
Modulation of the NOTCH1 Pathway by LUNATIC FRINGE Is Dominant over That of MANIC or RADICAL FRINGE

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. The Modification of NOTCH1 O-Fucose Sites by Endogenous Fringes in Different Cells Determines the Efficiency of Its Stimulation by Ligands
2.2. The Effect of LFNG on N1 Signaling Is Dominant over MFNG and RFNG
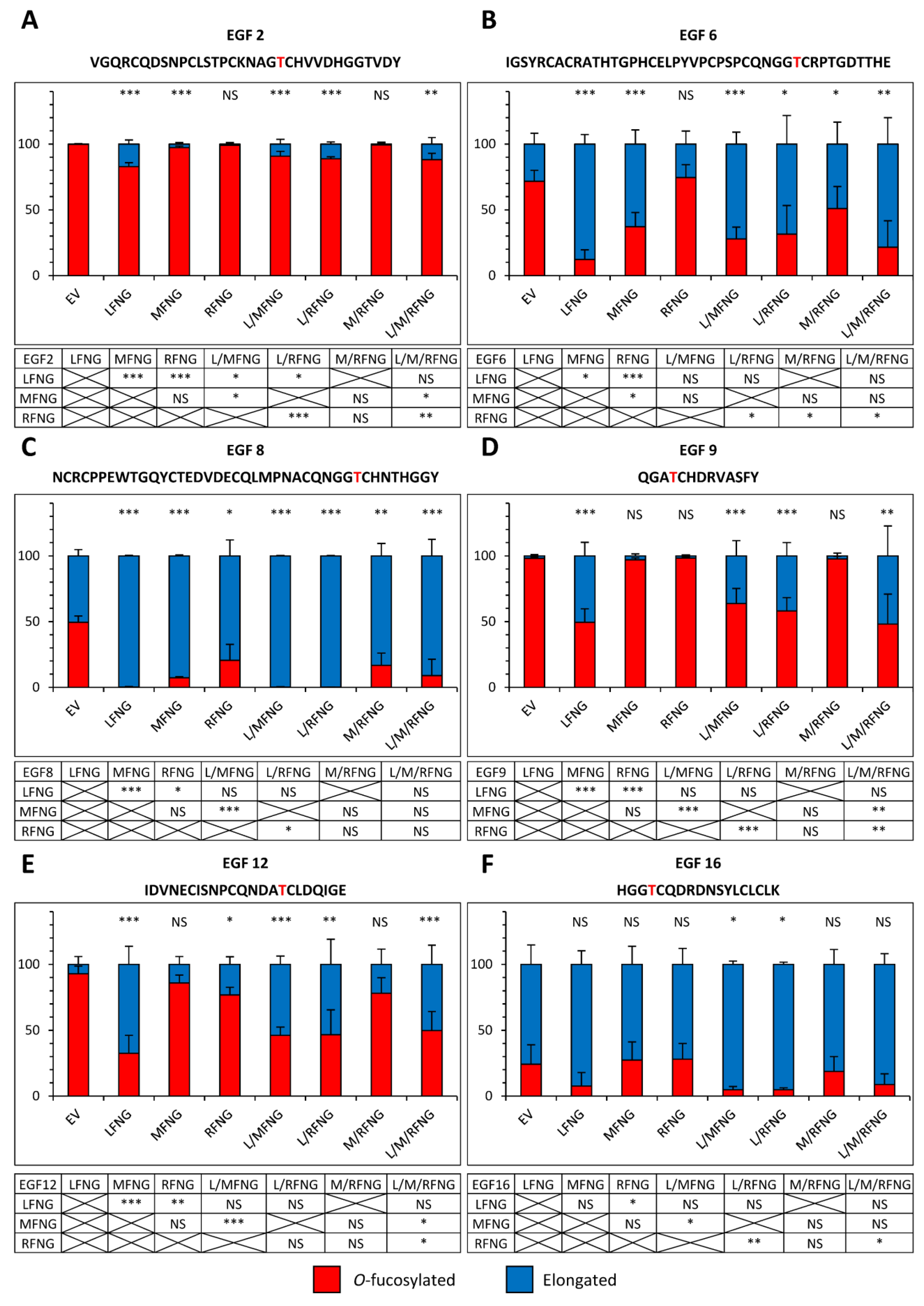
2.3. The Dominance of LFNG over MFNG and RFNG Is Due to the Ability to Modify a Larger Number of O-Fucose Sites with Greater Efficiency
3. Discussion
4. Materials and Methods
4.1. Plasmids
4.2. Cell Culture
4.3. RNA Isolation and qRT-PCR Analysis
4.4. Cell-Based Co-Culture N1 Activation Assay
4.5. Protein Expression and Purification
4.6. Mass Spectral Analysis
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Artavanis-Tsakonas, S.; Rand, M.D.; Lake, R.J. Notch Signaling: Cell Fate Control and Signal Integration in Development. Science 1999, 284, 770–776. [Google Scholar] [CrossRef]
- Kopan, R.; Ilagan, M.X.G. The Canonical Notch Signaling Pathway: Unfolding the Activation Mechanism. Cell 2009, 137, 216–233. [Google Scholar] [CrossRef]
- Wouters, M.A.; Rigoutsos, I.; Chu, C.K.; Feng, L.L.; Sparrow, D.B.; Dunwoodie, S.L. Evolution of Distinct EGF Domains with Specific Functions. Protein Sci. 2005, 14, 1091–1103. [Google Scholar] [CrossRef] [PubMed]
- Kakuda, S.; Haltiwanger, R.S. Deciphering the Fringe-Mediated Notch Code: Identification of Activating and Inhibiting Sites Allowing Discrimination between Ligands. Dev. Cell 2017, 40, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, H.; Schneider, M.; Williamson, D.B.; Ito, A.; Takeuchi, M.; Handford, P.A.; Haltiwanger, R.S. Two Novel Protein O-Glucosyltransferases That Modify Sites Distinct from POGLUT1 and Affect Notch Trafficking and Signaling. Proc. Natl. Acad. Sci. USA 2018, 115, E8395–E8402. [Google Scholar] [CrossRef]
- Rana, N.A.; Nita-Lazar, A.; Takeuchi, H.; Kakuda, S.; Luther, K.B.; Haltiwanger, R.S. O-Glucose Trisaccharide Is Present at High but Variable Stoichiometry at Multiple Sites on Mouse Notch1. J. Biol. Chem. 2011, 286, 31623–31637. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shao, L.; Shi, S.; Harris, R.J.; Spellman, M.W.; Stanley, P.; Haltiwanger, R.S. Modification of Epidermal Growth Factor-like Repeats With O-Fucose. J. Biol. Chem. 2001, 276, 40338–40345. [Google Scholar] [CrossRef]
- Kakuda, S.; LoPilato, R.K.; Ito, A.; Haltiwanger, R.S. Canonical Notch Ligands and Fringes Have Distinct Effects on NOTCH1 and NOTCH2. J. Biol. Chem. 2020, 295, 14710–14722. [Google Scholar] [CrossRef]
