
Topical Nano Clove/Thyme Gel against Genetically Identified Clinical Skin Isolates: In Vivo Targeting Behavioral Alteration and IGF-1/pFOXO-1/PPAR γ Cues

 , , ,
, , , 
Abstract
:1. Introduction
2. Material and Methods
2.1. Essential Oil (EOs) Study Samples
2.2. Authentication of Commercial EOs
2.3. Antioxidant Determination Using 2,2-Diphenyl-1-Picrylhydrazyl (DPPH) and Ferric Reducing Antioxidant Power (FRAP) Analyses
2.4. Development of Nano-Emulgels (NEG)
Characterization of NEGs and Selected Formula
2.5. Antibacterial Bioassays
2.5.1. Bacterial Culturing and Identification
2.5.2. Biochemical Identification of Bacterial Isolates Using Vitek2 Compact System
2.5.3. Molecular Identification of Clinically Isolated Bacteria
DNA Extraction
Analysis of Polymerase Chain Reaction (PCR) Products
Microbial Sequencing
2.5.4. Bioactivity Assay Using the Cup Agar Diffusion Method
2.5.5. Determination of Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC)
2.5.6. Biofilm Inhibition Assay, Quantification, and Percentage of Inhibition
2.6. In Vivo Anti-Skin Infection Potential of the Optimized Selected NEG
2.6.1. Behavioral Open Field Tests
2.6.2. Western Blotting
2.6.3. ELISA Parameters
2.6.4. RT-PCR Analysis
2.6.5. Histopathological Examinations
2.7. Statistical Analysis
3. Results and Discussion
3.1. GC-MS Chromatographic Analyses
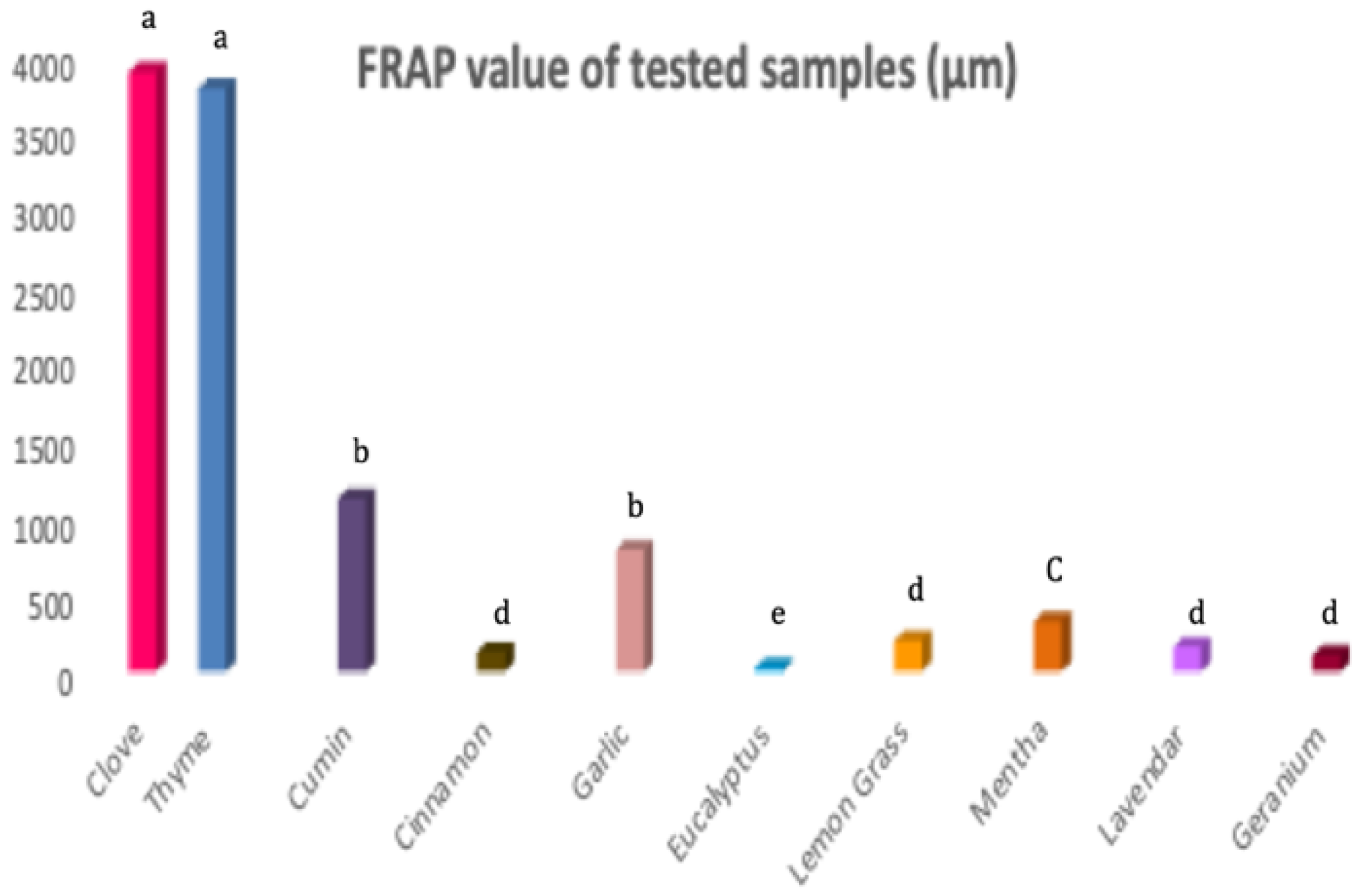
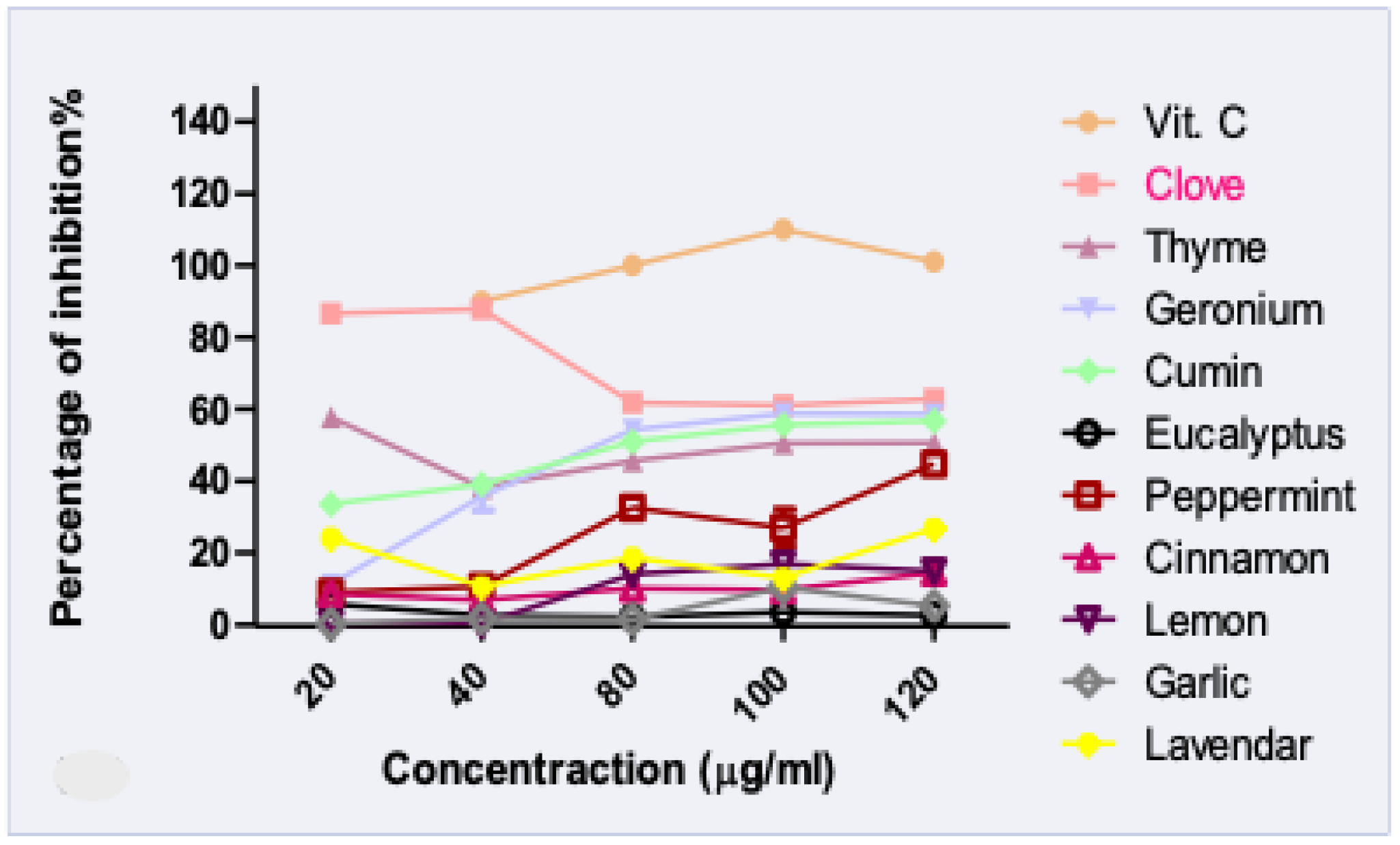
3.2. Antioxidant Activity of EOs
3.3. Characterization of the Prepared NEGs
3.4. In Vitro Antibacterial Activity of NEG8
3.4.1. Identification of Clinically Isolated Bacteria
Vitek Identification of Bacteria Isolate
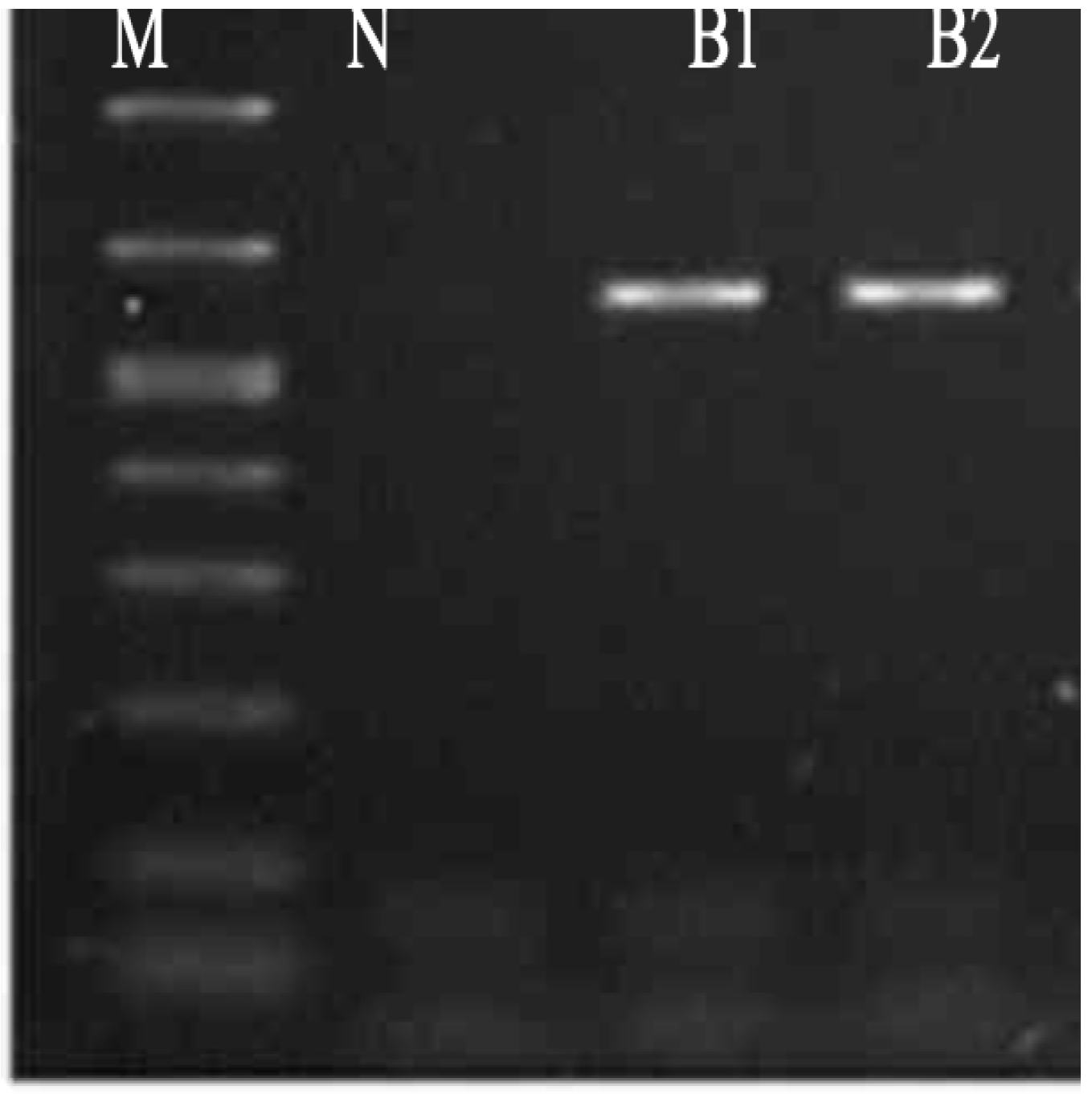
Molecular Identification of Bacterial Isolates
3.4.2. Sensitivity Test, MIC and MBC Assays of NEG8 against Bacterial Isolates
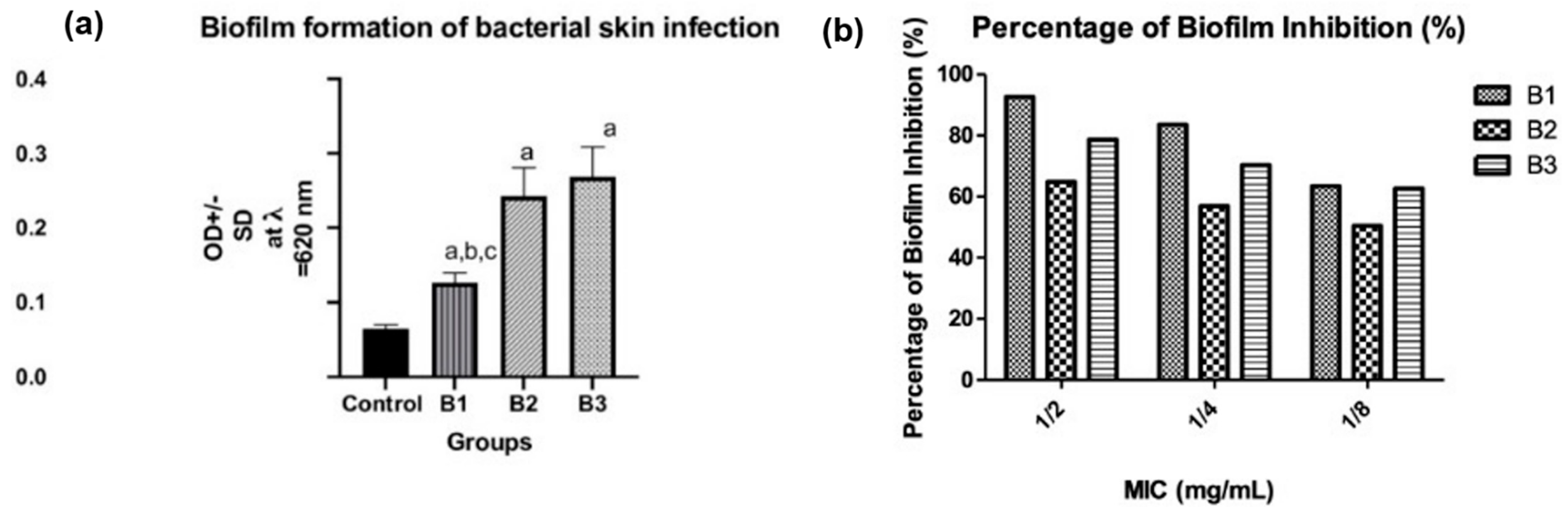
3.4.3. Biofilm Formation, Quantification, and Percentage of Inhibition by NEG8
3.5. In Vivo Antimicrobial Potential of NEG8
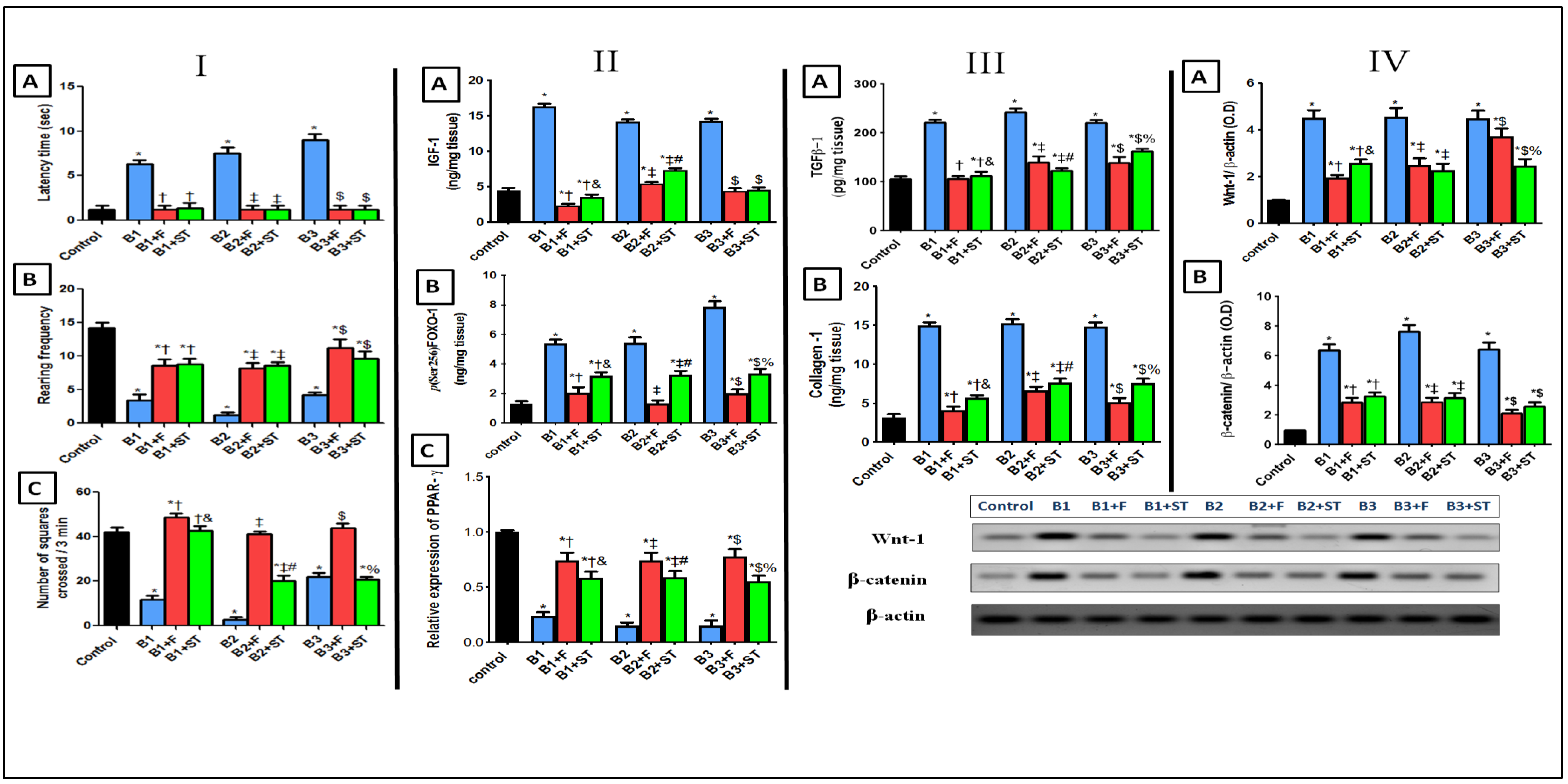
3.5.1. Rat Behavioral Alterations in Open Field Test
3.5.2. IGF-1/FOXO-1 and PPAR-γ Expression
3.5.3. TGFβ-1 and Collagen
3.5.4. Wnt-1/β-Catenin Expression
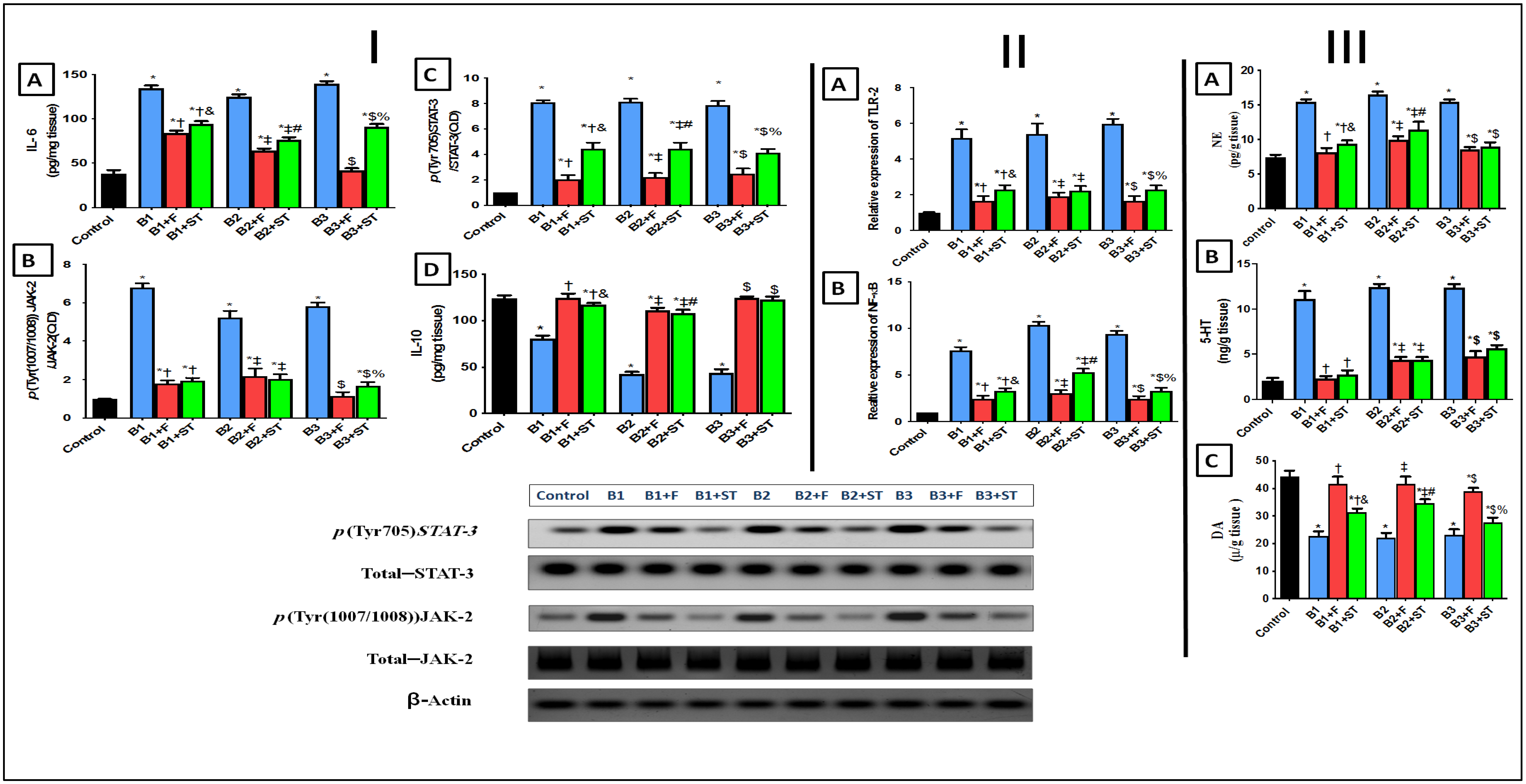
3.5.5. IL-6, IL-10, and p(Tyr (1007/1008))JAK-2/p(Tyr705)STAT-3 Signaling Pathway
3.5.6. TLR-2 and p(Ser536) NF-κBp65 Expression
3.5.7. Changes in Norepinephrine (NE), Serotonin (5-HT), and Dopamine (DA) Levels
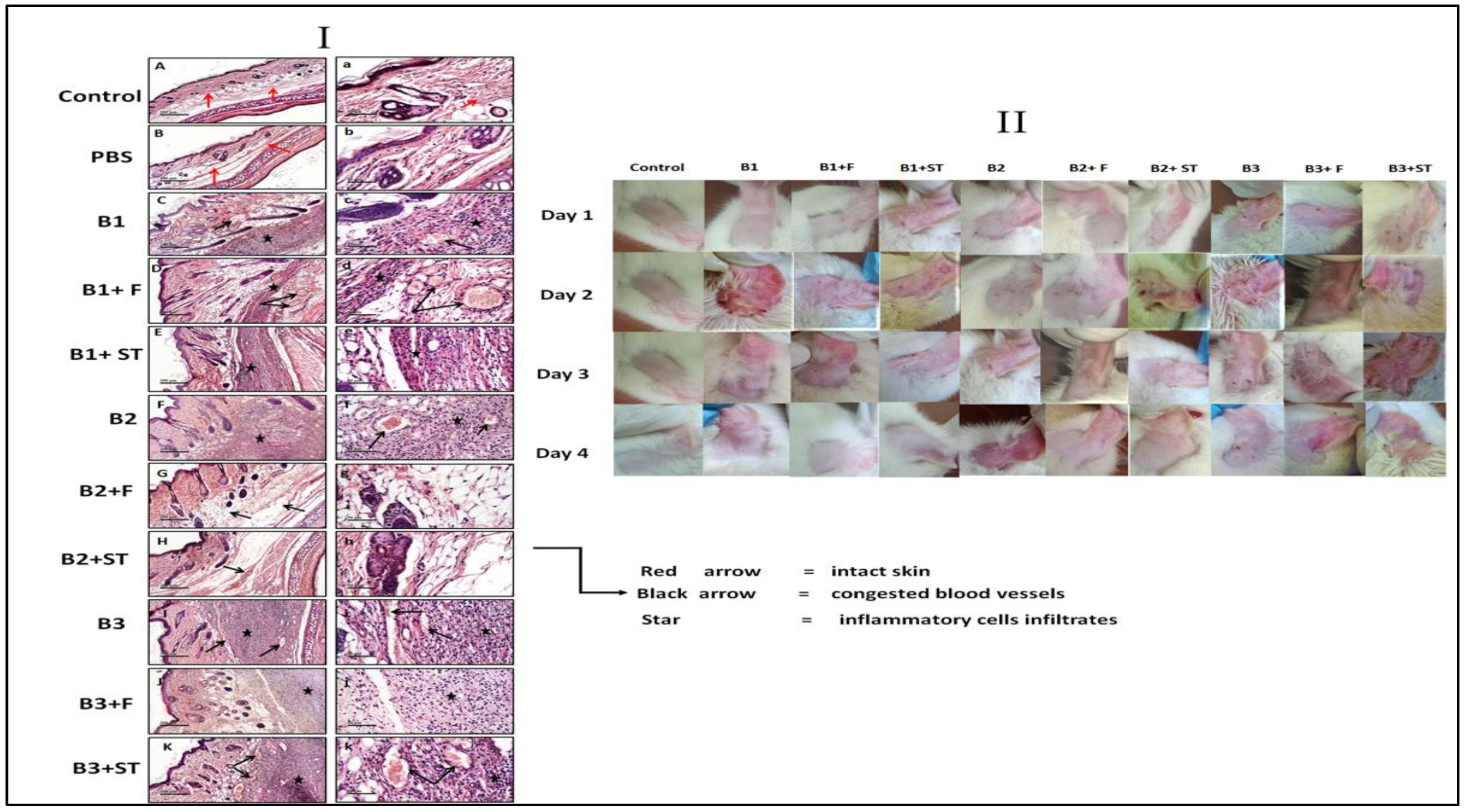
3.5.8. The Effect of NEG8 on Histopathology
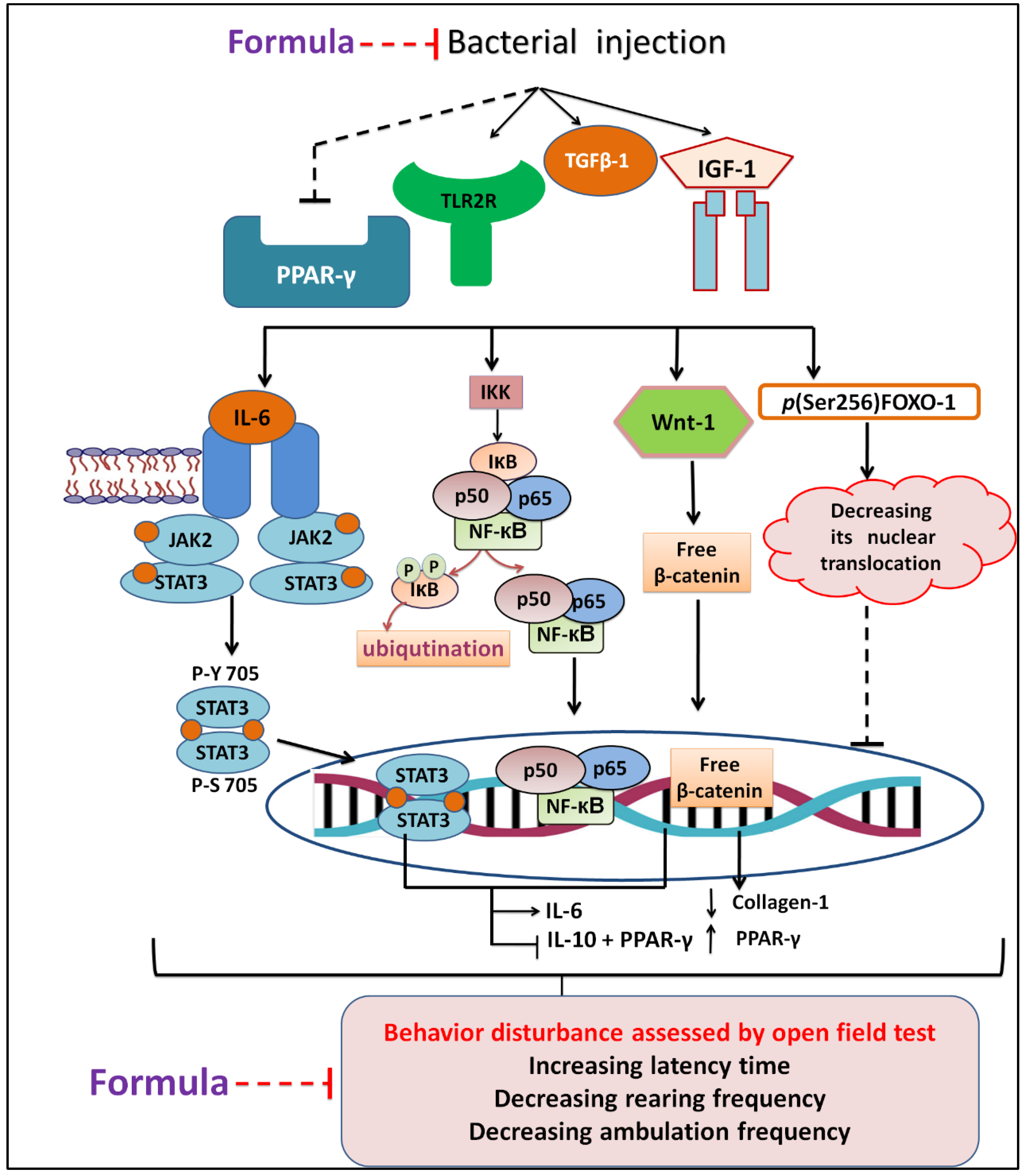
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviations
References
- Ledingham, K.; Hinchliffe, S.; Jackson, M.; Thomas, F.; Tomson, G.; World Health Organization. Antibiotic Resistance: Using a Cultural Contexts of Health Approach to Address a Global Health Challenge. Available online: https://apps.who.int/iris/handle/10665/330029 (accessed on 6 September 2021).
