
Contribution of the Adenosine 2A Receptor to Behavioral Effects of Tetrahydrocannabinol, Cannabidiol and PECS-101


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
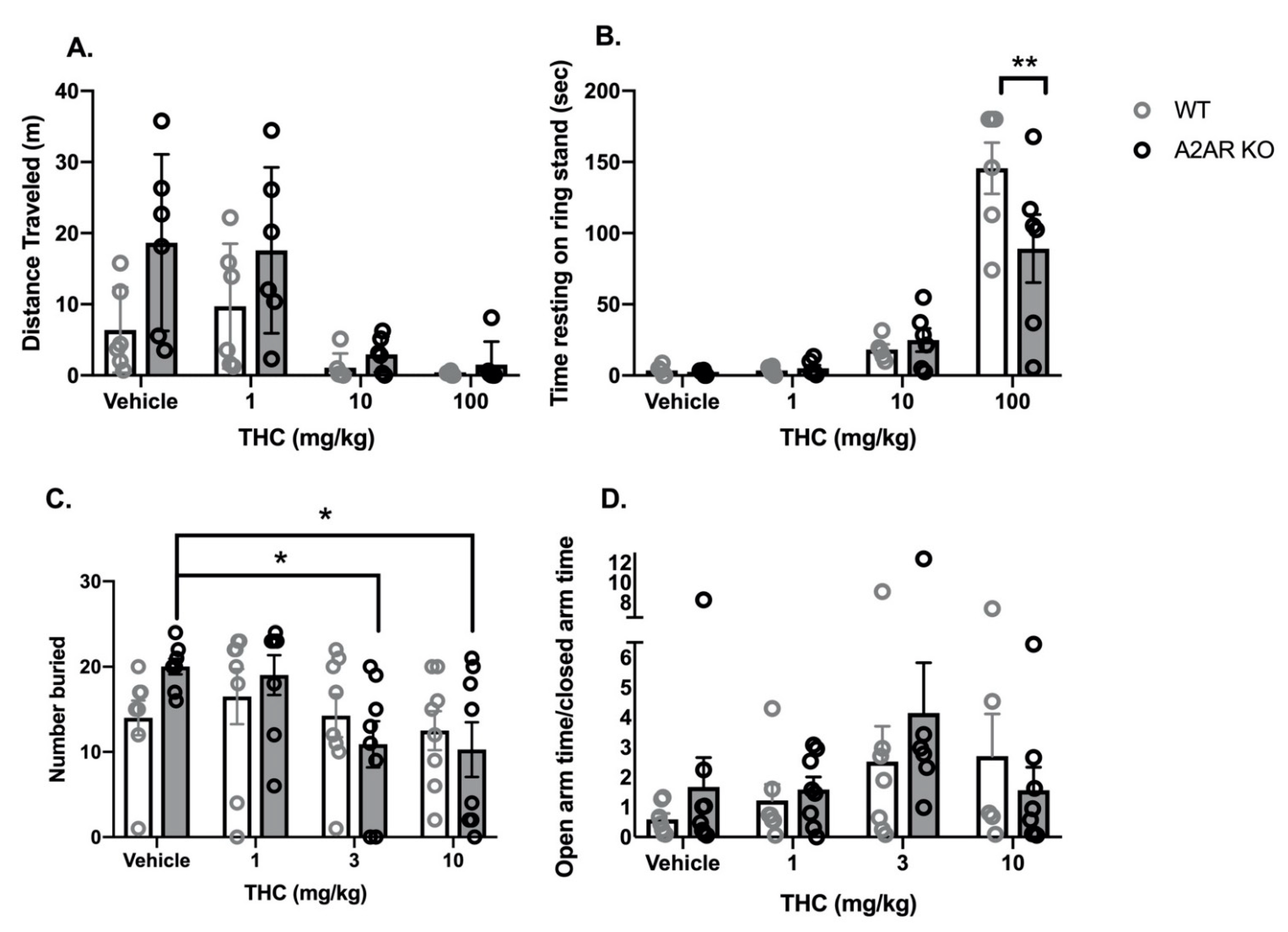
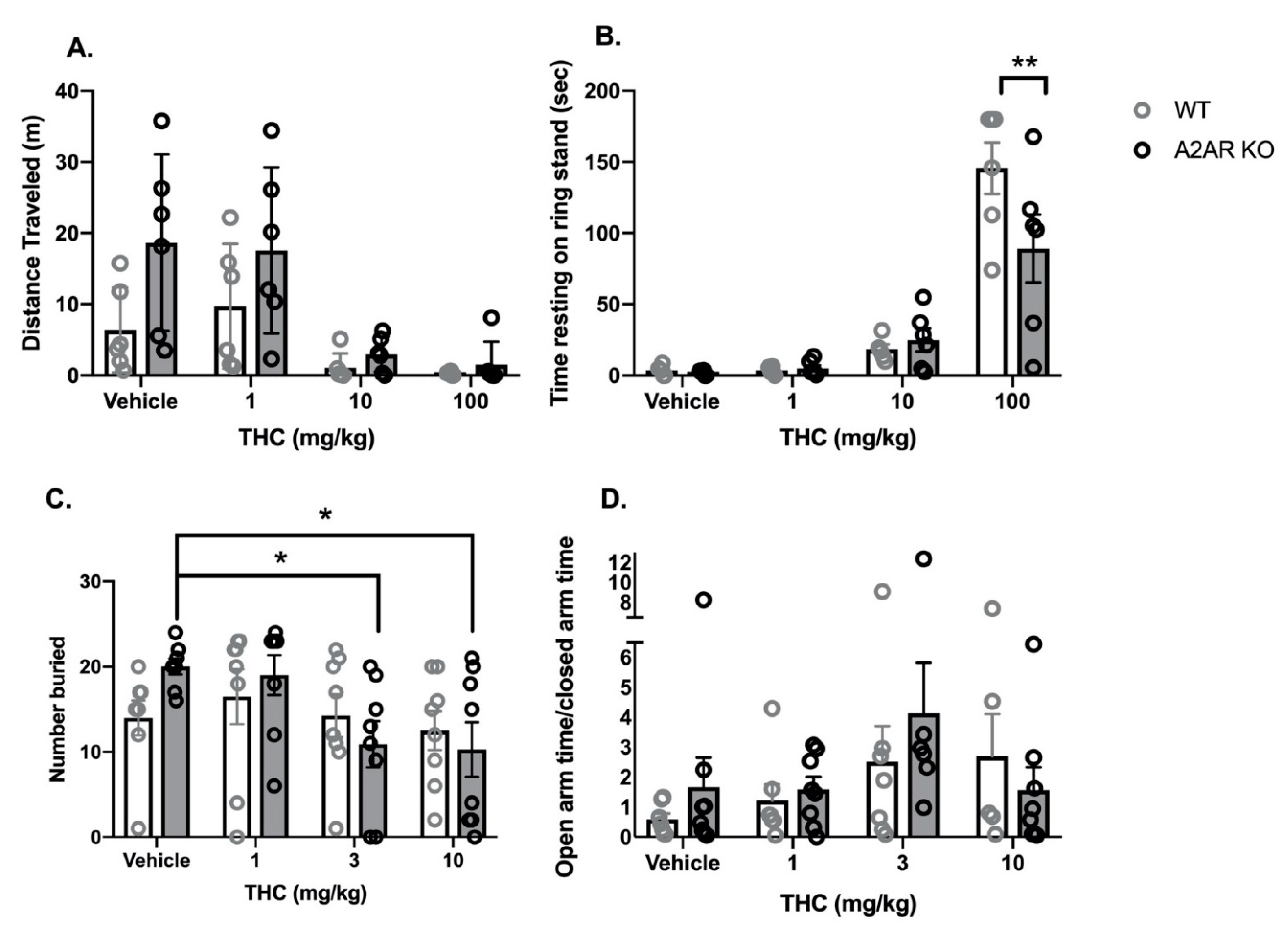
2.1. THC Dose Response Studies
2.1.1. Cannabinoid Tetrad
- Locomotor activity: Vehicle-treated, A2AR-KO mice exhibited a greater mean distance traveled in the open field than vehicle-treated, WT mice (Figure 1A). THC produced a dose-dependent reduction in locomotor activity in both WT and A2AR-KO mice. Two-way ANOVA indicated significant effects of both THC (F3,40 = 10.3, p < 0.0001) and genotype (F1,40 = 7.7, p < 0.01) without a significant interaction (F3,40 = 1.5, p = 0.22).
- Catalepsy: THC produced a dose-dependent increase in the time with front paws on a ring stand, a commonly employed assay for cannabinoid-induced catalepsy (Figure 1B). Two-way ANOVA indicated a significant effect of THC (F3,40 = 47.4, p < 0.0001) and a significant interaction between THC and genotype (F3,40 = 3.4, p < 0.05) without a significant effect of genotype alone (F1,40 = 2.4, p = 0.13). Sidak’s multiple comparison post hoc test revealed that, following treatment with 100 mg/kg THC, A2AR-KO mice exhibited significantly less catalepsy than WT.