- Harris, R.J.; Spellman, M.W. O-Linked Fucose and Other Post-Translational Modifications Unique to EGF Modules. Glycobiology 1993, 3, 219–224. [Google Scholar] [CrossRef]
- Lira-Navarrete, E.; Valero-González, J.; Villanueva, R.; Martínez-Júlvez, M.; Tejero, T.; Merino, P.; Panjikar, S.; Hurtado-Guerrero, R. Structural Insights into the Mechanism of Protein O-Fucosylation. PLoS ONE 2011, 6, e25365. [Google Scholar] [CrossRef]
- Holdener, B.C.; Haltiwanger, R.S. Protein O-Fucosylation: Structure and Function. Curr. Opin. Struct. Biol. 2019, 56, 78–86. [Google Scholar] [CrossRef]
- Wang, Y.; Spellman, M.W. Purification and Characterization of a GDP-Fucose:Polypeptide Fucosyltransferase from Chinese Hamster Ovary Cells. J. Biol. Chem. 1998, 273, 8112–8118. [Google Scholar] [CrossRef]
- Varshney, S.; Stanley, P. Multiple Roles for O-glycans in Notch Signalling. FEBS Lett. 2018, 592, 3819–3834. [Google Scholar] [CrossRef]
- Takeuchi, H.; Yu, H.; Hao, H.; Takeuchi, M.; Ito, A.; Li, H.; Haltiwanger, R.S. O-Glycosylation Modulates the Stability of Epidermal Growth Factor-like Repeats and Thereby Regulates Notch Trafficking. J. Biol. Chem. 2017, 292, 15964–15973. [Google Scholar] [CrossRef]
- Shi, S.; Stanley, P. Protein O-Fucosyltransferase 1 Is an Essential Component of Notch Signaling Pathways. Proc. Natl. Acad. Sci. USA 2003, 100, 5234–5239. [Google Scholar] [CrossRef]
- Varshney, S.; Wei, H.-X.; Batista, F.; Nauman, M.; Sundaram, S.; Siminovitch, K.; Tanwar, A.; Stanley, P. A Modifier in the 129S2/SvPasCrl Genome Is Responsible for the Viability of Notch1[12f/12f] Mice. BMC Dev. Biol. 2019, 19, 1–11. [Google Scholar] [CrossRef]
- Luca, V.C.; Jude, K.M.; Pierce, N.W.; Nachury, M.V.; Fischer, S.; Garcia, K.C. Structural Basis for Notch1 Engagement of Delta-like 4. Science 2015, 347, 847–853. [Google Scholar] [CrossRef]
- Luca, V.C.; Kim, B.C.; Ge, C.; Kakuda, S.; Wu, D.; Roein-Peikar, M.; Haltiwanger, R.S.; Zhu, C.; Ha, T.; Garcia, K.C. Notch-Jagged Complex Structure Implicates a Catch Bond in Tuning Ligand Sensitivity. Science 2017, 355, 1320–1324. [Google Scholar] [CrossRef]
- Moloney, D.J.; Shair, L.H.; Lu, F.M.; Xia, J.; Locke, R.; Matta, K.L.; Haltiwanger, R.S. Mammalian Notch1 Is Modified with Two Unusual Forms of O-Linked Glycosylation Found on Epidermal Growth Factor-like Modules. J. Biol. Chem. 2000, 275, 9604–9611. [Google Scholar] [CrossRef]