- León-Buitimea, A.; Garza-Cárdenas, C.R.; Garza-Cervantes, J.A.; Lerma-Escalera, J.A.; Morones-Ramírez, J.R. The demand for new antibiotics: Antimicrobial peptides, nanoparticles, and combinatorial therapies as future strategies in antibacterial agent design. Front Microbiol. 2020, 11, 1669. [Google Scholar] [CrossRef] [PubMed]
- Lagadinou, M.; Onisor, M.O.; Rigas, A.; Musetescu, D.-V.; Gkentzi, D.; Assimakopoulos, S.F.; Panos, G.; Marangos, M. Antimicrobial properties on non-antibiotic drugs in the era of increased bacterial resistance. Antibiotics 2020, 9, 107. [Google Scholar] [CrossRef] [Green Version]
- Monnet, D.L. Antibiotic development and the changing role of the pharmaceutical industry. Int. J. Risk Saf. Med. 2005, 1, 133–145. [Google Scholar]
- Saleem, M.; Nazir, M.; Ali, M.S.; Hussain, H.; Lee, Y.S.; Riaz, N.; Jabbar, A. ChemInform Abstract: Antimicrobial natural products: An update on future antibiotic drug candidates. ChemInform 2010, 41. [Google Scholar] [CrossRef]
- Naqvi, S.A.; Sana, N.; Sana, K.; Syed, A.N.; Muhammad, S.M.; Sajid, Y.Q.; Shahzad, A. Antioxidants: Natural antibiotics. In Antioxidants; IntechOpen: UK, London, 2019. [Google Scholar]
- Dreger, M.; Wielgus, K. Application of essential oils as natural cosmetic preservatives. Herba Pol. 2013, 59, 142–156. [Google Scholar] [CrossRef] [Green Version]
- McDaniel, A.; Tonyali, B.; Yucel, U.; Trinetta, V. Formulation, and development of lipid nanoparticle antifungal packaging films to control postharvest disease. J. Agric. Food Res. 2019, 1, 100013. [Google Scholar] [CrossRef]
- Bilia, A.R.; Guccione, C.; Isacchi, B.; Righeschi, C.; Firenzuoli, F.; Bergonzi, M.C. Essential oils loaded in nanosystems: A developing strategy for a successful therapeutic approach. Evidence-Based Complement. Altern. Med. 2014, 2014, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Daum, R.S.; Miller, L.G.; Immergluck, L.; Fritz, S.; Creech, C.; Young, D.; Kumar, N.; Downing, M.; Pettibone, S.; Hoagland, R.; et al. A placebo-controlled trial of antibiotics for smaller skin abscesses. N. Engl. J. Med. 2017, 376, 2545–2555. [Google Scholar] [CrossRef]
- Laube, S. Skin infections and ageing. Ageing Res. Rev. 2004, 3, 69–89. [Google Scholar] [CrossRef]
- Hassanzadeh, P.; Bahmani, M.; Mehrabani, D. Bacterial resistance to antibiotics in acne vulgaris: An in vitro study. Indian J. Dermatol. 2008, 53, 122. [Google Scholar] [CrossRef]
- Fabbrocini, G.; Cacciapuoti, S.; Monfrecola, G. A Qualitative investigation of the impact of Acne on health-related quality of life (HRQL): Development of a conceptual model. Dermatol. Ther. 2018, 8, 85–99. [Google Scholar] [CrossRef] [Green Version]
- Walsh, R.; Efthimiou, J.; Drén, B. Systematic review of antibiotic resistance in acne: An increasing topical and oral threat. Lancet Infect. Dis. 2016, 16, 23–33. [Google Scholar] [CrossRef] [Green Version]