- Body temperature: Rectal temperature was measured as an index of body temperature (Supplementary Materials Figure S1A). Two-way ANOVA indicated a significant effect of THC treatment (F3,40 = 35.0, p < 0.0001); genotype did not significantly affect rectal temperature (F1,40 = 0.9, p = 0.33), and the interaction was not significant (F3,40 = 0.3, p = 0.82).
- Nociceptive reflex: Latency to move the tail in response to a heat stimulus was used to assess the antinociceptive effects of THC in both genotypes (Supplementary Materials Figure S1B). Two-way ANOVA indicated a significant effect of THC (F3,40 = 17.0, p < 0.0001); genotype did not significantly affect the tail-flick latency (F1,40 = 0.9, p = 0.35) and the interaction was not significant (F3,40 = 1.3, p = 0.30).
2.1.2. Anxiety Assays
- Marble Bury Assay: Two-way ANOVA indicated a significant effect of THC (F3,56 = 3.2, p < 0.05) without a significant effect of genotype (F1,56 = 0.16, p = 0.88) or a significant interaction (F3,56 = 1.5, p = 0.41) (Figure 1C). Dunnett’s t-tests indicate that treatment with 3 and 10 mg/kg THC significantly reduces the number of marbles buried compared to vehicle treated in the A2AR-KO mice; there were no significant differences in the WT mice.
