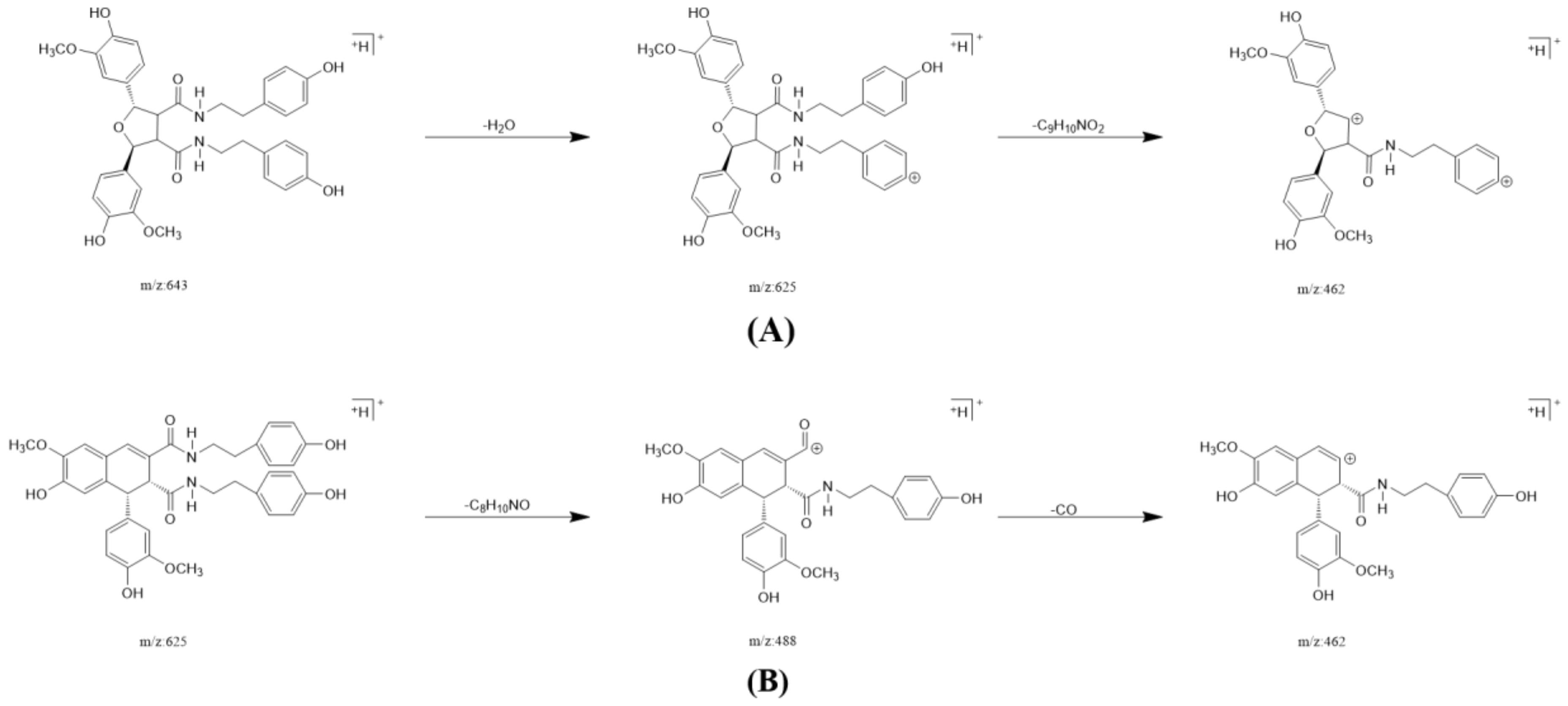
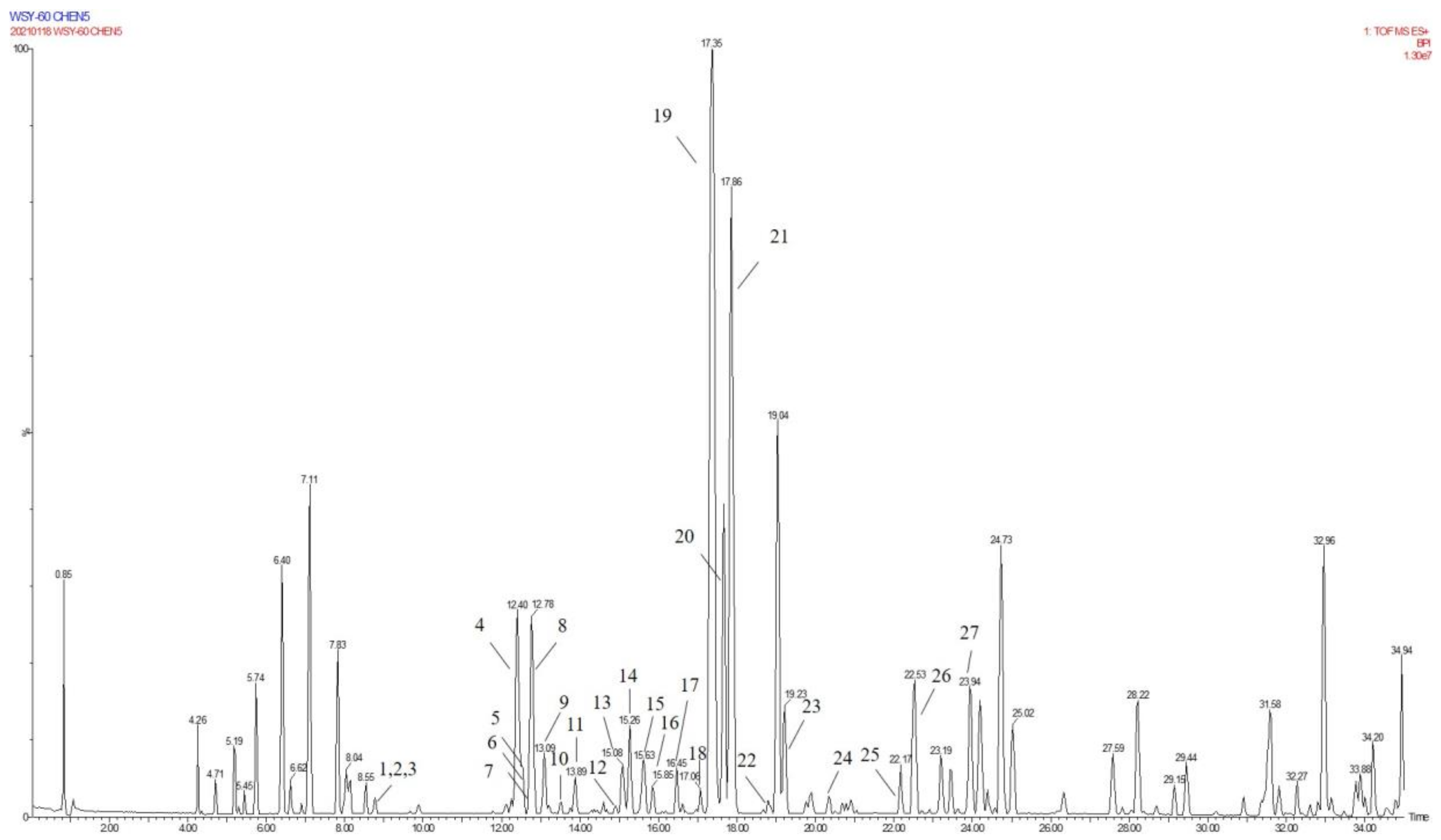
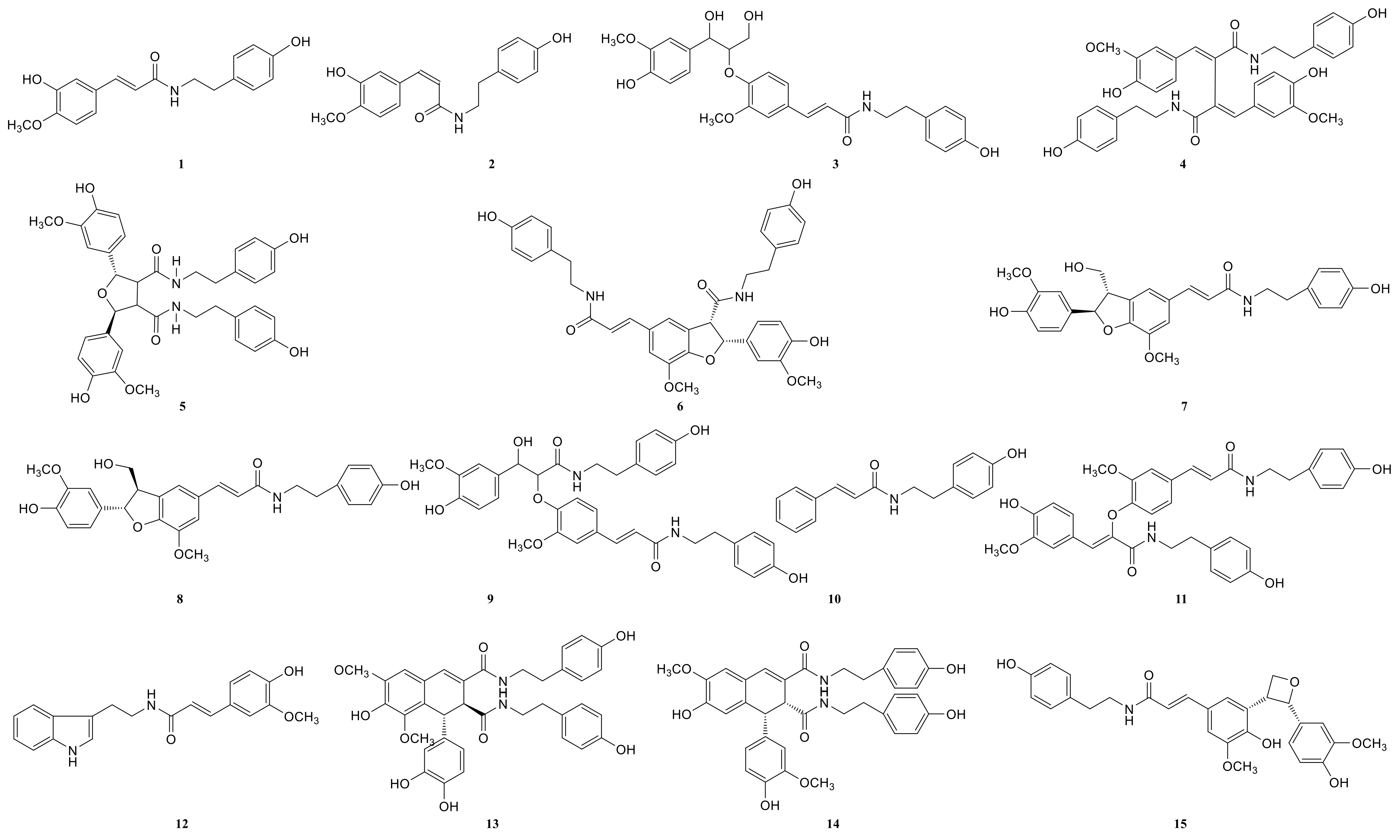
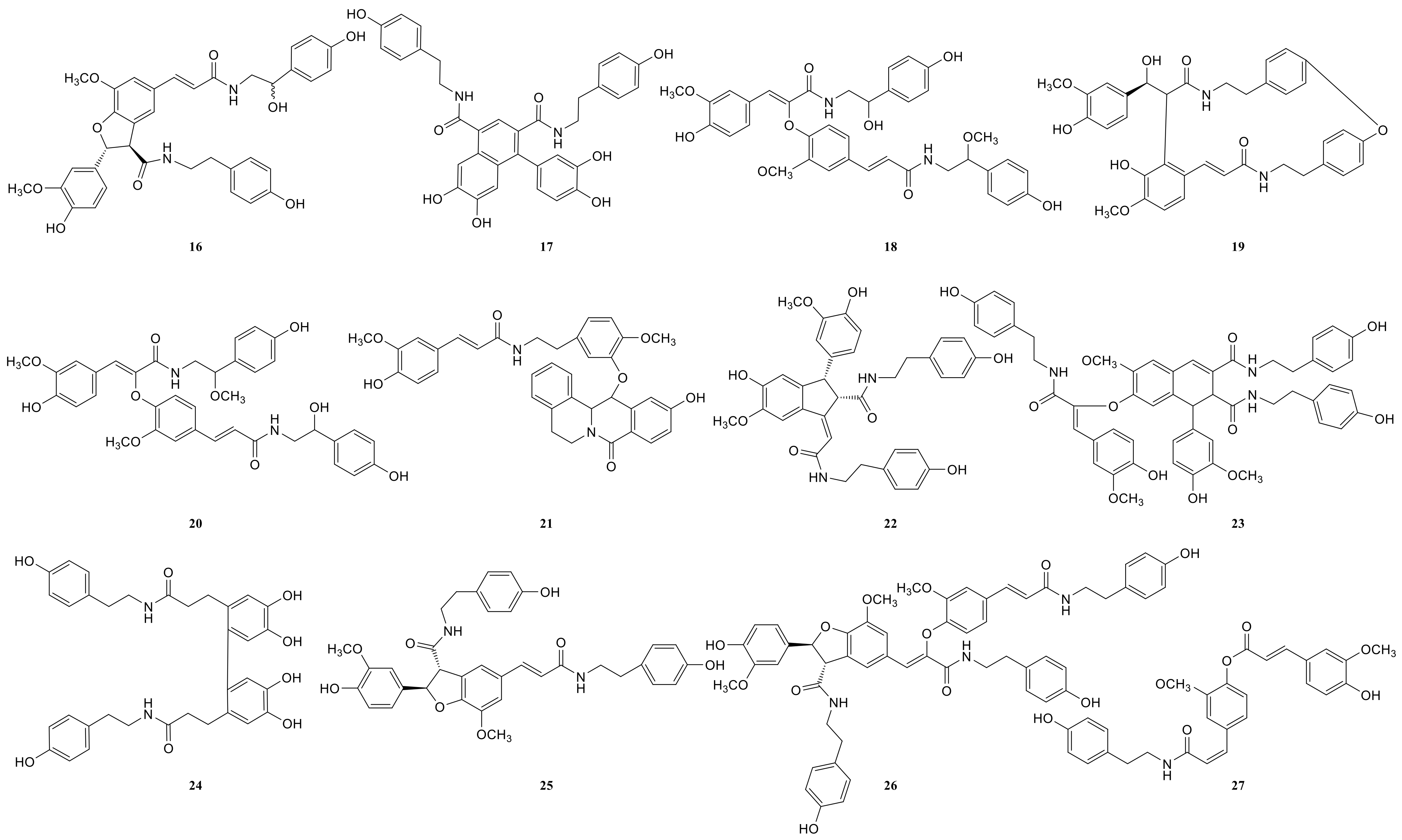
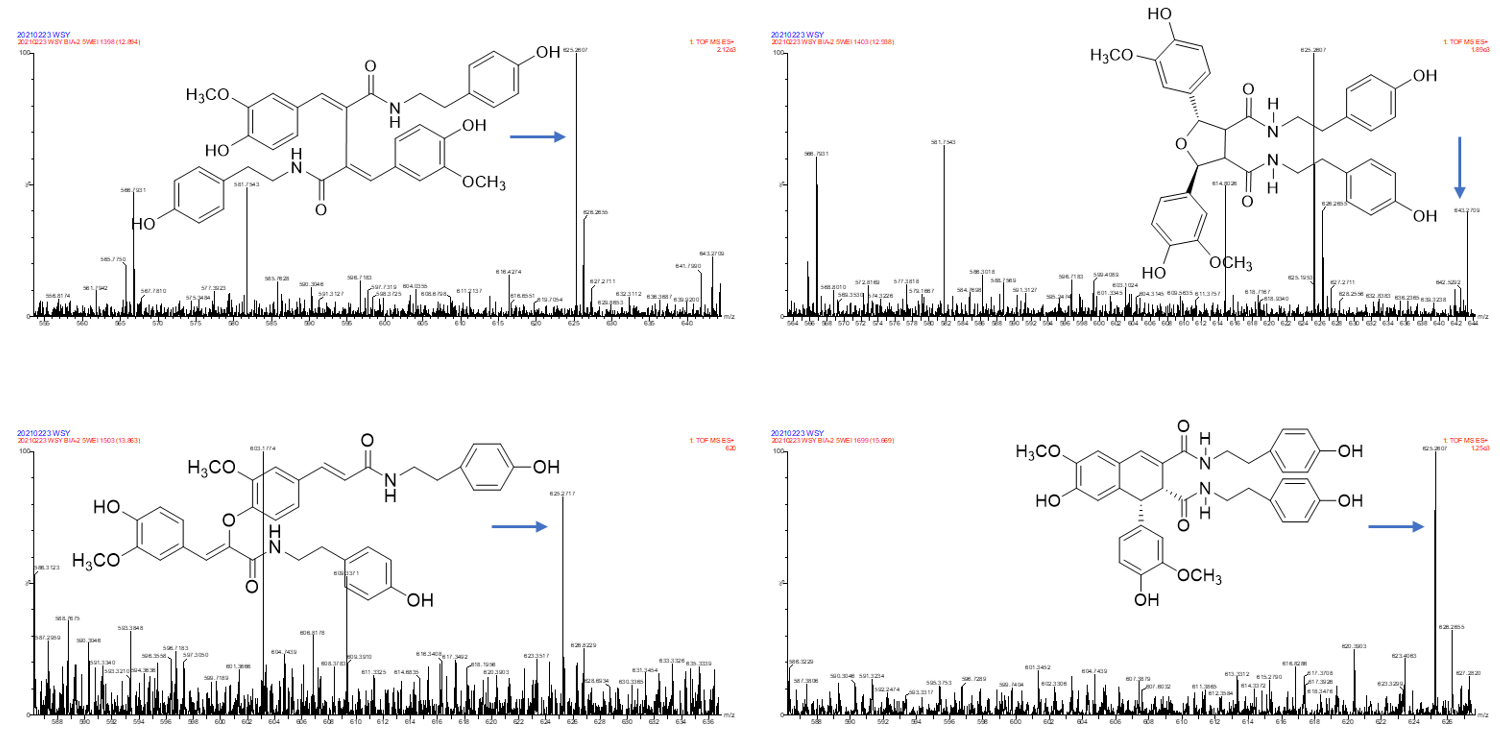
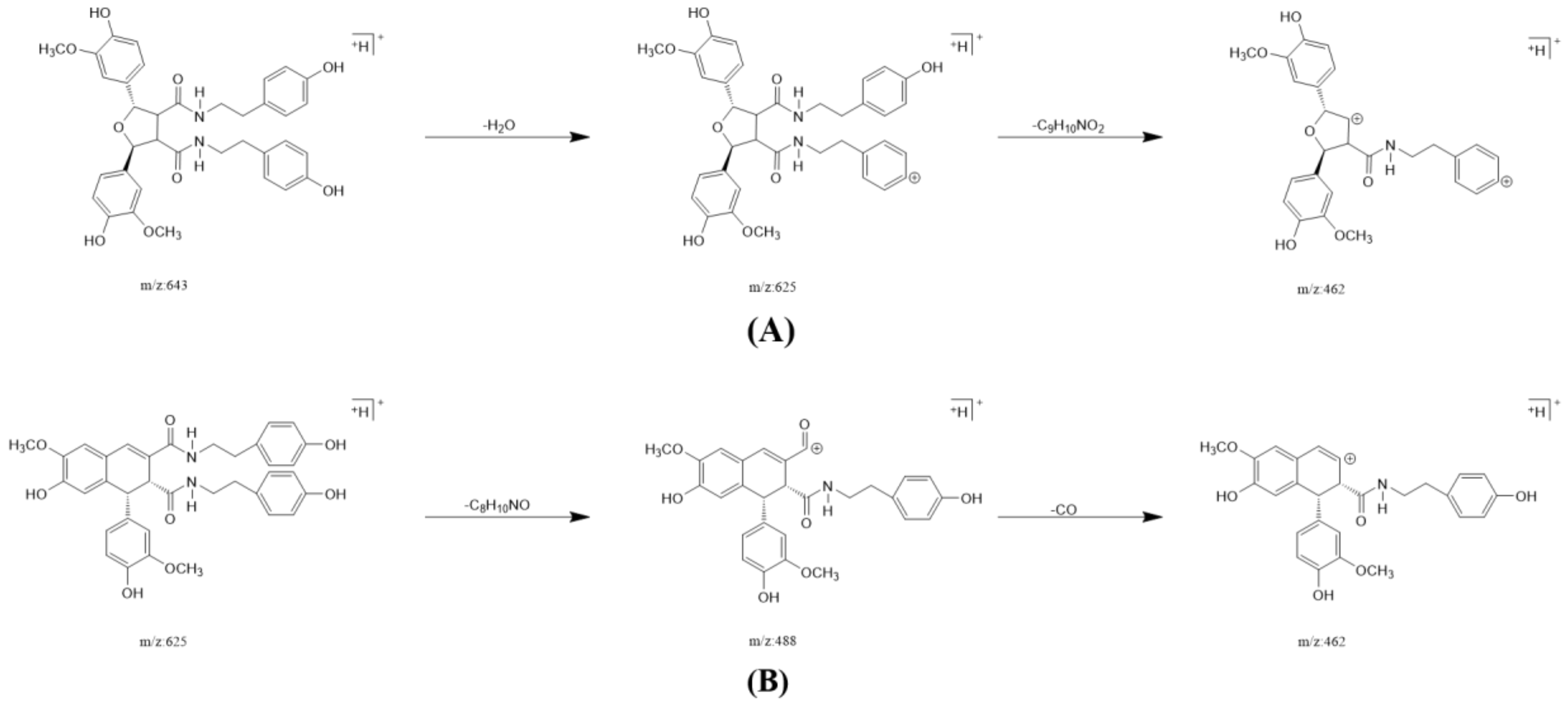
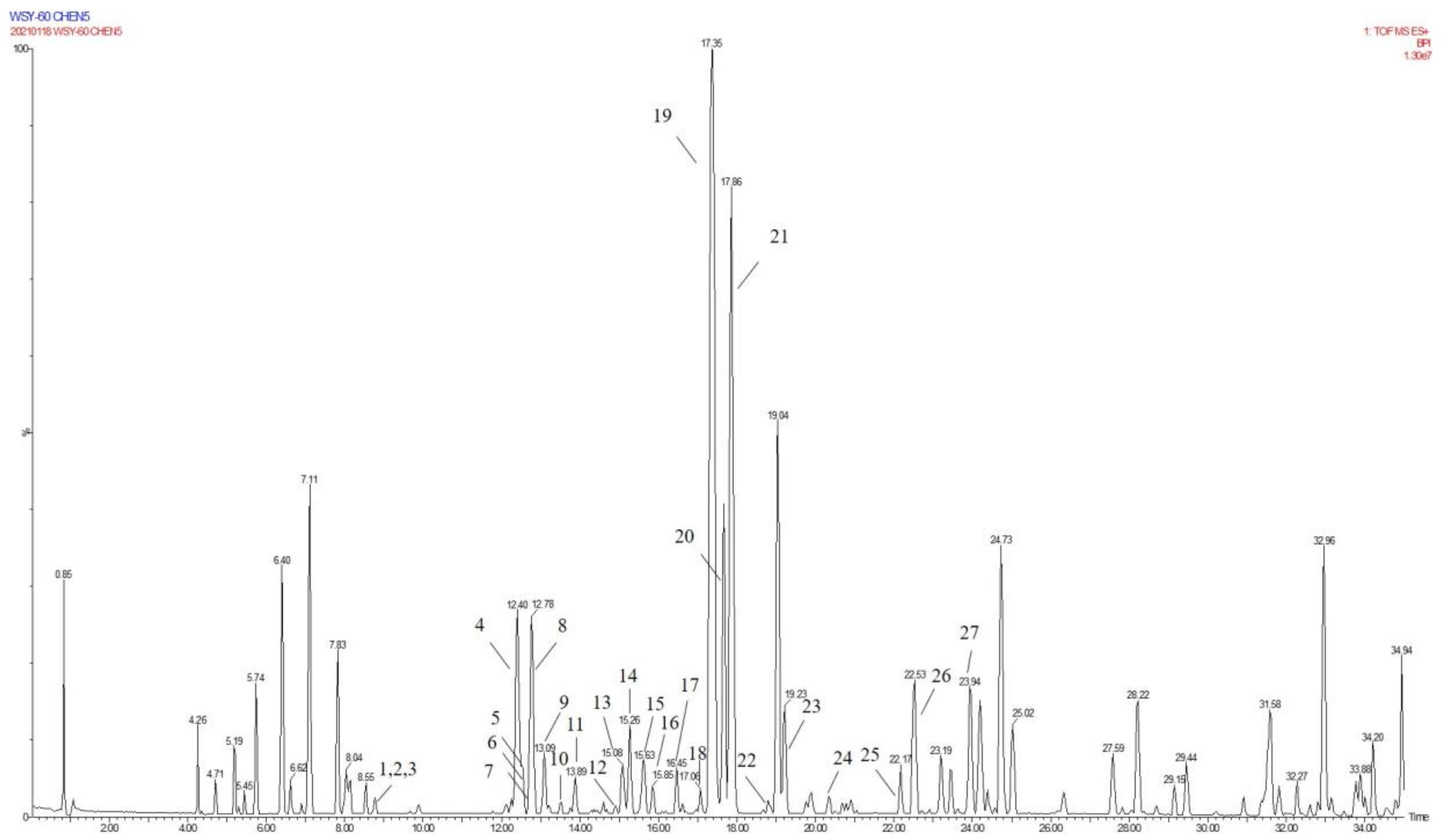
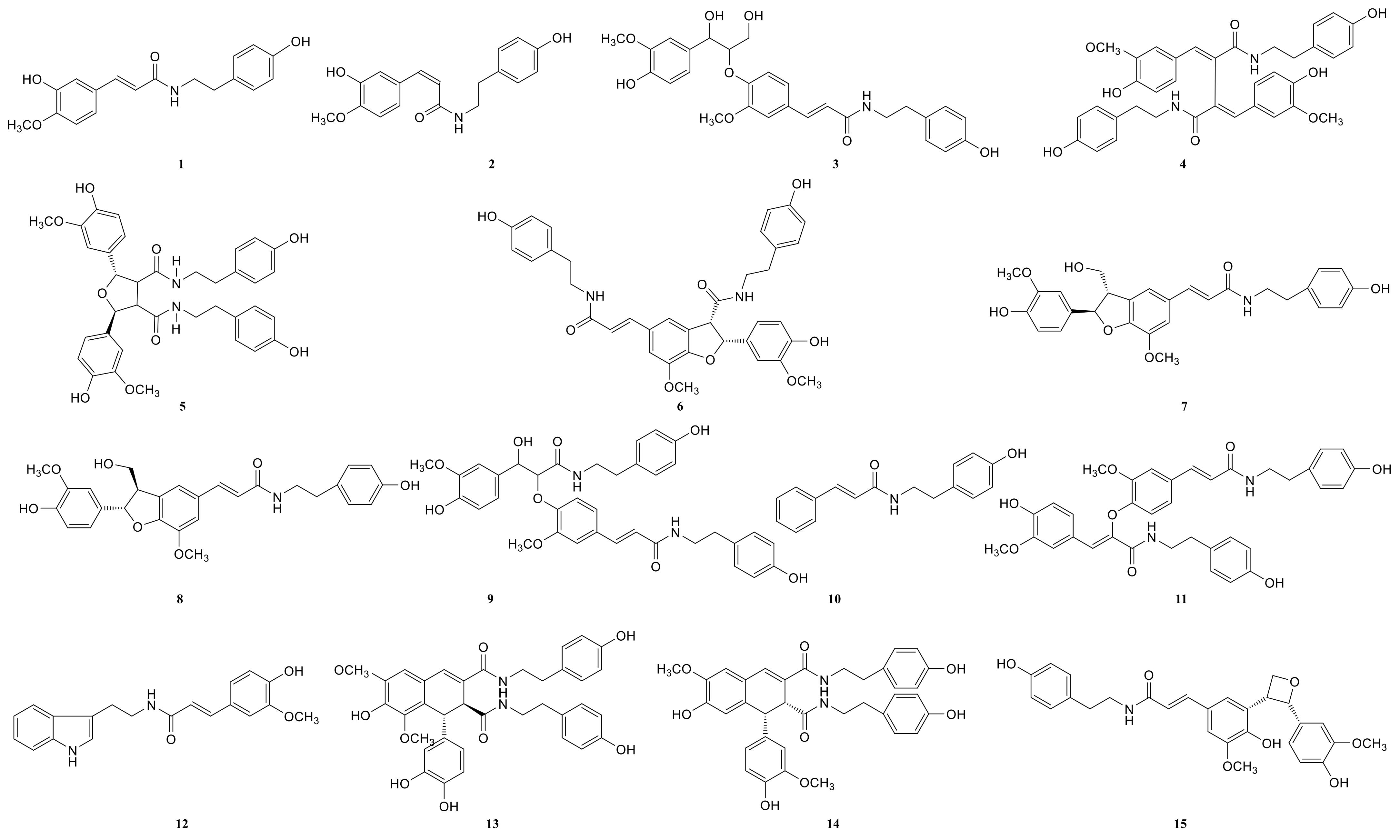
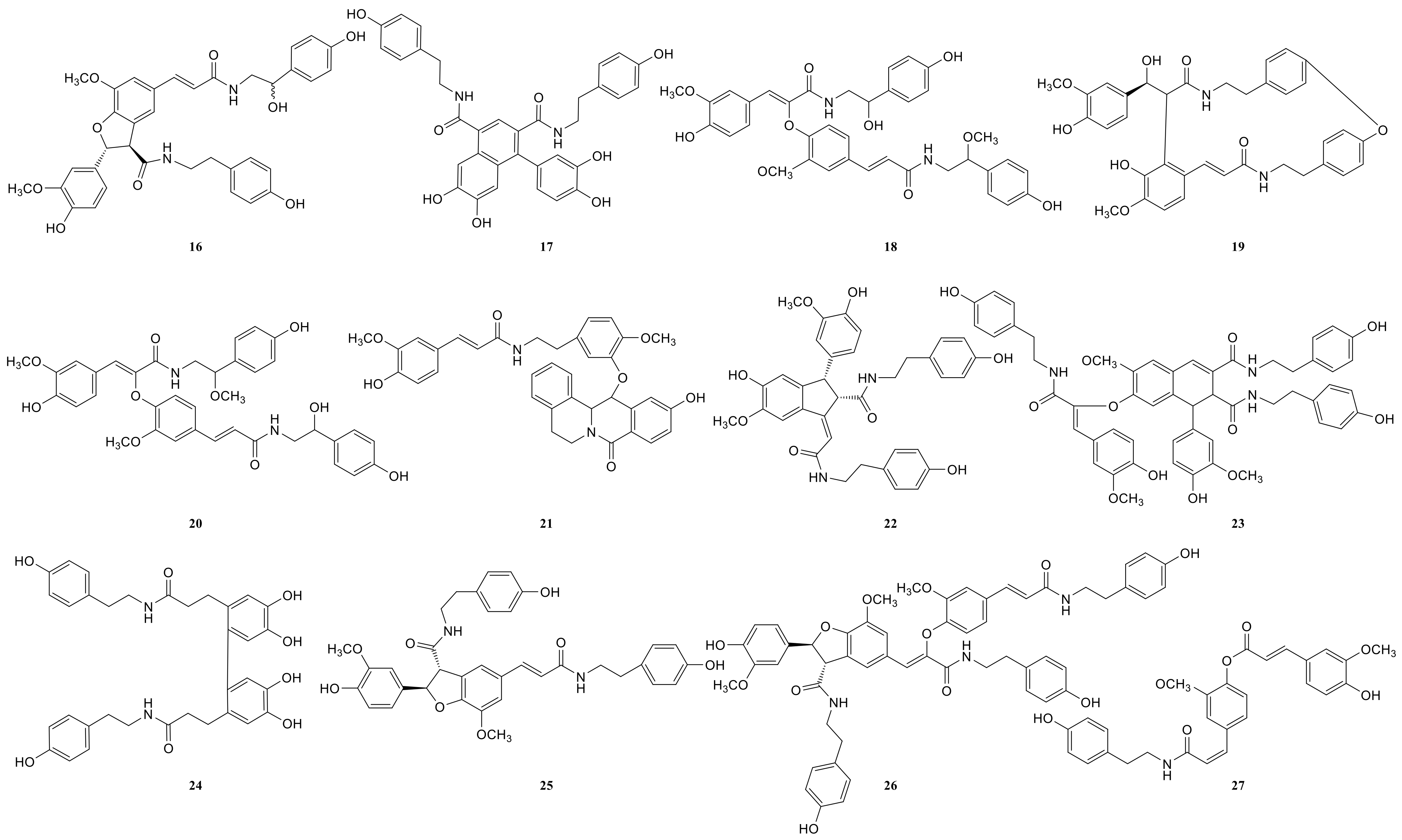
2.3. Annotation of the Enriched Ingredients LDS by UPLC-QTOF/MS Technique
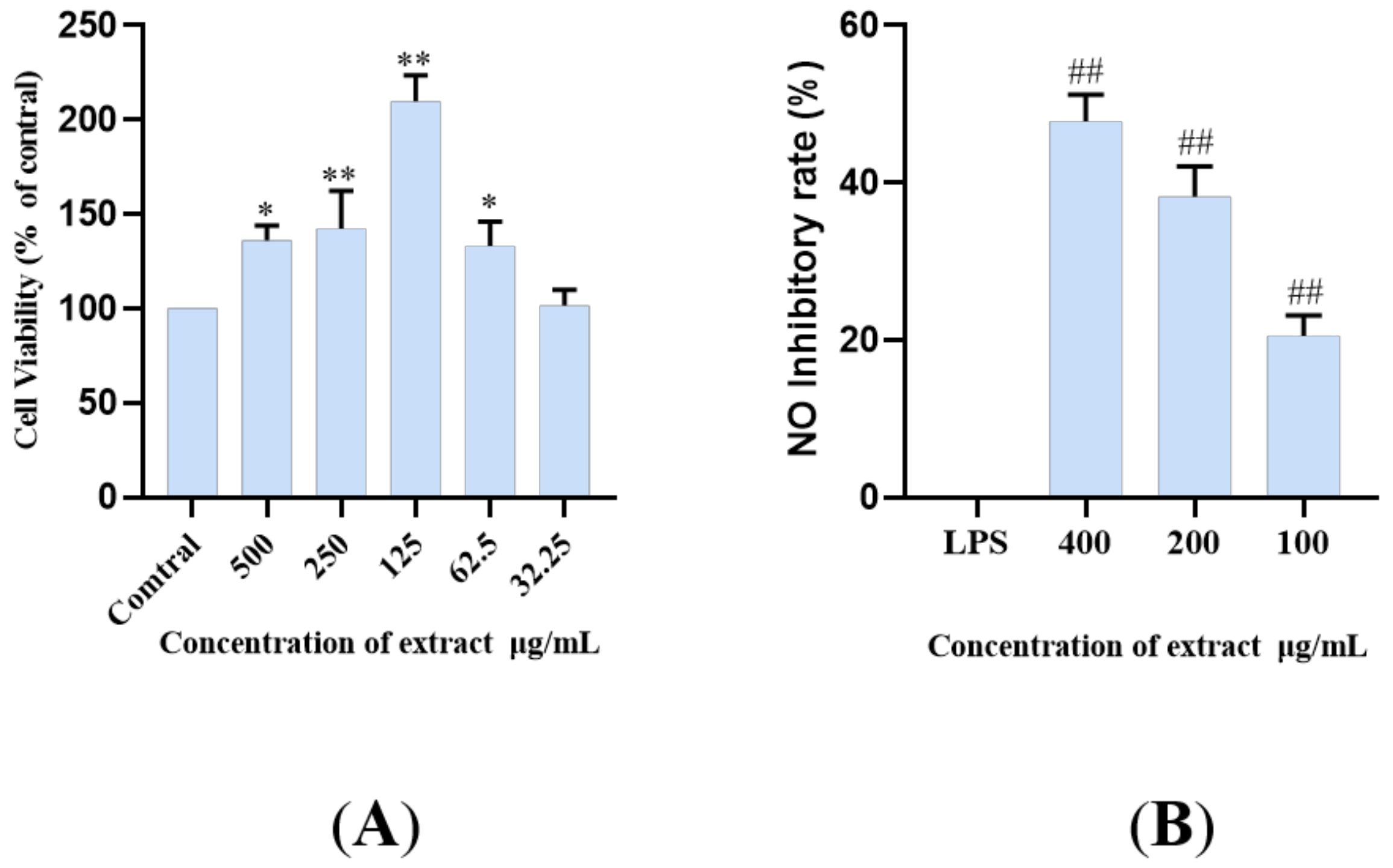
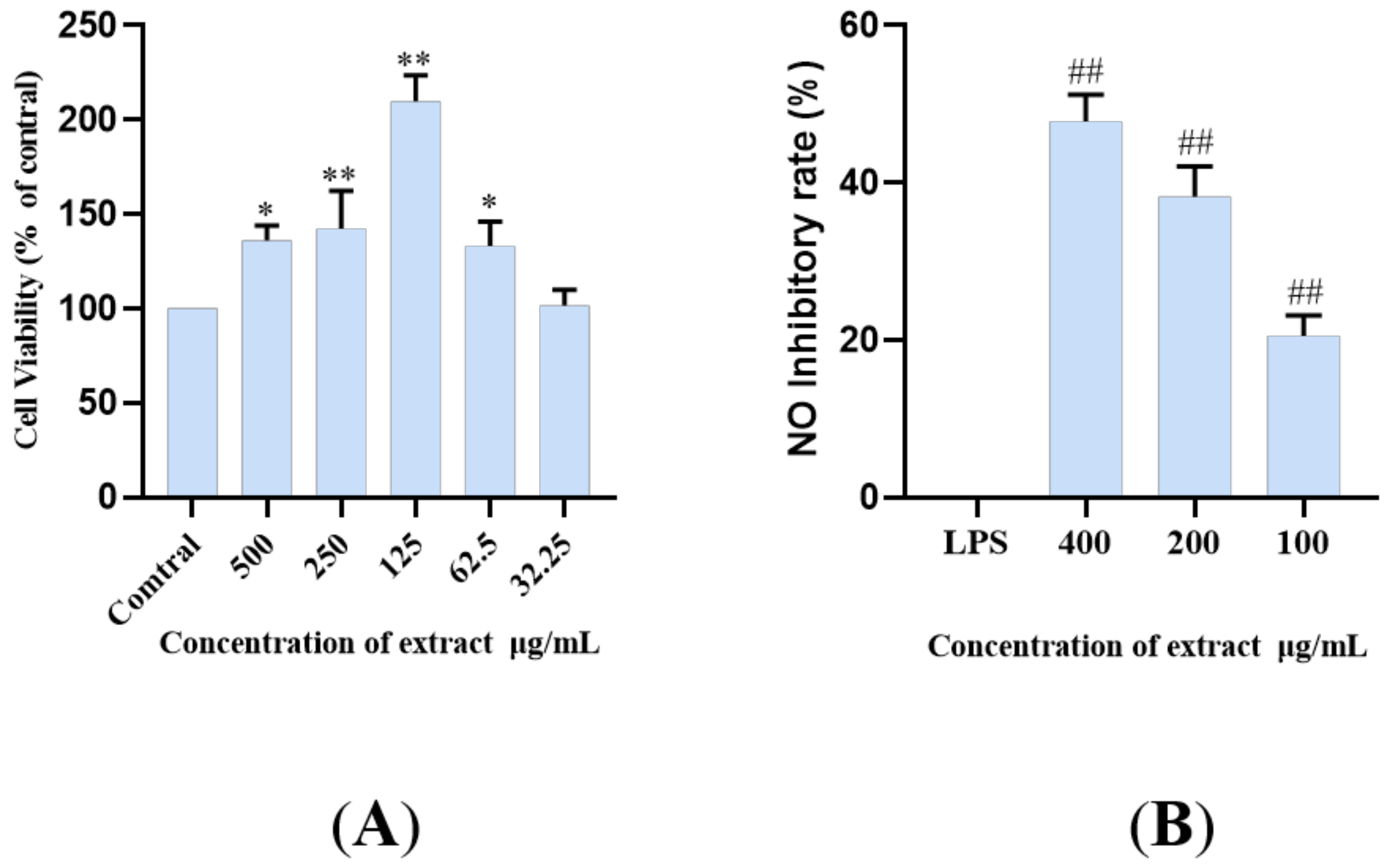
2.4. The LDS Inhibited LPS-Induced NO Release in BV2 Cells
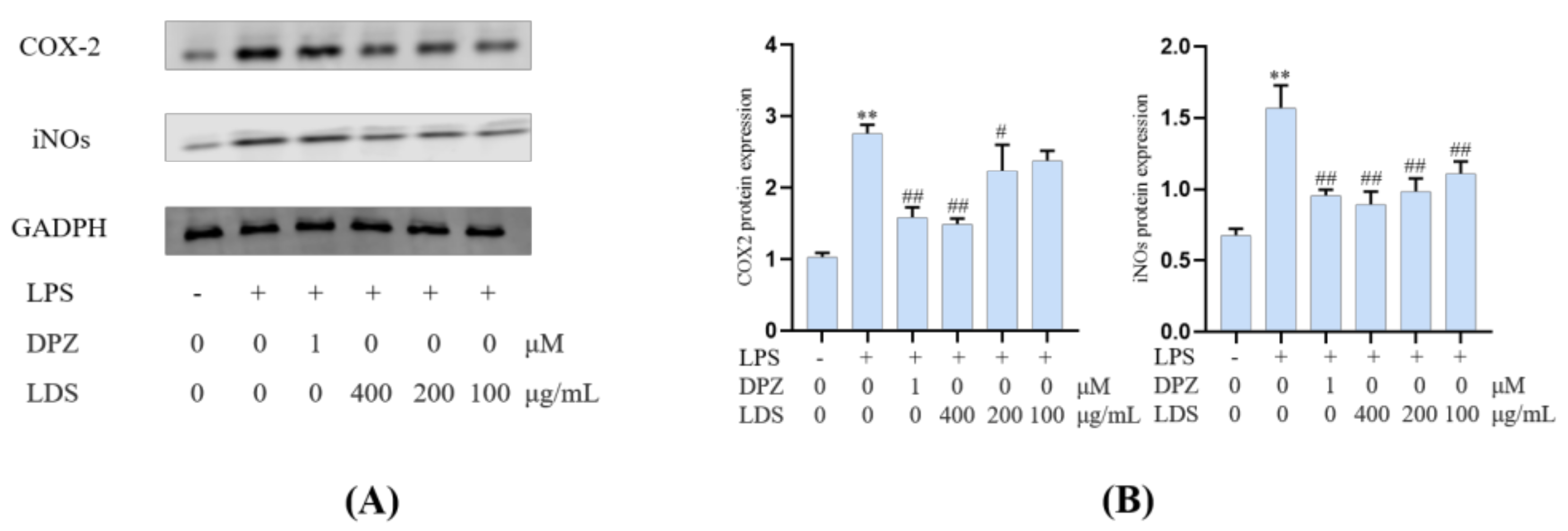
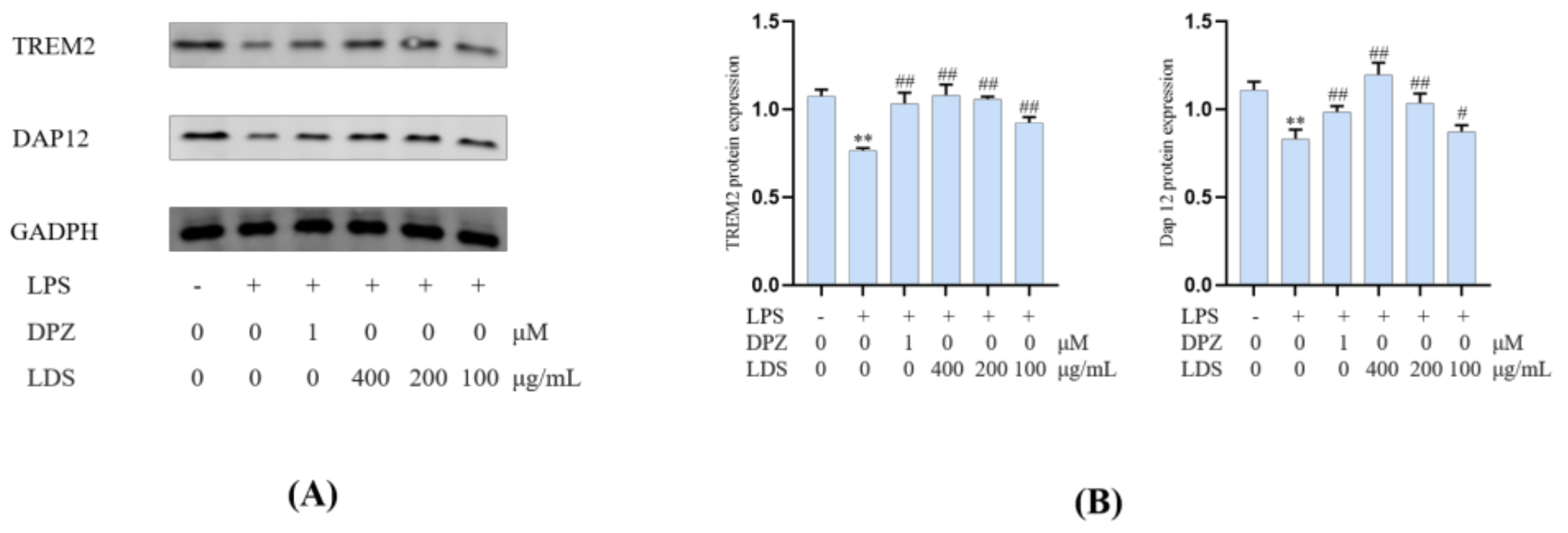
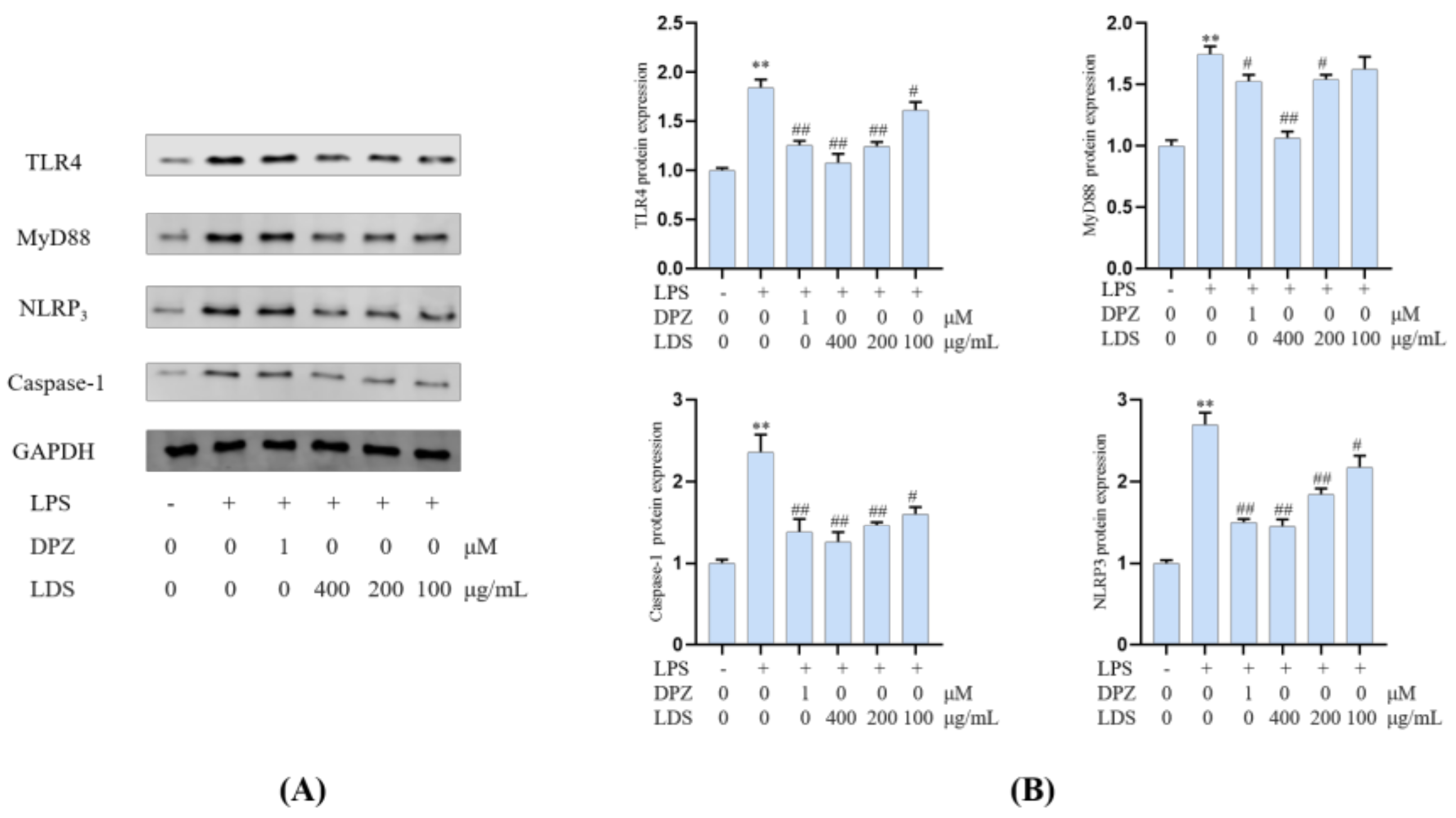
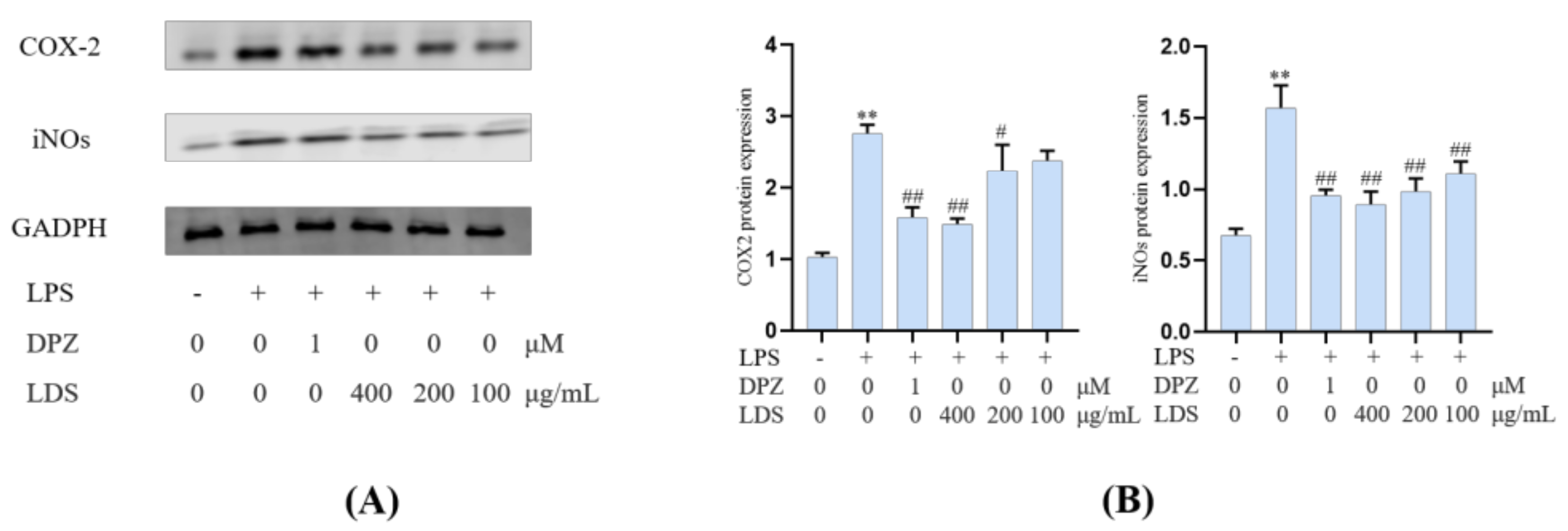
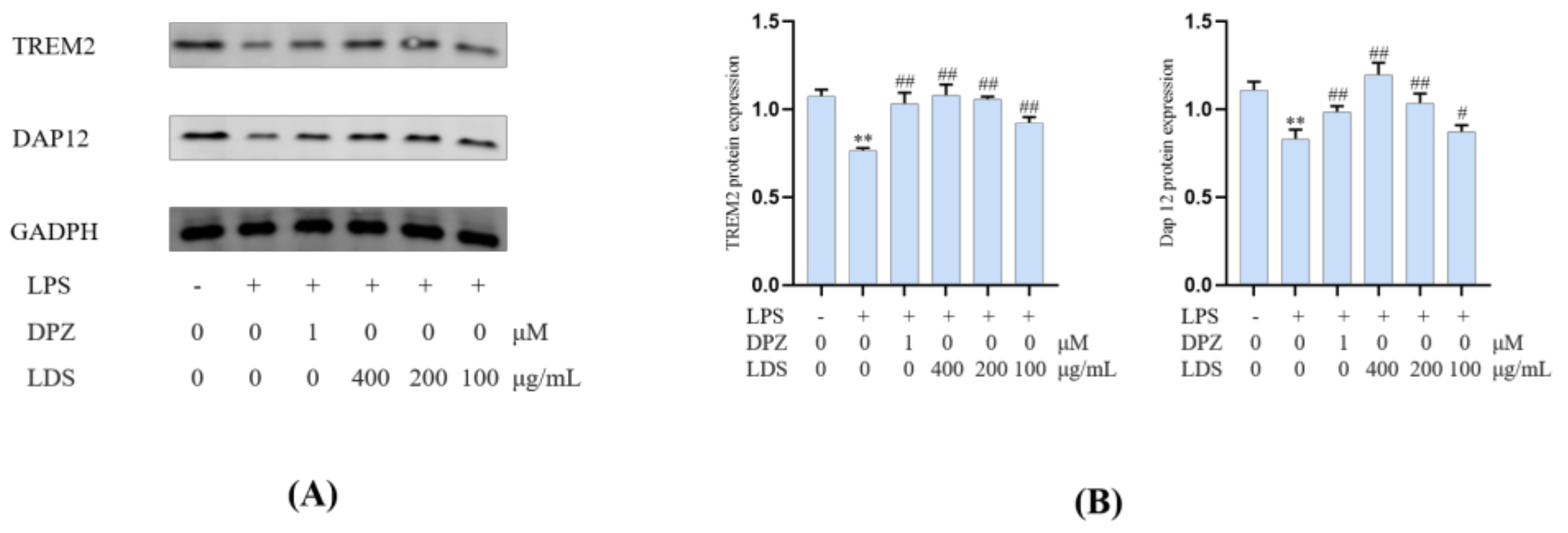
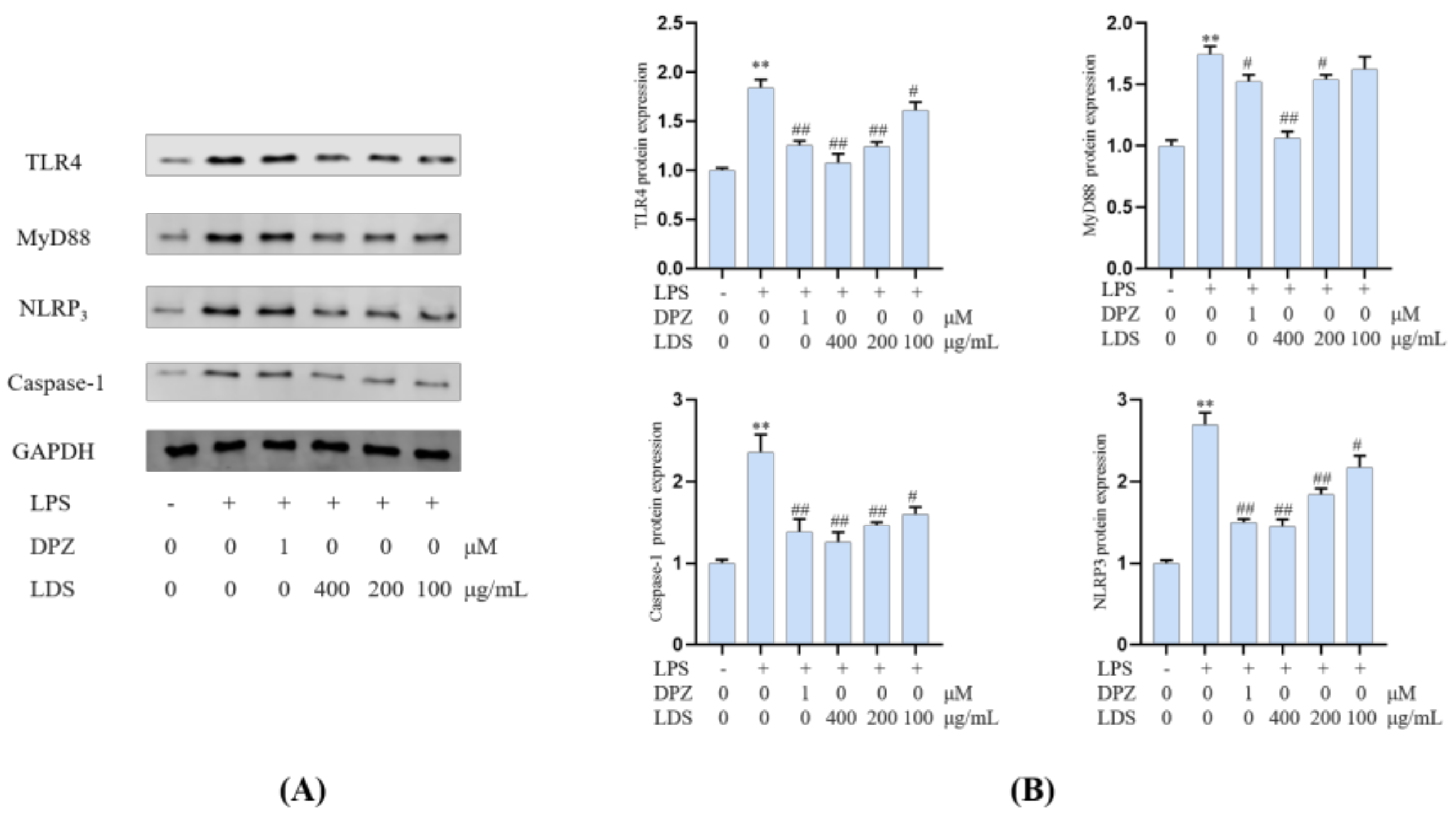
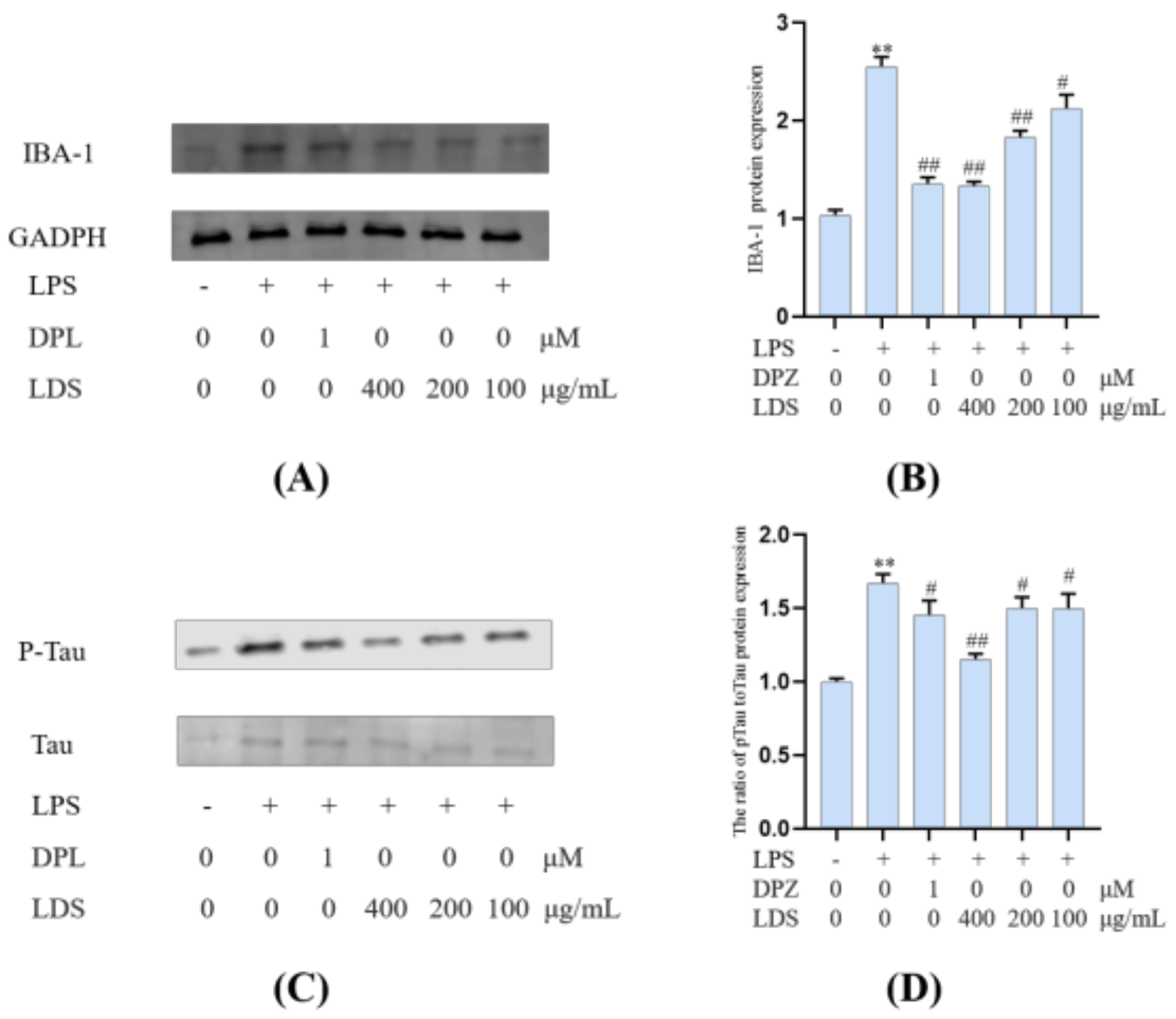
2.5. Protein Expression in BV2 Microglia Cells
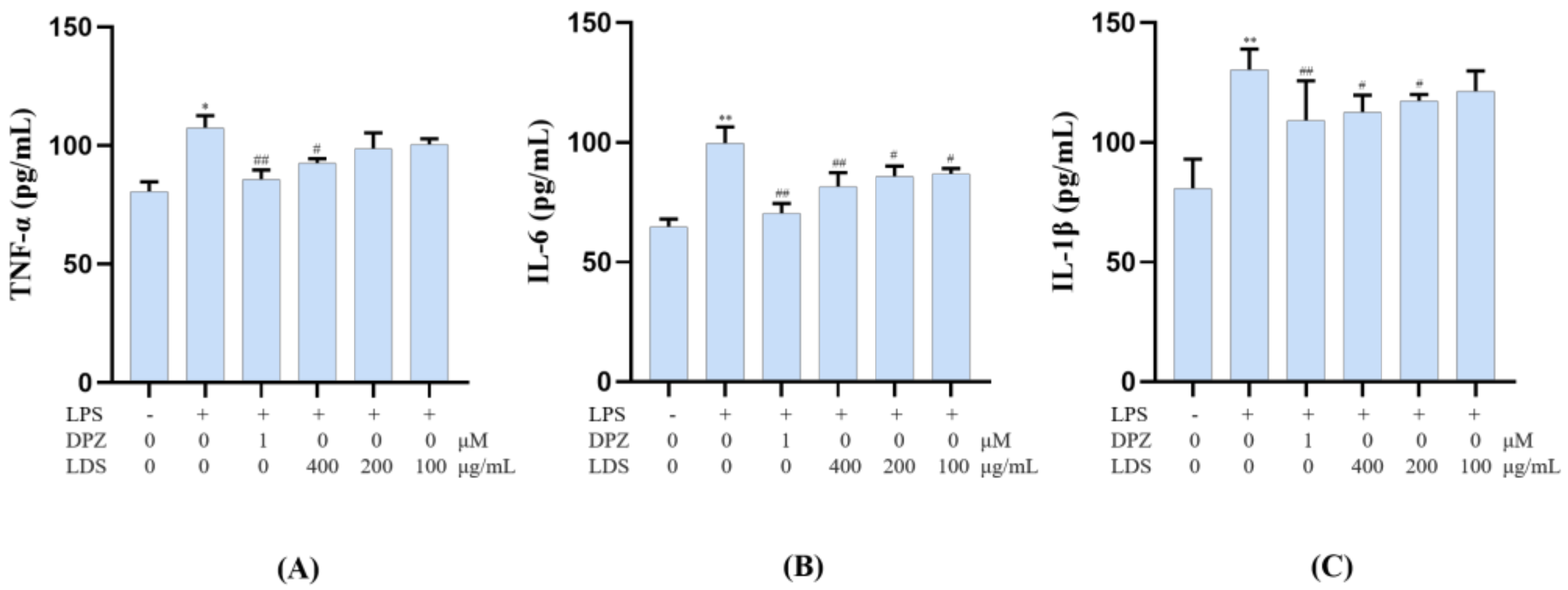
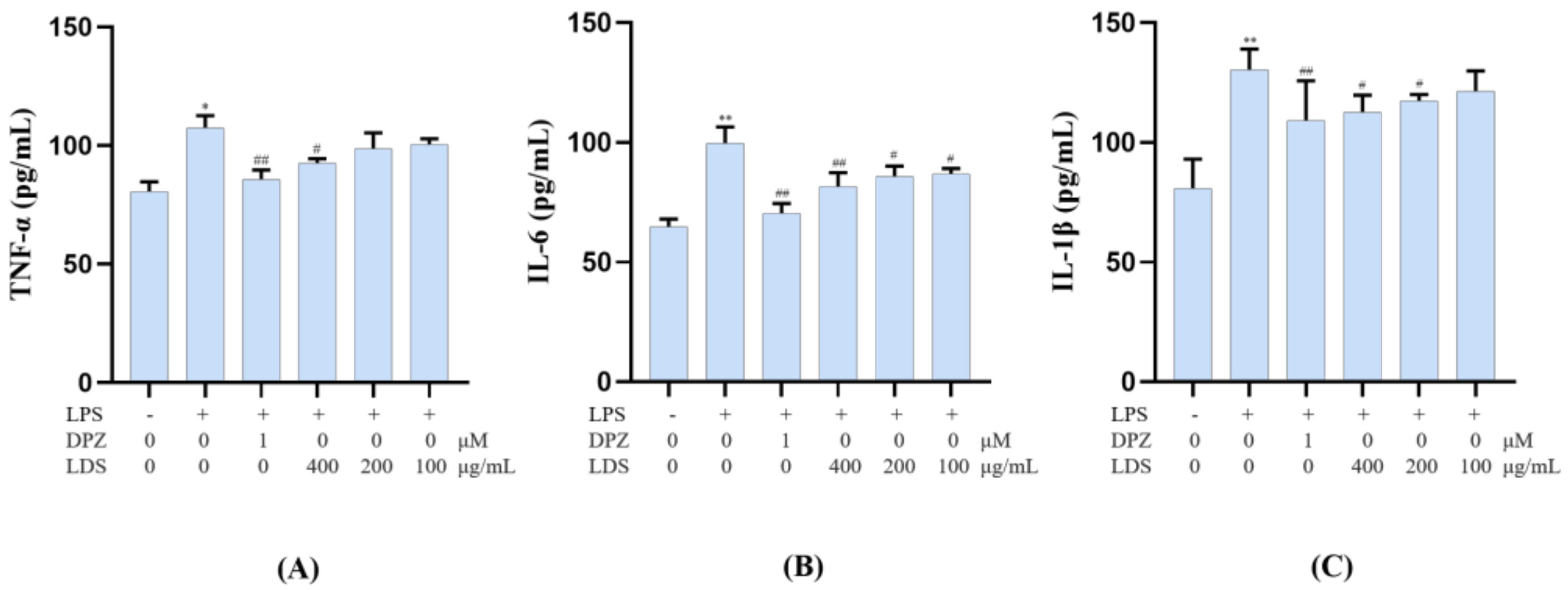
2.6. Modulation of Cellular Production of IL-6, IL-1β and TNF-α by the LDS Extract
3. Materials and Methods
3.1. Materials
3.2. Chemical, Reagents, Equipment
3.3. Sample Preparation
3.4. Construction of SPR Biosensor-UPLC/MS Active Ingredient Recognition System for TREM2 Target
3.4.1. TREM2 Protein Pre-Coupling
3.4.2. Recovery of TREM2-Bound Ingredients
3.4.3. UPLC-QTOF/MS Analysis
3.5. The Lignan-Amides Preconcentration
3.6. Cell Culture Assay
3.6.1. Cell Culture
3.6.2. Cell Viability: CCK-8 Assay