- Williams, H.C.; Dellavalle, R.P.; Garner, S. Acne vulgaris. Lancet 2012, 28, 361–372. [Google Scholar] [CrossRef]
- Bueno, J.; Demirci, F.; Baser, K.H.C. Essential oils against microbial resistance mechanisms, challenges, and applications in drug discovery. In Essential Oils and Nanotechnology for Treatment of Microbial Diseases; CRC Press: Boca Raton, FL, USA, 2017; pp. 143–158. [Google Scholar]
- Yap, P.S.X.; Yiap, B.C.; Ping, H.C.; Lim, E. Essential oils, a new horizon in combating bacterial antibiotic resistance. Open Microbiol. J. 2014, 8, 6–14. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Olszowy, M.; Dawidowicz, A. Essential oils as antioxidants: Their evaluation by DPPH, ABTS, FRAP, CUPRAC, and β-carotene bleaching methods. Monatsh. Chem. 2016, 147, 2083–2091. [Google Scholar] [CrossRef]
- Robledo, N.; Vera, P.; López, L.; Yazdani-Pedram, M.; Tapia, C.; Abugoch, L. Thymol nanoemulsions incorporated in quinoa protein/chitosan edible films; antifungal effect in cherry tomatoes. Food Chem. 2018, 246, 211–219. [Google Scholar] [CrossRef]
- Cooper, K.E. Theory of antibiotic inhibition zones in agar media. Nature 1955, 176, 510–511. [Google Scholar] [CrossRef]
- Gehardt, P.; Murray, E.; Costilow, N. Manual of Methods for General Biotechnology; American society for microbiology: Washington, DC, USA, 1981. [Google Scholar]
- Lagacé, L.; Pitre, M.; Jacques, M.; Roy, D. Identification of the bacterial community of Maple sap by using amplified ribosomal DNA (rDNA) restriction analysis and rDNA sequencing. Appl. Environ. Microbiol. 2004, 70, 2052–2060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, F.; Gish, W.; Miller, W.; Myers, W.; Lipmanl, J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Thompson, D.; Higgins, G.; Gibson, J.; Clustal, W. Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 11, 4673–4780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, K.E.; Woodman, D. The diffusion of antiseptics through agar gels, with special reference to the agar cup assay method of estimating the activity of penicillin. J. Pathol. Bacteriol. 1946, 58, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Moideen, J.; Raffick, M. Antibacterial, antifungal activity and phytochemical analysis of Sanseviveria roxburghiana root. Int. J. Phytopharmacol. 2012, 3, 21–26. [Google Scholar]
- Stepanović, S.; Vukovic, D.; Hola, V.; Di Bonaventura, G.; Djukić, S.; Ćirković, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS 2007, 115, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Coenye, T.; Nelis, H.J. In vitro and in vivo model systems to study microbial biofilm formation. J. Microbiol. Methods 2010, 83, 89–105. [Google Scholar] [CrossRef] [PubMed]