- EPM: Two-way ANOVA indicated no significant effects of either THC (F3,48 = 2.15, p = 0.1) or genotype (F1,48 = 0.52, p = 0.47) and no significant interaction (F3,48 = 0.74, p = 0.53) on the ratio of time spent in the open and closed arms (OAT/CAT) (Figure 1D). There were no significant effects of THC (F3,48 = 0.30, p = 0.73) or genotype (F3,48 = 0.05, p = 0.26) on the total number of arm entries (Supplementary Materials Figure S1D).
2.2. THC/CBD Combination Studies
- Locomotor Activity: The distance traveled in the open field was measured in the eight treatment groups (Figure 2A). Three-way ANOVA indicated significant effects of both THC (F1,55 = 13.0, p < 0.001) and genotype (F1,55 = 6.3, p < 0.05) but not CBD (F1,55 = 0.025, p = 0.87). There was a significant interaction between THC and genotype (F1,55 = 6.6, p < 0.05). Post hoc tests revealed that THC produced a significant reduction in locomotor activity in A2AR-KO mice also treated with CBD.
- Catalepsy: Three-way ANOVA indicated significant effects of THC (F1,52 = 20.0, p < 0.0001) and CBD (F1,52 = 5.0, p < 0.05) and a trend toward a significant effect of genotype (F1,52 = 2.4, p = 0.13) (Figure 2B). There was a significant interaction between CBD and genotype (F1,52 = 5.3, p < 0.05). Post hoc tests demonstrate that WT mice treated with a combination of THC and CBD exhibited significantly greater time resting on the ring stand than WT mice treated with either drug alone. Neither THC nor CBD or their combination produced catalepsy in A2AR-KO mice. Post hoc tests revealed a significant difference between the THC/CBD cotreatment groups in WT and A2AR-KO mice.
- Body Temperature: Three-way ANOVA indicates a significant effect of THC (F1,56 = 4.26, p < 0.05) without significant effects of either CBD (F1,56 = 0.9, p = 0.35) or genotype (F1,56 = 2.6, p = 0.11) (Supplementary Materials Figure S2A).
- Nociceptive Reflex: Three-way ANOVA indicates a significant effect of THC (F1,56 = 19, p < 0.001) without significant effects of either CBD (F1,56 = 0.8, p = 0.79) or genotype (F1,56 = 0.3, p = 0.58) (Supplementary Materials Figure S2B).
- Marble Bury Assay. Three-way ANOVA indicates significant effects of both THC (F1,56 = 10.4, p < 0.01) and CBD (F1,56 = 6.2, p < 0.05) but not genotype (F1,56 = 0.55, p = 0.46) (Figure 2C). The interaction between CBD and genotype is significant (F1,56 = 7.1, p < 0.01); post hoc tests demonstrated a significant reduction in the number of marbles buried between the vehicle treated and THC/CBD treated A2AR-KO mice.
- EPM: Three-way ANOVA indicated that both THC (F1,48 = 4.9, p < 0.05) and CBD (F1,48 = 5.4, p < 0.05) had significant effects on the OAT/CAT ratio, while genotype trended to a significant effect (F1,48 = 2.9, p = 0.09) (Figure 2D). The interaction between genotype and CBD was also significant (F1,48 = 4.1, p < 0.05), while the interaction between CBD and THC trended toward significance (F1,48 = 2.6, p = 0.1). Post hoc tests revealed a significant increase in OAT/CAT in WT mice treated with a combination of THC/CBD compared to those treated with vehicle and those treated with THC alone. Three-way ANOVA of the total arm entries indicated no significant effects of either THC (F1,48 = 0.5, p = 0.49) or CBD (F1,48 = 1.1, p = 0.31) (Supplementary Materials Figure S2C). While there was a significant effect of genotype (F1,48 = 5.0, p < 0.05) and a significant interaction between THC and genotype (F1,48 = 5.9, p < 0.05), post hoc tests did not elucidate any significant group differences.