3.6.3. LPS-Induced Model
3.6.4. NO Release Analysis
3.6.5. Western Blotting
3.6.6. Elisa Analysis
3.6.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviations
| Full Name | Acronyms |
| Alzheimer’s disease | AD |
| triggering receptor expressed on myeloid Cells 2 | TREM2 |
| surface plasmon resonance | SPR |
| quadrupole time-of-flight tandem mass spectrometry | UPLC-MS |
| the lignin-amides from Datura metel seeds | LDS |
| inducible Nitric Oxide Synthase | iNOS |
| cyclooxygenase 2 | COX-2 |
| microtubule-associated protein tau | Tau |
| ionized calcium-binding adapter molecule 1 | IBA-1 |
| DNAX-activating protein of 12 kDa | DAP12 |
| Toll-like receptor SX4 | TLR4 |
| Myeloid differentiation factor 88 | MyD88 |
| Recombinant NLR Family, Pyrin Domain Containing Protein 3 | NLRP3 |
| cysteinyl aspartate specific proteinase-1 | Caspase-1 |
| Inflammatory factors Interleukin 1 beta | IL-1β |
| Inflammatory factors Interleukin 6 | IL-6 |
| Tumor necrosis factor-alpha | TNFα |
| caspase recruitment domain protein 9 | CARD9 |
| phosphatidylinositol 3 kinase | PI3K |
| protein kinase B | AKT |
| nuclear factor activated B cells κ-Light chain reinforcement | NF-κB |
| central nervous system | CNS |
| Parkinson’s disease | PD |
| Amyotrophic lateral sclerosis | ALS |
References
- Wang, S.; Mustafa, M.; Yuede, C.M.; Salazar, S.V.; Kong, P.; Long, H.; Ward, M.; Siddiqui, O.; Paul, R.; Gilfillan, S.; et al. Anti-human TREM2 induces microglia proliferation and reduces pathology in an Alzheimer’s disease model. J. Exp. Med. 2020, 217, e20200785. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Liu, W.; Wang, X. TREM2 variants and risk of Alzheimer’s disease: A meta-analysis. Neurol. Sci. 2015, 36, 1881. [Google Scholar] [CrossRef] [PubMed]
- Nicastro, N.; Malpetti, M.; Mak, E.; Williams, G.B.; O’Brien, J.T. Grey matter changes related to microglial activation in alzheimer’s disease. Neurobiol. Aging 2020, 94, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Konishi, H.; Kiyama, H. Non-pathological roles of microglial trem2/DAP12: trem2/DAP12 regulates the physiological functions of microglia from development to aging. Neurochem. Int. 2020, 141, 1048–1078. [Google Scholar] [CrossRef]
- Heslegrave, A.; Heywood, W.; Paterson, R.; Magdalinou, N.; Svensson, J.; Johansson, P.; Ohrfelt, A.; Blennow, K.; Hardy, J.; Schott, J.; et al. Increased cerebrospinal fluid soluble TREM2 concentration in Alzheimer’s disease. Mol. Neurodegener. 2016, 11, 3. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.Z.; Tan, L.; Bi, Y.L.; Shen, X.N.; Xu, W.; Ma, Y.H.; Li, H.Q.; Dong, Q.; Yu, J.T. Dynamic changes of CSF sTREM2 in preclinical Alzheimer’s disease: The CABLE study. Mol. Neurodegener. 2020, 5, 25. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.H.; Yang, H.; Cheng, X.; Yang, Y.L.; Lyu, Y.; Du, G.H. P7 extract of ligusticum chuanxiong hort attenuates lps-induced neuroinflammation in bv2 microglia via down-regulation of tlr4/myd88 pathway. Biochem. Pharm. 2017, 139, 126–127. [Google Scholar] [CrossRef]