- Han, R.; Blencke, H.M.; Cheng, H.; Li, C. The antimicrobial effect of CEN1HC-Br against Propionibacterium acnes and its therapeutic and anti-inflammatory effects on acne vulgaris. Peptides 2018, 1, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Malik, D.S.; Kaur, G. Exploring therapeutic potential of azelaic acid loaded NLCs for the treatment of acne vulgaris. J. Drug Deliv. Sci. Technol. 2019, 55, 101418. [Google Scholar] [CrossRef]
- Abd-Elhady, R.M.; Elsheikh, A.M.; Khalifa, A. Anti-amnestic properties of Ginkgo biloba extract on impaired memory function induced by aluminum in rats. Int. J. Dev. Neurosci. 2012, 31, 598–607. [Google Scholar] [CrossRef]
- Ji, J.; Zhang, R.; Li, H.; Zhu, J.; Pan, Y.; Guo, Q. Analgesic and anti-inflammatory effects and mechanism of action of borneol on photodynamic therapy of acne. Environ. Toxicol. Pharmacol. 2020, 75, 103329. [Google Scholar] [CrossRef]
- Sarkic, A.; Stappen, I. Essential oils and their single compounds in cosmetics—A critical review. Cosmetics 2018, 5, 11. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Zu, Y.; Chen, L.; Shi, X.; Wang, Z.; Sun, S.; Efferth, T. Antimicrobial activity of clove and rosemary essential oils alone and in combination. Phytother. Res. 2007, 21, 989–994. [Google Scholar] [CrossRef]
- Salehi, B.; Mishra, A.P.; Shukla, I.; Sharifi-Rad, M.; Contreras, M.; Segura-Carretero Sharifi-Rad, J. Thymol, thyme, and other plant sources: Health and potential uses. Phytother Res. 2018, 32, 1688–1706. [Google Scholar] [CrossRef] [PubMed]
- Torres-Martínez, R.; García-Rodríguez, Y.M.; Ríos-Chávez, P.; Saavedra-Molina, A.; López-Meza, J.E.; Ochoa-Zarzosa, A.; Garciglia, R.S. Antioxidant activity of the essential oil and its major terpenes of Satureja macrostema (Moc. and Sessé ex Benth.) Briq. Pharmacogn. Mag. 2018, 13, S875–S880. [Google Scholar] [CrossRef] [PubMed]
- Okur, M.E.; Ayla, Ş.; Yozgatlı, V.; Aksu, N.B.; Yoltaş, A.; Orak, D.; Sipahi, H.; Okur, N.Ü. Evaluation of burn wound healing activity of novel fusidic acid loaded microemulsion based gel in male wistar albino rats. Saudi. Pharm. J. 2020, 1, 338–348. [Google Scholar]
- Gurpret, K.; Singh, S.K. Review of nanoemulsion formulation and characterization techniques. Indian J. Pharm. Sci. 2018, 80, 781–789. [Google Scholar] [CrossRef]
- Abdullah, B.H.; Hatem, S.F.; Jumaa, W. A comparative study of the antibacterial activity of clove and rosemary essential oils on multidrug-resistant bacteria. UK. J. Pharm. Biosci. 2015, 3, 18–22. [Google Scholar] [CrossRef]
- Nzeako, B.C.; Al-Kharousi, Z.S.N.; Al-Mahrooqui, Z. Antimicrobial activities of clove and thyme extracts. Sultan Qaboos Univ. Med. J. 2006, 6, 33–39. [Google Scholar]
- Miklasińska-Majdanik, M.; Kępa, M.; Wojtyczka, R.D.; Idzik, D.; Wąsik, T.J. Phenolic compounds diminish antibiotic resistance of Staphylococcus aureus clinical strains. Int. J. Environ. Res. Public Health 2018, 15, 2321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srinivasan, R.; Santhakumari, S.; Poonguzhali, P.; Geetha, M.; Dyavaiah, M.; Xiangmin, L. Bacterial biofilm inhibition: A focused review on recent therapeutic strategies for combating the biofilm mediated infections. Front. Microbiol. 2021, 12, 1106. [Google Scholar] [CrossRef]