- Johnston, S.H.; Rauskolb, C.; Wilson, R.; Prabhakaran, B.; Irvine, K.D.; Vogt, T.F. A Family of Mammalian Fringe Genes Implicated in Boundary Determination and the Notch Pathway. Development 1997, 124, 2245–2254. [Google Scholar] [CrossRef]
- Evrard, Y.A.; Lun, Y.; Aulehla, A.; Gan, L.; Johnson, R.L. Lunatic Fringe Is an Essential Mediator of Somite Segmentation and Patterning. Nature 1998, 394, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Gridley, T. Defects in Somite Formation in Lunatic Fringe-Deficient Mice. Nature 1998, 394, 374–377. [Google Scholar] [CrossRef] [PubMed]
- Sparrow, D.B.; Chapman, G.; Wouters, M.A.; Whittock, N.V.; Ellard, S.; Fatkin, D.; Turnpenny, P.D.; Kusumi, K.; Sillence, D.; Dunwoodie, S.L. Mutation of the LUNATIC FRINGE Gene in Humans Causes Spondylocostal Dysostosis with a Severe Vertebral Phenotype. Am. J. Hum. Genet. 2006, 78, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Svensson, P.; Bergqvist, I.; Norlin, S.; Edlund, H. MFng Is Dispensable for Mouse Pancreas Development and Function. Mol. Cell Biol. 2009, 29, 2129–2138. [Google Scholar] [CrossRef]
- Moran, J.L.; Shifley, E.T.; Levorse, J.M.; Mani, S.; Ostmann, K.; Perez-Balaguer, A.; Walker, D.M.; Vogt, T.F.; Cole, S.E. Manic Fringe Is Not Required for Embryonic Development, and Fringe Family Members Do Not Exhibit Redundant Functions in the Axial Skeleton, Limb, or Hindbrain. Dev. Dyn. 2009, 238, 1803–1812. [Google Scholar] [CrossRef]
- Hahn, K.L. Lunatic Fringe Null Female Mice Are Infertile Due to Defects in Meiotic Maturation. Development 2005, 132, 817–828. [Google Scholar] [CrossRef]
- Zhang, N.; Norton, C.R.; Gridley, T. Segmentation Defects of Notch Pathway Mutants and Absence of a Synergistic Phenotype in Lunatic Fringe/Radical Fringe Double Mutant Mice. Genes 2002, 33, 21–28. [Google Scholar] [CrossRef]
- Urata, Y.; Takeuchi, H. Effects of Notch Glycosylation on Health and Diseases. Dev. Growth Differ. 2020, 62, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Kumar, V.; Wei, H.-X.; Qiu, J.; Stanley, P. Lunatic, Manic, and Radical Fringe Each Promote T and B Cell Development. J. Immunol. 2016, 196, 232–243. [Google Scholar] [CrossRef]
- Gu, W.; Xu, W.; Ding, T.; Guo, X. Fringe Controls Naïve CD4+T Cells Differentiation through Modulating Notch Signaling in Asthmatic Rat Models. PLoS ONE 2012, 7, e47288. [Google Scholar] [CrossRef]
- Benedito, R.; Roca, C.; Sörensen, I.; Adams, S.; Gossler, A.; Fruttiger, M.; Adams, R.H. The Notch Ligands Dll4 and Jagged1 Have Opposing Effects on Angiogenesis. Cell 2009, 137, 1124–1135. [Google Scholar] [CrossRef]