- Wang, Y.; Cella, M.; Mallinson, K.; Ulrich, J.D.; Young, K.L.; Robinette, M.L.; Gilfillan, S.; Krishnan, G.M.; Sudhakar, S.; Zinselmeyer, B.H.; et al. TREM2 lipid sensing sustains the microglial response in an Alzheimer’s disease model. Cell 2015, 160, 1061–1071. [Google Scholar] [CrossRef] [Green Version]
- Cui, X.; Qiao, J.; Liu, S.; Wu, M.; Gu, W. Mechanism of trem2/DAP12 complex affecting β-amyloid plaque deposition in alzheimer’s disease modeled mice through mediating inflammatory response. Brain Res. Bull. 2021, 166, 21–28. [Google Scholar] [CrossRef]
- Ewers, M.; Franzmeier, N.; Suárez-Calvet, M.; Morenas-Rodriguez, E.; Caballero, M.A.A.; Kleinberger, G.; Piccio, L.; Cruchaga, C.; Deming, Y.; Dichgans, M.; et al. Increased soluble TREM2 in cerebrospinal fluid is associated with reduced cognitive and clinical decline in Alzheimer’s disease. Sci. Transl. Med. 2019, 11, 6221. [Google Scholar] [CrossRef]
- Tanaka, M.; Saito, S.; Inoue, T.; Satoh-Asahara, N.; Ihara, M. Potential Therapeutic Approaches for Cerebral Amyloid Angiopathy and Alzheimer’s Disease. Int. J. Mol. Sci. 2020, 14, 1992. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.; Tang, J.; Passos, C.D.S.; Nurisso, A.; Simoes-Pires, C.; Ji, M.; Lou, H.; Fan, P. Characterization of Lignanamides from Hemp (Cannabis. sativa. L.) Seed and their Antioxidant and Acetylcholinesterase Inhibitory Activities. J. Agric. Food Chem. 2015, 63, 10611–10619. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Li, H.; Gao, H.; Huang, Y.; Zhou, W.; Yu, Y.; Yao, X. Bioactive Nitrogenous Compounds from Acorus tatarinowii. Magn. Reson. Chem. 2016, 54, 396–399. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Luo, Q.; Fan, P. Cannabisin F from hemp (Cannabis sativa.) seed suppresses lipopolysaccharide-induced inflammatory responses in bv2 microglia as sirt1 modulator. Int. J. Mol. Sci. 2019, 20, 507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Wang, S.; Ji, J.; Lou, H.; Fan, P. Hemp (Cannabis sativa. L.) seed phenylpropionamides composition and effects on memory dysfunction and biomarkers of neuroinflammation induced by lipopolysaccharide in mice. ACS Omega 2018, 3, 15988–15995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nigro, E.; Crescente, G.; Formato, M.; Pecoraro, M.T.; Pacifico, S. Hempseed lignanamides rich-fraction: Chemical investigation and cytotoxicity towards u-87 glioblastoma cells. Molecules 2020, 25, 1049. [Google Scholar] [CrossRef] [Green Version]
- Bingyou, Y.; Haibing, J.; Yan, L.; Zhenpeng, X.; Haixue, K. Study on the chemical constituents of the seeds of Datura. Metel. (IV). Chin. Med. Mater. 2018, 41, 93–98. [Google Scholar]
- Bingyou, Y.; Yan, L.; Xin, W.; Yonggang, X.; Qiuhong, W.; Haixue, K. Studies on the chemical constituents of the seeds of Datura. metel. (I). Chin. Herb. Med. 2013, 44, 1877–1880. [Google Scholar]