- Cao, Y.; Naseri, M.; He, Y.; Xu, C.; Walsh, L.J.; Ziora, Z.M. Non-antibiotic antimicrobial agents to combat bio-film-forming bacteria. J. Glob. Antimicrob. Resist. 2020, 21, 445–451. [Google Scholar] [CrossRef]
- Cong, T.-X.; Hao, D.; Wen, X.; Li, X.-H.; He, G.; Jiang, X. From pathogenesis of acne vulgaris to anti-acne agents. Arch. Dermatol. Res. 2019, 311, 337–349. [Google Scholar] [CrossRef]
- Sertznig, P.; Reichrath, J. Peroxisome proliferator-activated receptors (PPARs) in dermatology: Challenge and promise. Dermato-Endocrinol. 2011, 1, 130–135. [Google Scholar] [CrossRef]
- Trivedi, R.; Cong, Z.; Nelson, M.; Albert, J.; Rosamilia, L.; Sivarajah, S.; Thiboutot, M. Peroxisome prolifera-tor-activated receptors increase human sebum production. J. Investig. Dermatol. 2006, 1, 2002–2009. [Google Scholar] [CrossRef] [Green Version]
- Fan, W.; Imamura, T.; Sonoda, N.; Sears, D.; Patsouris, D.; Kim, J. FOXO1 transrepresses peroxisome prolifera-tor-activated receptor γ transactivation, coordinating an insulin-induced feed-forward response in adipocytes. J. Biol. Chem. 2009, 284, 12188–12197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arden, K.C.; Biggs, W., 3rd. Regulation of the FoxO family of transcription factors by phosphatidylinositol-3 ki-nase-activated signaling. Arch. Biochem. Biophys. 2002, 403, 292. [Google Scholar] [CrossRef]
- Agak, G.W.; Qin, M.; Nobe, J.; Kim, M.-H.; Krutzik, S.R.; Tristan, G.R.; Elashoff, D.; Garbán, H.J.; Kim, J. Propionibacterium acnes induces an IL-17 response in Acne vulgaris that is regulated by vitamin A and vitamin D. J. Investig. Dermatol. 2014, 134, 366–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Depino, A.M.; Lucchina, L.; Pitossi, F. Early and adult hippocampal TGF-β1 overexpression have opposite effects on behavior. Brain Behav. Immun. 2011, 25, 1582–1591. [Google Scholar] [CrossRef]
- Logan, C.Y.; Nusse, R. The WNT signaling pathway in development and disease. Annu. Rev. Cell Dev. Biol. 2004, 20, 781–810. [Google Scholar] [CrossRef] [Green Version]
- Vallée, A.; Lecarpentier, Y.; Guillevin, R.; Vallée, N. Interactions between TGF-β1, canonical WNT/β-catenin pathway and PPAR γ in radiation-induced fibrosis. Oncotarget 2017, 27, 90579. [Google Scholar] [CrossRef] [Green Version]
- O’Shea, J.J.; Murray, P.J. Cytokine signaling modules in inflammatory responses. Immunity 2008, 28, 477–487. [Google Scholar] [CrossRef] [Green Version]
- Sharma, M.; Schoop, R.; Suter, A.; Hudson, J.B. The potential use of Echinacea in acne: Control of Propionibacterium acnes growth and inflammation. Phytother. Res. 2010, 25, 517–521. [Google Scholar] [CrossRef] [PubMed]