- Thélu, J.; Viallet, J.P.; Dhouailly, D. Differential Expression Pattern of the Three Fringe Genes Is Associated with Epidermal Differentiation. J. Investig. Derm. 1998, 111, 903–906. [Google Scholar] [CrossRef]
- Hahn, K.L.; Beres, B.; Rowton, M.J.; Skinner, M.K.; Chang, Y.; Rawls, A.; Wilson-Rawls, J. A Deficiency of Lunatic Fringe Is Associated with Cystic Dilation of the Rete Testis. Reproduction 2009, 137, 79–93. [Google Scholar] [CrossRef][Green Version]
- Moloney, D.J.; Panin, V.M.; Johnston, S.H.; Chen, J.; Shao, L.; Wilson, R.; Wang, Y.; Stanley, P.; Irvine, K.D.; Haltiwanger, R.S.; et al. Fringe Is a Glycosyltransferase That Modifies Notch. Nature 2000, 406, 369–375. [Google Scholar] [CrossRef]
- Yang, L.-T.; Nichols, J.T.; Yao, C.; Manilay, J.O.; Robey, E.A.; Weinmaster, G. Fringe Glycosyltransferases Differentially Modulate Notch1 Proteolysis Induced by Delta1 and Jagged1. Mol. Biol. Cell 2005, 16, 927–942. [Google Scholar] [CrossRef]
- Hou, X.; Tashima, Y.; Stanley, P. Galactose Differentially Modulates Lunatic and Manic Fringe Effects on Delta1-Induced NOTCH Signaling. J. Biol. Chem. 2012, 287, 474–483. [Google Scholar] [CrossRef]
- LeBon, L.; Lee, T.V.; Sprinzak, D.; Jafar-Nejad, H.; Elowitz, M.B. Fringe Proteins Modulate Notch-Ligand Cis and Trans Interactions to Specify Signaling States. eLife 2014, 3, e02950. [Google Scholar] [CrossRef]
- Gordon, W.R.; Zimmerman, B.; He, L.; Miles, L.J.; Huang, J.; Tiyanont, K.; McArthur, D.G.; Aster, J.C.; Perrimon, N.; Loparo, J.J.; et al. Mechanical Allostery: Evidence for a Force Requirement in the Proteolytic Activation of Notch. Dev. Cell 2015, 33, 729–736. [Google Scholar] [CrossRef] [PubMed]
- McMillan, B.J.; Zimmerman, B.; Egan, E.D.; Lofgren, M.; Xu, X.; Hesser, A.; Blacklow, S.C. Structure of Human POFUT1, Its Requirement in Ligand-Independent Oncogenic Notch Signaling, and Functional Effects of Dowling-Degos Mutations. Glycobiology 2017, 27, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Shao, L.; Moloney, D.J.; Haltiwanger, R. Fringe Modifies O-Fucose on Mouse Notch1 at Epidermal Growth Factor-like Repeats within the Ligand-Binding Site and the Abruptex Region. J. Biol. Chem. 2003, 278, 7775–7782. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Nita-Lazar, A.; Haltiwanger, R.S. Two Distinct Pathways for O-Fucosylation of Epidermal Growth Factor-like or Thrombospondin Type 1 Repeats. J. Biol. Chem. 2006, 281, 9385–9392. [Google Scholar] [CrossRef]