- Bingyou, Y.; Yan, L.; Peiyan, Z.; Qiuhong, W.; Haixue, K. Research on the chemical constituents of alkaloids in the seeds of Datura. metel. In Proceedings of the World Conference on Traditional Chinese Medicine Summer Summit and “One Belt One Road” International Symposium on the Development of Traditional Chinese Medicine; 2015; pp. 1–2. Available online: https://d.wanfangdata.com.cn/conference/9013855 (accessed on 12 June 2021).
- You, Y.B.; Xue, K.H.; Hui, Y.S.; Yan, L.; Ping, S.Y. Identification and Quantification of Alkaloid Compounds from Different Parts and Production Areas of Datura metel. L. Heterocycles 2020, 100, 568. [Google Scholar] [CrossRef] [Green Version]
- Murthy, B.K.; Nammi, S.; Kota, M.K.; Rao, R.; Rao, N.K.; Annapurna, A. Evaluation of hypoglycemic and antihyperglycemic effects of Datura metel. (linn.) seeds in normal and alloxan-induced diabetic rats. J. Ethnopharmacol. 2004, 91, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Wannang, N.N.; Ndukwe, H.C.; Nnabuife, C. Evaluation of the analgesic properties of the Datura metel. seeds aqueous extract. J. Med. Plants Res. 2009, 3, 192–195. [Google Scholar]
- Bing-You, Y.; Hai-Bing, J.; Yan, L.; Jing, C.; Hai-Xue, K. Steroids from the seeds of Datura metel. J. Asian Nat. Prod. Res. 2020, 22, 257–263. [Google Scholar]
- Handrick, V.; Vogt, T.; Frolov, A. Profiling of hydroxycinnamic acid amides in Arabidopsis thaliana pollen by tandem mass spectrometry. Anal. Bioanal. Chem. 2010, 398, 2789–2801. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-X.; Shi, Q.; Xiong, Q.-B.; Prasain, J.; Tezuka, Y.; Hareyama, T.; Wang, Z.-T.; Tanaka, K.; Namba, T.; Kadota, S. Tribulusamide A and B, new hepatoprotective lignan amides from the fruits of Tribulus terrestris: Indications of cytoprotective activity in murine hepatocyte culture. Planta Med. 1998, 64, 628–631. [Google Scholar] [CrossRef] [PubMed]