- Titus, S.; Hodge, J. Diagnosis and treatment of acne. Am. Fam. Physician 2012, 86, 734–740. [Google Scholar] [PubMed]
- Kim, J.; Ochoa, M.T.; Krutzik, S.R.; Takeuchi, O.; Uematsu, S.; Legaspi, A.J.; Brightbill, H.D. Activation of toll-like receptor 2 in acne triggers inflammatory cytokine responses. J. Immunol. 2002, 1, 1535–1541. [Google Scholar] [CrossRef] [Green Version]
- Joo, S.S.; Kil Jang, S.; Kim, S.G.; Choi, J.-S.; Hwang, K.W.; Lee, D.I. Anti-acne activity of Selaginella involvens extract and its non-antibiotic antimicrobial potential on Propionibacterium acnes. Phytotherapy Res. 2008, 22, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Koistinaho, J.; Malm, T.; Goldsteins, G. Glycogen synthase kinase-3β: A mediator of inflammation in Alzheimer’s disease? Int. J. Alzheimers Dis. 2011, 2011, 129753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thierry, A.-M.; Fekete, M.; Glowinski, J. Effects of stress on the metabolism of noradrenaline, dopamine and serotonin (5HT) in the central nervous system of the rat (II) modifications of serotonin metabolism. Eur. J. Pharmacol. 1968, 4, 384–389. [Google Scholar] [CrossRef]
- Corrodi, H.; Fuxe, K.; Hökfelt, T. A Possible role played by central monoamine neurones in thermo-regulation. Acta Physiol. Scand. 1967, 71, 224–232. [Google Scholar] [CrossRef]
- Raison, C.L.; Capuron, L.; Miller, A.H. Cytokines sing the blues: Inflammation and the pathogenesis of depression. Trends Immunol. 2006, 27, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Yap, P.S.; Yang, S.K.; Lai, K.S.; Lim, S.H. Essential oils: The ultimate solution to antimicrobial resistance in Escherichia coli. In Escherichia coli: Recent Advances on Physiology, Pathogenesis and Biotechnological Applications; IntechOpen: London, UK, 2017; p. 15. [Google Scholar]
- Yang, S.-K.; Tan, N.-P.; Chong, C.-W.; Abushelaibi, A.; Lim, S.-H.; Lai, K.-S. The missing piece: Recent approaches investigating the antimicrobial mode of action of essential oils. Evol. Bioinform. 2021, 17. [Google Scholar] [CrossRef]
- El-Tarabily, K.A.; El-Saadony, M.T.; Alagawany, M.; Arif, M.; Batiha, G.E.; Khafaga, A.F.; Elwan, H.A.; Elnesr, S.S.; El-Hack, M.E.A. Using essential oils to overcome bacterial biofilm formation and their antimicrobial resistance. Saudi J. Biol. Sci. 2021, 28, 5145–5156. [Google Scholar] [CrossRef]
- Jesus, G.S.; Micheletti, A.C.; Takahashi, K.M.; Matayoshi, T.; Pott, A.; Yoshida, N.C. Antimicrobial potential of Pectis substriata essential oil (Asteraceae) against drug-resistant Staphylococcus strains. An Acad Bras Cienc. 2020, 14, 92. [Google Scholar]
- Pazyar, N.; Yaghoobi, R.; Bagherani, N.; Kazerouni, A. A review of applications of tea tree oil in dermatology. Int. J. Dermatol. 2012, 52, 784–790. [Google Scholar] [CrossRef]