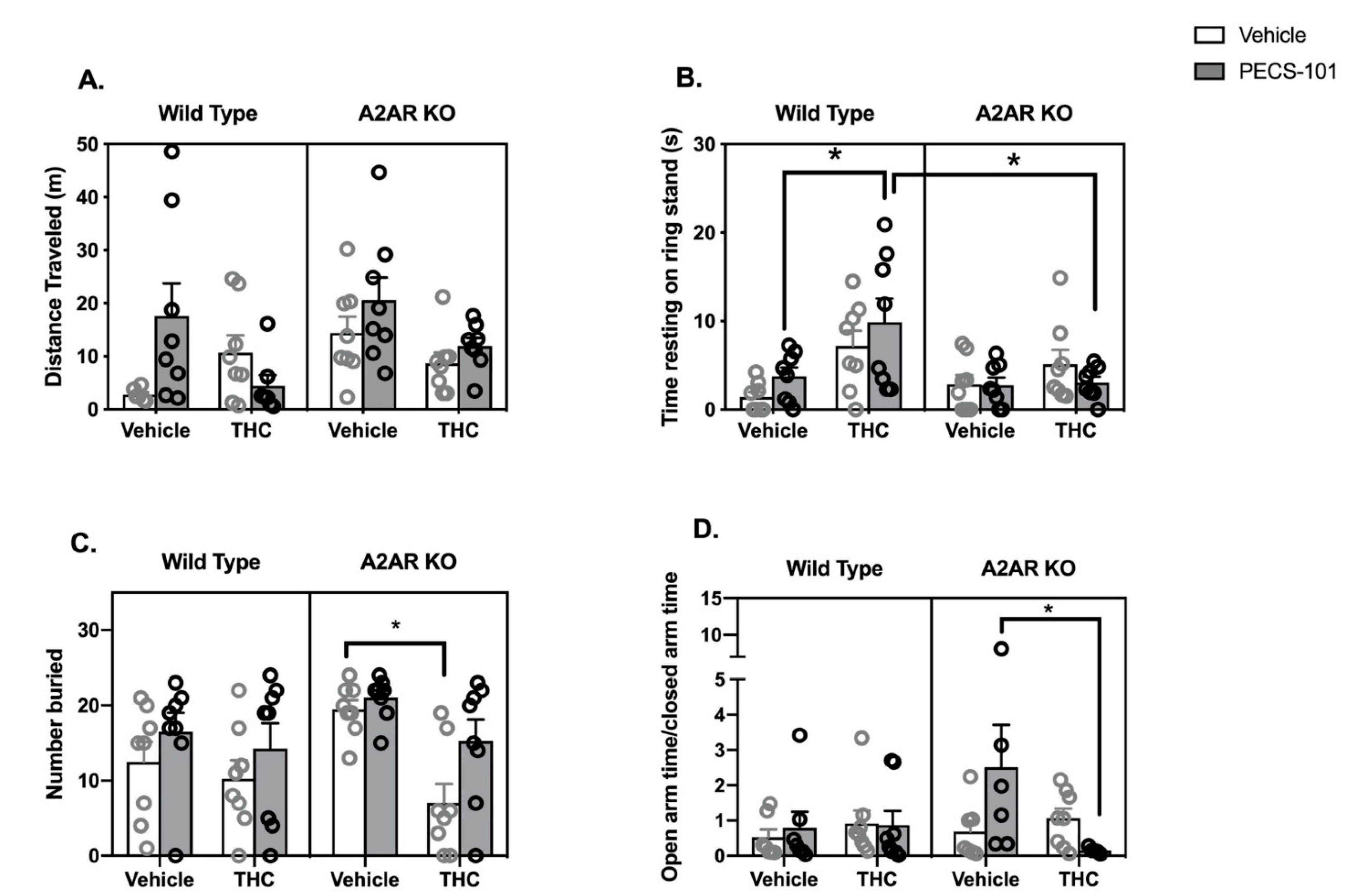
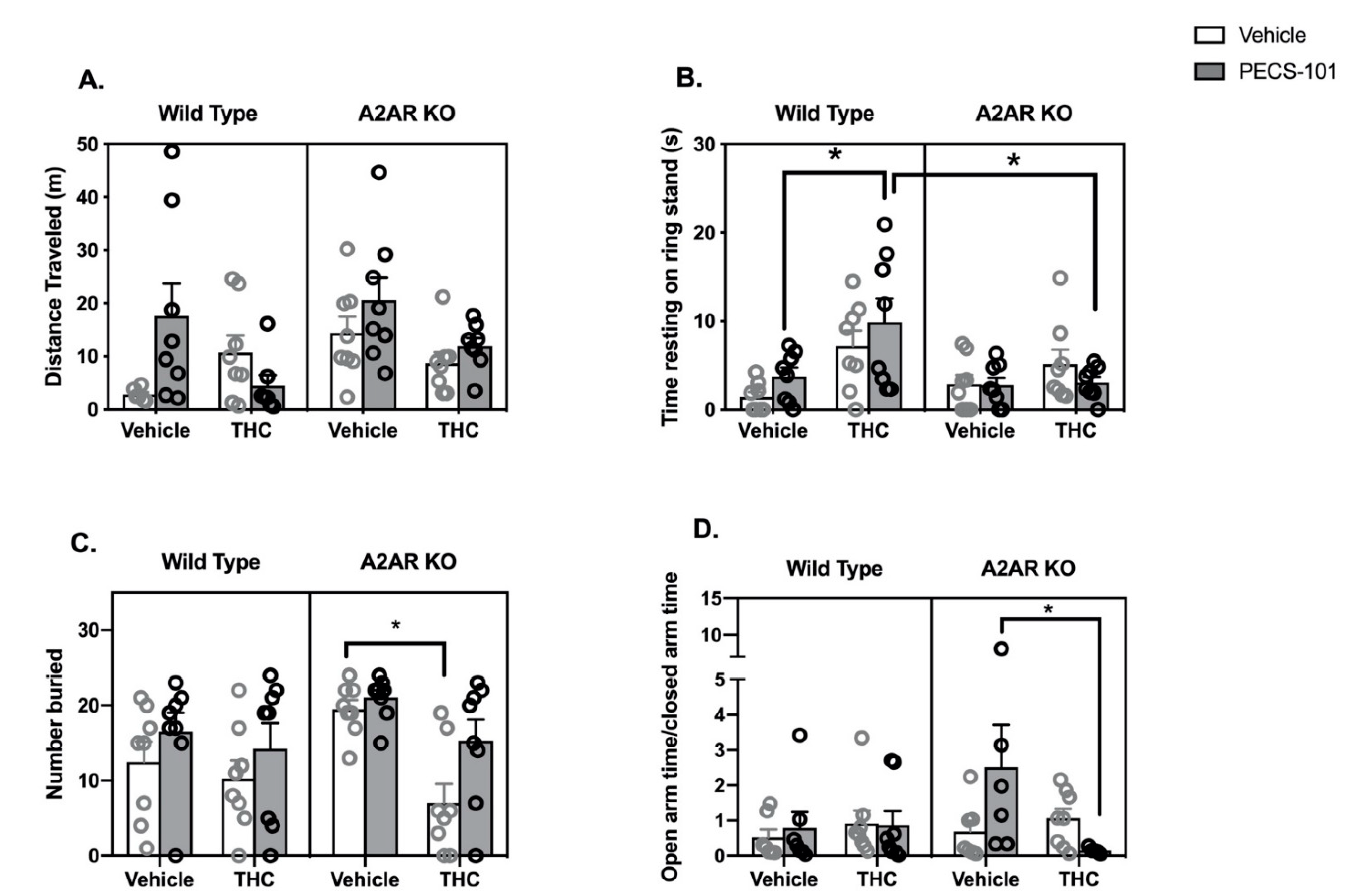
2.3. THC/PECS-101 Combination Studies
- Locomotor Activity: Three-way ANOVA indicated a significant effect of genotype (F1,53 = 4.2, p < 0.05), while the effects of both THC (F1,53 = 4.0, p = 0.051) and PECS-101 trended to significance (F1,53 = 3.4, p = 0.07) (Figure 3A). There was a significant interaction between THC and PECS-101 (F1,53 = 6.0, p < 0.05) and a trend to an interaction among THC and PECS-101 and genotype (F1,53 = 3.4, p = 0.07).
- Catalepsy: Three-way ANOVA indicated significant effects of THC (F1,56 = 12.8, p < 0.001) and genotype (F1,56 = 4.3, p < 0.05) but not PECS-101 (F1,56 = 0.5, p = 0.48) (Figure 3B). There was a significant interaction between THC and genotype (F1,56 = 5.3, p < 0.05). Post hoc tests demonstrate that WT mice treated with a combination of THC and PECS-101 exhibited significantly greater time resting on the ring stand than WT mice treated with PECS-101 alone. However, this did not occur in A2AR-KO mice, and there was a significant difference between the WT and A2AR-KO THC/PECS-101 combined treatment groups.
- Body Temperature: Three-way ANOVA indicates a significant effect of THC (F1,56 = 4.1, p < 0.05) without significant effects of either PECS-101 (F1,56 = 0.5, p = 0.97) or genotype (F1,56 = 1.9, p = 0.17) (Supplementary Materials Figure S3A).