- Young, S.J.; Ae, K.M.; Jo, K.M.; Wanjoo, C.; Yongsoo, K. Acetylcholinesterase Inhibitors from the Stem of Zea mays. Nat. Prod. Sci. 2014, 20, 13–16. [Google Scholar]
- Yang, B.Y.; Yin, X.; Liu, Y.; Ye, H.L.; Kuang, H.X. Bioassay-guided isolation of lignan amides with potential anti-inflammatory effect from the roots of Solanum melongena. L. Phytochem. Lett. 2019, 30, 160–164. [Google Scholar] [CrossRef]
- Zhu, P.F.; Dai, Z.; Wang, B.; Wei, X.; Yu, H.F.; Yan, Z.R.; Zhao, X.; Liu, Y.; Luo, X. The anticancer activities phenolic amides from the stem of Lycium barbarum. Nat. Prod. Bioprospect. 2017, 7, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Cutillo, F.; D’Abrosca, B.; Dellagreca, M.; Zarrelli, A. Chenoalbicin, a Novel Cinnamic Acid Amide Alkaloid from Chenopodium album. Chem. Bio. 2010, 1, 1579–1583. [Google Scholar]
- Chen, H.; Li, Y.J.; Sun, Y.J.; Gong, J.-H.; Du, K.; Zhang, Y.-L.; Su, C.-F.; Han, Q.-Q.; Zheng, X.-K.; Feng, W.S. Lignanamides with potent antihyperlipidemic activities from the root bark of Lycium chinense. Fitoterapia 2017, 122, 119–125. [Google Scholar] [CrossRef] [PubMed]
- National Center for Biotechnology Information. PubChem Compound Summary for CID 101938451. 2021. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/101938451 (accessed on 1 September 2021).