- Li, Z.-H.; Cai, M.; Liu, Y.-S.; Sun, P.-L.; Luo, S.-L. Antibacterial activity, and mechanisms of essential oil from Citrus medica L. var. sarcodactylis. Molecules 2019, 24, 1577. [Google Scholar] [CrossRef] [Green Version]
- Munir, M.U.; Ahmed, A.; Usman, M.; Salman, S. Recent advances in nanotechnology-aided materials in combating microbial resistance and functioning as antibiotics substitutes. Int. J. Nanomedicine. 2020, 15, 7329. [Google Scholar] [CrossRef]
- Rai, M.; Paralikar, P.; Jogee, P.; Agarkar, G.; Ingle, A.P.; Derita, M.; Zacchino, S. Synergistic antimicrobial potential of essential oils in combination with nanoparticles: Emerging trends and future perspectives. Int. J. Pharm. 2017, 519, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Taleb, M.H.; Abdeltawab, N.F.; Shamma, R.N.; Abdelgayed, S.S.; Mohamed, S.S.; Farag, M.A.; Ramadan, M.A. Origanum vulgare L. essential oil as a potential anti-acne topical nanoemulsion—In vitro and in vivo study. Molecules 2018, 23, 2164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamil, B.; Abbasi, R.; Abbasi, S.; Imran, M.; Khan, S.U.; Ihsan, A.; Javed, S.; Bokhari, H. Encapsulation of Cardamom essential oil in chitosan nanocomposites: In-vitro Efficacy on antibiotic-resistant bacterial pathogens and cytotoxicity studies. Front. Microbiol. 2016, 7, 1580. [Google Scholar] [CrossRef] [PubMed]
- Swamy, M.K.; Akhtar, M.S.; Sinniah, U.R. Antimicrobial properties of plant essential oils against human pathogens and their mode of action: An updated review. Evid. Based Complement. Altern. Med. 2016, 2016, 1–21. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Surf. Ratio | Formula Code | Oil | Surfactants | Water | Visual Appearance | |
|---|---|---|---|---|---|---|
| Tween 80 | Cremophor RH40 | |||||
| (1:1) | NEG1 | 10 | 30 | 30 | 30 | turbid |
| NEG2 | 20 | 30 | 30 | 20 | gel | |
| NEG3 | 30 | 30 | 30 | 10 | fluid | |
| (2:1) | NEG4 | 10 | 40 | 20 | 30 | turbid |
| NEG5 | 20 | 40 | 20 | 20 | gel | |
| NEG6 | 30 | 40 | 20 | 10 | fluid | |
| (3:1) | NEG7 | 10 | 45 | 15 | 30 | turbid |
| NEG8 | 20 | 45 | 15 | 20 | gel | |
| NEG9 | 30 | 45 | 15 | 10 | fluid | |
| Tested Oil | Major Area % | Rt | M.wt | Molecular Formula | Compound | Fragmentation Pattern |
|---|---|---|---|---|---|---|
| Garlic | 40.87 | 12.17 | 178 | C6H10S3 | Allyl trisulfide | 39, 41, 73, 113, 178 |
| Eucalyptus | 48.88 | 8.65 | 152 | C10H16O | 2-Bornanone | 41, 55, 69, 81, 95, 108, 152 |
| Lavender | 39.39 | 7.41 | 154 | C10H18O | L-Linalool | 55, 69, 71, 80, 93, 121, 136 |
| Cinnamon | 60.97 | 11.73 | 132 | C9H8O | Cinnamaldehyde | 51, 77, 78, 103, 131, 132 |
| Thyme | 56.62 * | 11.85 | 150 | C10H14O | Thymol | 77, 91, 115, 117, 135, 150 |
| Lemongrass | 52.46 | 5.86 | 136 | C10H16 | D-Limonene | 41, 53, 67, 93, 107, 121, 136 |
| Cumin | 34.77 | 6.47 | 136 | C10H16 | Terpinene | 41, 77, 79, 91, 93, 105, 121, 136 |