- Nociceptive Reflex: Three-way ANOVA indicates significant effects of THC (F1,56 = 8.3, p < 0.01) and PECS-101 (F1,56 = 10.8, p < 0.01) but not genotype (F1,56 = 0.3, p = 0.87) (Supplementary Materials Figure S3B). The examination of the data suggest that THC inhibits the nociceptive reflex in the absence but not in the presence of PECS-101, although the interaction of THC and PECS-101 was not significant (p = 0.15).
- Marble Bury Assay: Three-way ANOVA indicates significant effects of THC (F1,56 = 10.8, p < 0.01) and PECS-101 (F1,56 = 6.6, p < 0.05) but not genotype (F1,56 = 1.8, p = 0.19) (Figure 3C). There was a nearly significant interaction between THC and genotype (F1,56 = 3.9, p = 0.052). Post hoc tests demonstrated that, in the A2AR-KO mice only, THC reduced the number of marbles buried compared to both vehicle and PECS-101 treated mice.
- EPM: Three-way ANOVA indicates that none of the factors had a significant effect on the OAT/CAT ratio (THC: F1,48 = 1.2, p = 0.28; PECS-101: F1,48 = 0.7, p = 0.42; and genotype: F1,48 = 1.0, p = 0.34) (Figure 3D). There was, however, a significant interaction between THC and PECS-101 (F1,48 = 4.8, p < 0.05) and a trending interaction among THC, PECS-101 and genotype (F1,48 = 3.0, p = 0.09). Post hoc tests indicated that the combination of THC and PECS-101 significantly reduced the OAT/CAT ratio compared to the effect of PECS-101 alone in the A2AR-KO mice. Surprisingly, PECS-101 had a very significant effect on the total arm entries; the three-way ANOVA results for PECS-101 were F1,48 = 21, p < 0.0001; neither THC (F1,48 = 2.1, p = 0.16) nor genotype (F1,48 = 0.5, p = 0.46) significantly affected the total arm entries (Supplementary Materials Figure S3C).