- Rampal, R.; Li, A.S.Y.; Moloney, D.J.; Georgiou, S.A.; Luther, K.B.; Nita-Lazar, A.; Haltiwanger, R.S. Lunatic Fringe, Manic Fringe, and Radical Fringe Recognize Similar Specificity Determinants in O-Fucosylated Epidermal Growth Factor-like Repeats. J. Biol. Chem. 2005, 280, 42454–42463. [Google Scholar] [CrossRef]
- Nye, J.S.; Kopan, R.; Axel, R. An Activated Notch Suppresses Neurogenesis and Myogenesis but Not Gliogenesis in Mammalian Cells. Development 1994, 120, 2421–2430. [Google Scholar] [CrossRef]
- Nairn, A.V.; Kinoshita-Toyoda, A.; Toyoda, H.; Xie, J.; Harris, K.; Dalton, S.; Kulik, M.; Pierce, J.M.; Toida, T.; Moremen, K.W.; et al. Glycomics of Proteoglycan Biosynthesis in Murine Embryonic Stem Cell Differentiation. J. Proteome Res. 2007, 6, 4374–4387. [Google Scholar] [CrossRef]
- Nairn, A.V.; dela Rosa, M.; Moremen, K.W. Transcript Analysis of Stem Cells. Methods Enzymol. 2010, 479, 73–91. [Google Scholar] [CrossRef]
- Yamamoto, S.; Charng, W.-L.; Rana, N.A.; Kakuda, S.; Jaiswal, M.; Bayat, V.; Xiong, B.; Zhang, K.; Sandoval, H.; David, G.; et al. A Mutation in EGF Repeat-8 of Notch Discriminates Between Serrate/Jagged and Delta Family Ligands. Science 2012, 338, 1229–1232. [Google Scholar] [CrossRef]
- Bern, M.W.; Kil, Y.J. Two-Dimensional Target Decoy Strategy for Shotgun Proteomics. J. Proteome Res. 2011, 10, 5296–5301. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE Database and Related Tools and Resources in 2019: Improving Support for Quantification Data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pennarubia, F.; Nairn, A.V.; Takeuchi, M.; Moremen, K.W.; Haltiwanger, R.S. Modulation of the NOTCH1 Pathway by LUNATIC FRINGE Is Dominant over That of MANIC or RADICAL FRINGE. Molecules 2021, 26, 5942. https://doi.org/10.3390/molecules26195942
Pennarubia F, Nairn AV, Takeuchi M, Moremen KW, Haltiwanger RS. Modulation of the NOTCH1 Pathway by LUNATIC FRINGE Is Dominant over That of MANIC or RADICAL FRINGE. Molecules. 2021; 26(19):5942. https://doi.org/10.3390/molecules26195942
Chicago/Turabian StylePennarubia, Florian, Alison V. Nairn, Megumi Takeuchi, Kelley W. Moremen, and Robert S. Haltiwanger. 2021. "Modulation of the NOTCH1 Pathway by LUNATIC FRINGE Is Dominant over That of MANIC or RADICAL FRINGE" Molecules 26, no. 19: 5942. https://doi.org/10.3390/molecules26195942
APA StylePennarubia, F., Nairn, A. V., Takeuchi, M., Moremen, K. W., & Haltiwanger, R. S. (2021). Modulation of the NOTCH1 Pathway by LUNATIC FRINGE Is Dominant over That of MANIC or RADICAL FRINGE. Molecules, 26(19), 5942. https://doi.org/10.3390/molecules26195942