| Geranium | 36.27 | 10.3 | 156 | C10H20O | Citronellol | 41, 55, 69, 82, 95, 109, 123, 138, 156 |
| Clove | 94.13 * | 13.25 | 164 | C10H12O2 | Eugenol | 55, 77, 91, 103, 121, 131, 149, 164 |
| Peppermint | 23.90 | 9.26 | 156 | C10H20O | Menthanol | 41, 43, 55, 71, 81, 95, 123, 138, 156 |
| Formula Code | PS (nm) | PDI | ZP (mV) |
|---|---|---|---|
| NEG2 | 52.44 ± 3.22 | 0.413 ± 0.00 | −12.40 ± 0.70 |
| NEG5 | 36.43 ± 1.63 a | 0.260 ± 0.06 a | −12.50 ± 2.26 |
| NEG8 | 33.46 ± 1.31 a | 0.315 ± 0.05 | −14.2 ± 2.31 |
| NEG8 (6 months storage) | 29.49 ± 2.51 a,b | 0.42 ± 0.06 b | −19.43 ± 2.10 a,b,c |
| 1 | BXYL | - | 3 | LysA | - | 4 | AspA | - | 5 | LeuA | + | 7 | PheA | + | 8 | ProA | - |
| 9 | BGAL | - | 10 | PyrA | + | 11 | AGAL | - | 12 | AlaA | - | 13 | TyrA | - | 14 | BNAG | + |
| 15 | APPA | + | 18 | CDEX | - | 19 | dGAL | - | 21 | GLYG | - | 22 | INO | - | 24 | MdG | - |
| 25 | ELLM | - | 26 | MdX | - | 27 | AMAN | - | 29 | MTE | - | 30 | GlyA | - | 31 | dMAN | - |
| 32 | dMNE | - | 34 | dMLZ | - | 36 | NAG | + | 37 | PLE | - | 39 | IRHA | - | 41 | BGLU | + |
| 43 | BMAN | - | 44 | PHC | - | 45 | PVATE | + | 46 | AGLU | - | 47 | dTAG | - | 48 | dTRE | + |
| 50 | INU | - | 53 | dGLU | + | 54 | dRIB | + | 56 | PSCNa | - | 58 | NaCl | + | 59 | KAN | - |
| 60 | OLD | - | 61 | ESC | + | 62 | TIZ | - | 63 | POLYB | + | 1 |
| Bacterial Name | Zone of Inhibition | MIC | MBC |
|---|---|---|---|
| Pseudomonas stutzeri (B1) | 19 | 1.25 | 5.0 |
| Enterococcus faecium (B2) | 25 | 0.62 | 2.5 |
| Bacillus thuringiensis (B3) | 23 | 1.25 | 5.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nazeam, J.A.; Ragab, G.M.; El-Gazar, A.A.; El-Mancy, S.S.; Jamil, L.; Fayez, S.M. Topical Nano Clove/Thyme Gel against Genetically Identified Clinical Skin Isolates: In Vivo Targeting Behavioral Alteration and IGF-1/pFOXO-1/PPAR γ Cues. Molecules 2021, 26, 5608. https://doi.org/10.3390/molecules26185608
Nazeam JA, Ragab GM, El-Gazar AA, El-Mancy SS, Jamil L, Fayez SM. Topical Nano Clove/Thyme Gel against Genetically Identified Clinical Skin Isolates: In Vivo Targeting Behavioral Alteration and IGF-1/pFOXO-1/PPAR γ Cues. Molecules. 2021; 26(18):5608. https://doi.org/10.3390/molecules26185608
Chicago/Turabian StyleNazeam, Jilan A., Ghada M. Ragab, Amira A. El-Gazar, Shereen S. El-Mancy, Lina Jamil, and Sahar M. Fayez. 2021. "Topical Nano Clove/Thyme Gel against Genetically Identified Clinical Skin Isolates: In Vivo Targeting Behavioral Alteration and IGF-1/pFOXO-1/PPAR γ Cues" Molecules 26, no. 18: 5608. https://doi.org/10.3390/molecules26185608
APA StyleNazeam, J. A., Ragab, G. M., El-Gazar, A. A., El-Mancy, S. S., Jamil, L., & Fayez, S. M. (2021). Topical Nano Clove/Thyme Gel against Genetically Identified Clinical Skin Isolates: In Vivo Targeting Behavioral Alteration and IGF-1/pFOXO-1/PPAR γ Cues. Molecules, 26(18), 5608. https://doi.org/10.3390/molecules26185608