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drugs
4.3. Behavioral Assays
4.4. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Dos Santos, R.G.; Hallak, J.E.C.; Crippa, J.A.S. Neuropharmacological Effects of the Main Phytocannabinoids: A Narrative Review. Adv. Exp. Med. Biol. 2021, 1264, 29–45. [Google Scholar] [PubMed]
- Britch, S.C.; Babalonis, S.; Walsh, S.L. Cannabidiol: Pharmacology and therapeutic targets. Psychopharmacology 2021, 238, 9–28. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Ruiz, J.; Galve-Roperh, I.; Sagredo, O.; Guzman, M. Possible therapeutic applications of cannabis in the neuropsychopharmacology field. Eur. Neuropsychopharmacol. 2020, 36, 217–234. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, S.E.; Stevenson, C.W.; Laviolette, S.R. Could Cannabidiol Be a Treatment for Coronavirus Disease-19-Related Anxiety Disorders? Cannabis Cannabinoid Res. 2021, 6, 7–18. [Google Scholar] [CrossRef]
- Mechoulam, R.; Hanus, L.O.; Pertwee, R.; Howlett, A.C. Early phytocannabinoid chemistry to endocannabinoids and beyond. Nat. Rev. Neurosci. 2014, 15, 757–764. [Google Scholar] [CrossRef]
- Lee, J.L.C.; Bertoglio, L.J.; Guimaraes, F.S.; Stevenson, C.W. Cannabidiol regulation of emotion and emotional memory processing: Relevance for treating anxiety-related and substance abuse disorders. Br. J. Pharmacol. 2017, 174, 3242–3256. [Google Scholar] [CrossRef] [Green Version]
- Ford, T.C.; Hayley, A.C.; Downey, L.A.; Parrott, A.C. Cannabis: An Overview of its Adverse Acute and Chronic Effects and its Implications. Curr. Drug Abuse Rev. 2017, 10, 6–18. [Google Scholar] [CrossRef] [Green Version]
- Carrier, E.J.; Auchampach, J.A.; Hillard, C.J. Inhibition of an equilibrative nucleoside transporter by cannabidiol: A mechanism of cannabinoid immunosuppression. Proc. Natl. Acad. Sci. USA 2006, 103, 7895–7900. [Google Scholar] [CrossRef] [Green Version]
- Pastor-Anglada, M.; Perez-Torras, S. Who Is Who in Adenosine Transport. Front. Pharmacol. 2018, 9, 627. [Google Scholar] [CrossRef] [Green Version]
- Liou, G.I.; Auchampach, J.A.; Hillard, C.J.; Zhu, G.; Yousufzai, B.; Mian, S.; Khan, S.; Khalifa, Y.M. Mediation of Cannabidiol Anti-inflammation in the Retina by Equilibrative Nucleoside Transporter and A2A Adenosine Receptor. Investig. Ophthalmol. Vis. Sci. 2008, 49, 5526–5531. [Google Scholar] [CrossRef] [Green Version]
- Castillo, A.; Tolon, M.R.; Fernandez-Ruiz, J.; Romero, J.; Martinez-Orgado, J. The neuroprotective effect of cannabidiol in an in vitro model of newborn hypoxic-ischemic brain damage in mice is mediated by CB(2) and adenosine receptors. Neurobiol. Dis. 2010, 37, 434–440. [Google Scholar] [CrossRef]
- Mecha, M.; Feliu, A.; Inigo, P.M.; Mestre, L.; Carrillo-Salinas, F.J.; Guaza, C. Cannabidiol provides long-lasting protection against the deleterious effects of inflammation in a viral model of multiple sclerosis: A role for A2A receptors. Neurobiol. Dis. 2013, 59, 141–150. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Wydra, K.; Fores-Pons, R.; Vasudevan, L.; Romero-Fernandez, W.; Frankowska, M.; Ferraro, L.; Beggiato, S.; Crespo-Ramirez, M.; Rivera, A.; et al. The Balance of MU-Opioid, Dopamine D2 and Adenosine A2A Heteroreceptor Complexes in the Ventral Striatal-Pallidal GABA Antireward Neurons May Have a Significant Role in Morphine and Cocaine Use Disorders. Front. Pharmacol. 2021, 12, 627032. [Google Scholar] [CrossRef]
- Ferre, S.; Lluis, C.; Justinova, Z.; Quiroz, C.; Orru, M.; Navarro, G.; Canela, E.I.; Franco, R.; Goldberg, S.R. Adenosine-cannabinoid receptor interactions. Implications for striatal function. Br. J. Pharmacol. 2010, 160, 443–453. [Google Scholar] [CrossRef] [Green Version]
- Boggs, D.L.; Nguyen, J.D.; Morgenson, D.; Taffe, M.A.; Ranganathan, M. Clinical and Preclinical Evidence for Functional Interactions of Cannabidiol and Delta(9)-Tetrahydrocannabinol. Neuropsychopharmacology 2018, 43, 142–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perucca, E.; Bialer, M. Critical Aspects Affecting Cannabidiol Oral Bioavailability and Metabolic Elimination, and Related Clinical Implications. CNS Drugs 2020, 34, 795–800. [Google Scholar] [CrossRef]