- Choi, H.S.; Cho, J.-Y.; Jin, M.R.; Lee, Y.G.; Kim, S.-J.; Ham, K.-S.; Moon, J.-H. Phenolics, acyl galactopyranosyl glycerol, and lignan amides from Tetragonia tetragonioides. (Pall.) Kuntze. Food Sci. Biotechnol. 2016, 25, 1275–1281. [Google Scholar] [CrossRef]
- Luo, Q.; Yan, X.; Bobrovskaya, L.; Ji, M.; Yuan, H.; Lou, H.; Fan, P. Anti-neuroinflammatory effects of grossamide from hemp seed via suppression of TLR-4-mediated NF-κB signaling pathways in lipopolysaccharide-stimulated BV2 microglia cells. Mol. Cell Biochem. 2017, 428, 129–137. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, S.; Lou, H.; Fan, P. Chemical constituents of hemp (Cannabis sativa. L.) seed with potential anti-neuroinflammatory activity. Phytochem. Lett. 2018, 23, 57–61. [Google Scholar] [CrossRef]
- Wang, Y.; Ji, S.; Zang, W.; Wang, N.; Cao, J.; Li, X.; Sun, C. Identification of phenolic compounds from a unique citrus species, finger lime (citrus australasica) and their inhibition of lps-induced no-releasing in bv-2 cell line. Food Chem. Toxicol. 2019, 129, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H.; Lv, H.N.; Cui, Q.H.; Tu, P.F.; Zeng, K.W. Isosibiricin inhibits microglial activation by targeting the dopamine d1/d2 receptor-dependent nlrp3/caspase-1 inflammasome pathway. Acta Pharm. Sin. 2020, 41, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Sloane, J.A.; Hollander, W.; Moss, M.B.; Rosene, D.L.; Carmela, R. Abraham Increased microglial activation and protein nitration in white matter of the aging monkey. Neurobiol. Aging 1999, 20, 395. [Google Scholar] [CrossRef]
- Wysscoray, T.; Lucin, K.M. Immune activation in brain aging and neurodegeneration: Too much or too little. Neuron 2009, 64, 110–122. [Google Scholar]
- El Khoury, J.; Toft, M.; Hickman, S.E.; Means, T.K.; Terada, K.; Geula, C.; Luster, A.D. Ccr2 Deficiency Impairs Microglial Accumulation and Accelerates Progression of Alzheimer’s Disease. Nat. Med. 2007, 13, 432–438. [Google Scholar] [CrossRef]
- Kitazawa, M.; Oddo, S.; Yamasaki, T.R.; Green, K.N.; LaFerla, F.M. Lipopolysaccharide-Induced Inflammation Exacerbates Tau Pathology by a Cyclin-Dependent Kinase 5-Mediated Pathway in a Transgenic Model of Alzheimer’s Disease. J. Neurosci. 2005, 25, 8843–8853. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.C.; Rizer, J.; Selenica, M.-L.B.; Reid, P.; Kraft, C.; Johnson, A.; Blair, L.; Gordon, M.N.; Dickey, C.; Morgan, D. LPS-induced inflammation exacerbates phospho-tau pathology in rTg4510 mice. J. Neuroinflamm. 2010, 7, 56. [Google Scholar] [CrossRef] [Green Version]
- Papasozomenos, S.C.; Binder, L.I. Phosphorylation determines two distinct species of tau in the central nervous system. Cell. Motil. Cytoskelet. 1987, 8, 210–226. [Google Scholar] [CrossRef]
- Ghoshal, N.; García-Sierra Fu, Y.; Beckett, L.A.; Mufson, E.J.; Kuret, J.; Berry, R.W.; Binder, L.I. Tau-66: Evidence for a novel tau conformation in Alzheimer’s disease. J. Neurochem. 2010, 77, 1372–1385. [Google Scholar] [CrossRef] [Green Version]
- Uchihara, T.; Tsuchiya, K.; Nakamura, A.; Ikeda, K. Appearance of tau-2 immunoreactivity in glial cells in human brain with cerebral infarction. Neurosci. Lett. 2000, 286, 99–102. [Google Scholar] [CrossRef]
- Uchihara, T.; Nakamura, A.; Arai, T.; Ikeda, K.; Tsuchiya, K. Microglial tau undergoes phosphorylation-independent modification after ischemia. Glia 2004, 45, 180–187. [Google Scholar] [CrossRef]
- Majerova, P.; Zilkova, M.; Kazmerova, Z.; Kovac, A.; Paholikova, K.; Kovacech, B.; Zilka, N.; Novak, M. Microglia display modest phagocytic capacity for extracellular tau oligomers. J. Neuroinflamm. 2014, 11, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; Mishra, G.; Maurya, A.; Rajendra, A.; Komal, K.; Abhimanyu, T.; Arati, R.; Gopal, R.K.; Bhupesh, S.; Giriraj, K.T.; et al. Role of TREM2 in Alzheimer’s disease and its consequences on β-amyloid, Tau and neurofibrillary tangles. Curr. Alzheimer Res. 2019, 16, 1216–1229. [Google Scholar] [CrossRef]
- Marucci, G.; Moruzzi, M.; Amenta, F. Donepezil in the treatment of Alzheimer’s disease. In Diagnosis and Management in Dementia; Academic Press: Cambridge, MA, USA, 2020. [Google Scholar]
- Atagi, Y.; Liu, C.; Painter, M.M.; Chen, X.; Verbeeck, C.; Zheng, H.; Li, X.; Rademakers, R.; Kang, S.S.; Xu, H.; et al. Apolipoprotein E Is a Ligand for Triggering Receptor Expressed on Myeloid Cells 2 (TREM2). J. Biol. Chem. 2015, 290, 26043–26050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, Z.; Gu, M.; Zhou, J.; Zhang, W.; Zhao, N.; Lü, Y.; Yu, W. Triggering receptor expressed on myeloid Cells 2 activation downregulates toll-like receptor 4 expression and ameliorates cognitive impairment in the Aβ1-42-induced Alzheimer’s disease mouse model. Synapse 2020, 10, e22161. [Google Scholar] [CrossRef]
- Li, C.; Zhao, B.; Lin, C.; Gong, Z.; An, X. TREM2 inhibits inflammatory responses in mouse microglia by suppressing the PI3K/NF-κB signaling. Cell Biol. Int. 2019, 43, 360–372. [Google Scholar] [CrossRef]
- Gratuze, M.; Leyns, C.; Holtzman, D.M. New insights into the role of TREM2 in Alzheimer’s disease. Mol. Neurodegener. 2018, 13, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | Observed tR (min) | Observed Ms (m/z) | Adduct | Compounds | Products Ions |
|---|---|---|---|---|---|
| 1 | 12.80 | 625.2607 | H | cannabisin G | 625 [M+H]+ 488 [M+H-C8H10NO]+ |
| 2 | 12.94 | 643.2651 | H | lyciumamide K | 643 [M+H]+ |
| 3 | 13.87 | 625.2607 | H | cannabisin F | 625 [M+H]+ 462 [M+H–C8H10NO-CO]+ |
| 4 | 15.60 | 625.2607 | H | cannabisin D | 625 [M+H]+ 488 [M+H–C8H10NO]+ 462 [M+H–C8H10NO-CO]+ |
| Peak No. | Observed tR (min) | Observed MS (m/z) | Adduct | Compounds | Mass Error (ppm) | Products Ions | Reference |
|---|---|---|---|---|---|---|---|
| 1 | 8.78 | 314.1388 | H | N-trans-feruloyl tyramine | −1.3 | 314 [M+H]+ 177 [M+H C8H10NO]+ | [18] |
| 2 | 8.78 | 314.1388 | H | N-cis-feruloyl tyramine | −1.3 | 314 [M+H]+ 177 [M+H-C8H10NO]+ | [18] |
| 3 | 8.80 | 510.2124 | H | cannabisin H | 2.4 | 510 [M+H]+ 312 [M+H-H2O-3CH3COOH]+ | [18] |
| 4 | 12.41 | 625.2547 | H | cannabisin G (2) | −0.43 | 643 [M+H+H2O]+ 625 [M+H]+ 488 [M+H-C8H10NO]+ | |
| 5 | 12.50 | 643.2651 | H | lyciumamide K | −0.67 | 643 [M+H]+ 625 [M+H-H2O]+ 462 [M+H-H2O-C10H11NO2]+ | [18] |
| 6 | 12.51 | 625.2550 | H | Tribulusamide A | 0.05 | 625 [M+H]+ 488 [M+H-C8H10NO]+ 462 [M+H-C8H10NO-H2O]+ 177 [M+H-C27H26NO6]+ | [23] |
| 7 | 12.75 | 492.2021 | H | grossamide K | −0.21 | 492 [M+H]+ 462 [M+H-OCH2]+ 357 [M+H-C8H10NO]+ | [18] |
| 8 | 12.78 | 492.2021 | H | cis-grossamide K | −0.21 | 492 [M+H]+ 462 [M+H-OCH2]+ 357 [M+H-C8H10NO]+ | [15] |
| 9 | 13.09 | 643.2657 | H | cannabisin E | 0.26 | 643 [M+H]+ 462 [M+H-H2O-C9H11NO]+ 113 [M+H-C30H28NO8]+ | [16] |
| 10 | 13.49 | 268.1342 | H | N-trans-cinnamoyl tyramine | 1.76 | 268 [M+H]+ 136 [M+H-C7H16O3]+ 128 [M+H-H2O-C10H10]+ 113 [M+H-C11H7O]+ | [18] |
| 11 | 13.89 | 625.2556 | H | cannabisin F | 1.01 | 625 [M+H]+ | [18] |
| 12 | 15.05 | 337.1552 | H | N-trans-ferulyl tryptamine | 0.02 | 337 [M+H]+ 177 [M+H C10H11N2]+ | [24] |
| 13 | 15.08 | 641.2493 | H | compound 13 * | −0.9 | 641 [M+H]+ | [31] |
| 14 | 15.26 | 625.2553 | H | cannabisin D | 0.53 | 625 [M+H]+ 488 [M+H-C8H10NO]+ 462 [M+H-C8H10NO-CO]+ | [18] |
| 15 | 15.63 | 492.2021 | H | lyciumamide C | −0.21 | 492 [M+H]+ 462 [M+H-OCH3]+ | [26] |
| 16 | 15.85 | 641.2493 | H | Melongenamide B | −0.91 | 641 [M+H]+ 625 [M+H-H2O]+ 537 [M+H-3H2O-CH3COOH]+ | [18] |
| 17 | 16.45 | 657.2644 | H+CH3COOH | cannabisin A | 2.1 | 656 [M+H+CH3COOH]+ 494 [M+H+CH3COOH-C9H10NO2]+ | [32] |
| 18 | 17.06 | 671.2614 | H | Melongenamide F | 1.5 | 671 [M+H]+ 437 [M+H-C12H13NO4]+ 318 [M+H-C20H21NO5]+ | [25] |
| 19 | 17.35 | 625.2556 | H | lyciumamide B | 1.1 | 625 [M+H]+ 462 [M+H-H2O-CH3OH-C8H11NO]+ | [18] |
| 20 | 17.67 | 671.2471 | H | Melongenamide G | 1.5 | 671 [M+H]+ 639 [M+H-2H2O]+ 437 [M+2H-C12H13NO4]+ 419 [M+2H-C12H13NO4-H2O]+ | [25] |
| 21 | 17.86 | 625.2550 | H+H2O | Chenoalbicin | −4.4 | 625 [M+H+H2O]+ | [27] |
| 22 | 18.86 | 625.2553 | H | (2aS,3aS) lyciumamide D | 0.37 | 625 [M+H]+ 113 [M+H-C30H25NO7]+ | [18] |
| 23 | 19.23 | 936.3699 | H | Thoreliamide C | −0.45 | 936 [M+H]+ 625 [M+H-C18H17NO4]+ 454 [M+H-C25H23NO9]+ 113 [M+H-C48H42N2O11]+ | [18] |
| 24 | 20.82 | 623.2397 | Na | lyciumamide F | 4.3 | 623 [M+Na]+ | [28] |
| 25 | 22.17 | 625.2470 | H | grossamide | −0.51 | 625 [M+H]+ 462 [M+H-C9H9NO2]+ 316 [M+H-C18H18NO2]+ 298 [M+H-H2O-C18H18NO2]+ 113 [M+H-C30H25NO7]+ | [18] |
| 26 | 22.53 | 936.3713 | H | cannabisin O | 0.6 | 936 [M+H]+ 599 [M+H-H2O-CH3OH-CO-C17H17NO2]+ | [12] |
| 27 | 23.94 | 538.2020 | H+HCOOH | hibiscuwanin B | −0.37 | 538 [M+H+HCOOH]+ 439 [M+H+HCOOH-2H2O-2HOCH3]+ | [18] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.-Y.; Liu, Y.; Li, X.-M.; Algradi, A.M.; Jiang, H.; Sun, Y.-P.; Guan, W.; Pan, J.; Kuang, H.-X.; Yang, B.-Y. Discovery of Active Ingredients Targeted TREM2 by SPR Biosensor-UPLC/MS Recognition System, and Investigating the Mechanism of Anti-Neuroinflammatory Activity on the Lignin-Amides from Datura metel Seeds. Molecules 2021, 26, 5946. https://doi.org/10.3390/molecules26195946
Wang S-Y, Liu Y, Li X-M, Algradi AM, Jiang H, Sun Y-P, Guan W, Pan J, Kuang H-X, Yang B-Y. Discovery of Active Ingredients Targeted TREM2 by SPR Biosensor-UPLC/MS Recognition System, and Investigating the Mechanism of Anti-Neuroinflammatory Activity on the Lignin-Amides from Datura metel Seeds. Molecules. 2021; 26(19):5946. https://doi.org/10.3390/molecules26195946
Chicago/Turabian StyleWang, Si-Yi, Yan Liu, Xiao-Mao Li, Adnan Mohammed Algradi, Hai Jiang, Yan-Ping Sun, Wei Guan, Juan Pan, Hai-Xue Kuang, and Bing-You Yang. 2021. "Discovery of Active Ingredients Targeted TREM2 by SPR Biosensor-UPLC/MS Recognition System, and Investigating the Mechanism of Anti-Neuroinflammatory Activity on the Lignin-Amides from Datura metel Seeds" Molecules 26, no. 19: 5946. https://doi.org/10.3390/molecules26195946
APA StyleWang, S.-Y., Liu, Y., Li, X.-M., Algradi, A. M., Jiang, H., Sun, Y.-P., Guan, W., Pan, J., Kuang, H.-X., & Yang, B.-Y. (2021). Discovery of Active Ingredients Targeted TREM2 by SPR Biosensor-UPLC/MS Recognition System, and Investigating the Mechanism of Anti-Neuroinflammatory Activity on the Lignin-Amides from Datura metel Seeds. Molecules, 26(19), 5946. https://doi.org/10.3390/molecules26195946