- Breuer, A.; Haj, C.G.; Fogaca, M.V.; Gomes, F.V.; Silva, N.R.; Pedrazzi, J.F.; Del Bel, E.A.; Hallak, J.C.; Crippa, J.A.; Zuardi, A.W.; et al. Fluorinated Cannabidiol Derivatives: Enhancement of Activity in Mice Models Predictive of Anxiolytic, Antidepressant and Antipsychotic Effects. PLoS ONE 2016, 11, e0158779. [Google Scholar]
- Silva, N.R.; Gomes, F.V.; Fonseca, M.D.; Mechoulam, R.; Breuer, A.; Cunha, T.M.; Guimaraes, F.S. Antinociceptive effects of HUF-101, a fluorinated cannabidiol derivative. Prog. Neuropsychopharmacol. Biol. Psychiatry 2017, 79, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.; Pertwee, R.G. A metabolic interaction in vivo between cannabidiol and 1 -tetrahydrocannabinol. Br. J. Pharmacol. 1972, 45, 375–377. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.L.; Gadegbeku, B.; Wu, D.; Viallon, V.; Laumon, B. Cannabis, alcohol and fatal road accidents. PLoS ONE 2017, 12, e0187320. [Google Scholar] [CrossRef]
- Monory, K.; Blaudzun, H.; Massa, F.; Kaiser, N.; Lemberger, T.; Schutz, G.; Wotjak, C.T.; Lutz, B.; Marsicano, G. Genetic dissection of behavioural and autonomic effects of Delta(9)-tetrahydrocannabinol in mice. PLoS Biol. 2007, 5, e269. [Google Scholar] [CrossRef]
- De Giacomo, V.; Ruehle, S.; Lutz, B.; Haring, M.; Remmers, F. Differential glutamatergic and GABAergic contributions to the tetrad effects of Delta(9)-tetrahydrocannabinol revealed by cell-type-specific reconstitution of the CB1 receptor. Neuropharmacology 2020, 179, 108287. [Google Scholar] [CrossRef]
- Soria-Gomez, E.; Pagano Zottola, A.C.; Mariani, Y.; Desprez, T.; Barresi, M.; Bonilla-Del Rio, I.; Muguruza, C.; Le Bon-Jego, M.; Julio-Kalajzic, F.; Flynn, R.; et al. Subcellular specificity of cannabinoid effects in striatonigral circuits. Neuron 2021, 109, 1513–1526.e11. [Google Scholar] [CrossRef]
- Carriba, P.; Ortiz, O.; Patkar, K.; Justinova, Z.; Stroik, J.; Themann, A.; Muller, C.; Woods, A.S.; Hope, B.T.; Ciruela, F.; et al. Striatal Adenosine A(2A) and Cannabinoid CB(1) Receptors Form Functional Heteromeric Complexes that Mediate the Motor Effects of Cannabinoids. Neuropsychopharmacology 2007, 32, 2249–2259. [Google Scholar] [CrossRef] [Green Version]
- Aso, E.; Fernandez-Duenas, V.; Lopez-Cano, M.; Taura, J.; Watanabe, M.; Ferrer, I.; Lujan, R.; Ciruela, F. Adenosine A2A-Cannabinoid CB1 Receptor Heteromers in the Hippocampus: Cannabidiol Blunts Delta(9)-Tetrahydrocannabinol-Induced Cognitive Impairment. Mol. Neurobiol. 2019, 56, 5382–5391. [Google Scholar] [CrossRef] [Green Version]
- DeMet, E.M.; Chicz-DeMet, A. Localization of adenosine A2A-receptors in rat brain with [3H]ZM-241385. Naunyn Schmiedebergs Arch. Pharmacol. 2002, 366, 478–481. [Google Scholar] [CrossRef] [PubMed]
- Rosin, D.L.; Hettinger, B.D.; Lee, A.; Linden, J. Anatomy of adenosine A2A receptors in brain: Morphological substrates for integration of striatal function. Neurology 2003, 61, S12–S18. [Google Scholar] [CrossRef] [PubMed]
- Gangarossa, G.; Espallergues, J.; Mailly, P.; De Bundel, D.; de Kerchove d’Exaerde, A.; Herve, D.; Girault, J.A.; Valjent, E.; Krieger, P. Spatial distribution of D1R- and D2R-expressing medium-sized spiny neurons differs along the rostro-caudal axis of the mouse dorsal striatum. Front. Neural Circuits 2013, 7, 124. [Google Scholar] [CrossRef] [Green Version]
- Tebano, M.T.; Martire, A.; Chiodi, V.; Pepponi, R.; Ferrante, A.; Domenici, M.R.; Frank, C.; Chen, J.F.; Ledent, C.; Popoli, P. Adenosine A2A receptors enable the synaptic effects of cannabinoid CB1 receptors in the rodent striatum. J. Neurochem. 2009, 110, 1921–1930. [Google Scholar] [CrossRef] [PubMed]
- Ferre, S.; Rubio, A.; Fuxe, K. Stimulation of adenosine A2 receptors induces catalepsy. Neurosci. Lett. 1991, 130, 162–164. [Google Scholar] [CrossRef]
- Mandhane, S.N.; Chopde, C.T.; Ghosh, A.K. Adenosine A2 receptors modulate haloperidol-induced catalepsy in rats. Eur. J. Pharmacol. 1997, 328, 135–141. [Google Scholar] [CrossRef]
- Long, L.E.; Chesworth, R.; Huang, X.F.; McGregor, I.S.; Arnold, J.C.; Karl, T. A behavioural comparison of acute and chronic Delta9-tetrahydrocannabinol and cannabidiol in C57BL/6JArc mice. Int. J. Neuropsychopharmacol. 2009, 13, 861–876. [Google Scholar] [CrossRef] [Green Version]
- Schiavon, A.P.; Bonato, J.M.; Milani, H.; Guimaraes, F.S.; Weffort de Oliveira, R.M. Influence of single and repeated cannabidiol administration on emotional behavior and markers of cell proliferation and neurogenesis in non-stressed mice. Prog. Neuropsychopharmacol. Biol. Psychiatry 2016, 64, 27–34. [Google Scholar] [CrossRef]
- Zieba, J.; Sinclair, D.; Sebree, T.; Bonn-Miller, M.; Gutterman, D.; Siegel, S.; Karl, T. Cannabidiol (CBD) reduces anxiety-related behavior in mice via an FMRP-independent mechanism. Pharmacol. Biochem. Behav. 2019, 181, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Scott, B.W.; Burnham, W.M. Effects of cannabidiol and Delta9-tetrahydrocannabinol in the elevated plus maze in mice. Behav. Pharmacol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Borgkvist, A.; Marcellino, D.; Fuxe, K.; Greengard, P.; Fisone, G. Regulation of DARPP-32 phosphorylation by Delta(9)-tetrahydrocannabinol. Neuropharmacology 2008, 54, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Marangos, P.J.; Boulenger, J.P. Basic and clinical aspects of adenosinergic neuromodulation. Neurosci. Biobehav. Rev. 1985, 9, 421–430. [Google Scholar] [CrossRef]
- Johansson, B.; Halldner, L.; Dunwiddie, T.V.; Masino, S.A.; Poelchen, W.; Gimenez-Llort, L.; Escorihuela, R.M.; Fernandez-Teruel, A.; Wiesenfeld-Hallin, Z.; Xu, X.J.; et al. Hyperalgesia, anxiety, and decreased hypoxic neuroprotection in mice lacking the adenosine A1 receptor. Proc. Natl. Acad. Sci. USA 2001, 98, 9407–9412. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Rinaldo, L.; Lim, S.J.; Young, H.; Messing, R.O.; Choi, D.S. The type 1 equilibrative nucleoside transporter regulates anxiety-like behavior in mice. Genes Brain Behav. 2007, 6, 776–783. [Google Scholar] [CrossRef]
- Perez, M.; Cartarozzi, L.P.; Chiarotto, G.B.; Oliveira, S.A.; Guimaraes, F.S.; Oliveira, A.L.R. Neuronal preservation and reactive gliosis attenuation following neonatal sciatic nerve axotomy by a fluorinated cannabidiol derivative. Neuropharmacology 2018, 140, 201–208. [Google Scholar] [CrossRef]
- Wydra, K.; Gawlinski, D.; Gawlinska, K.; Frankowska, M.; Borroto-Escuela, D.O.; Fuxe, K.; Filip, M. Adenosine A2AReceptors in Substance Use Disorders: A Focus on Cocaine. Cells 2020, 9, 1372. [Google Scholar] [CrossRef] [PubMed]
- Hurd, Y.L.; Spriggs, S.; Alishayev, J.; Winkel, G.; Gurgov, K.; Kudrich, C.; Oprescu, A.M.; Salsitz, E. Cannabidiol for the Reduction of Cue-Induced Craving and Anxiety in Drug-Abstinent Individuals with Heroin Use Disorder: A Double-Blind Randomized Placebo-Controlled Trial. Am. J. Psychiatry 2019, 176, 911–922. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.F.; Huang, Z.; Ma, J.; Zhu, J.; Moratalla, R.; Standaert, D.; Moskowitz, M.A.; Fink, J.S.; Schwarzschild, M.A. A(2A) adenosine receptor deficiency attenuates brain injury induced by transient focal ischemia in mice. J. Neurosci. 1999, 19, 9192–9200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cradock, J.C.; Davignon, J.P.; Litterst, C.L.; Guarino, A.M. An intravenous formulation of 9-tetrahydrocannabinol using a non-ionic surfactant. J. Pharm. Pharmacol. 1973, 25, 345. [Google Scholar] [CrossRef] [PubMed]
- Marshell, R.; Kearney-Ramos, T.; Brents, L.K.; Hyatt, W.S.; Tai, S.; Prather, P.L.; Fantegrossi, W.E. In vivo effects of synthetic cannabinoids JWH-018 and JWH-073 and phytocannabinoid Delta-THC in mice: Inhalation versus intraperitoneal injection. Pharmacol. Biochem. Behav. 2014, 124C, 40–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stollenwerk, T.M.; Pollock, S.; Hillard, C.J. Contribution of the Adenosine 2A Receptor to Behavioral Effects of Tetrahydrocannabinol, Cannabidiol and PECS-101. Molecules 2021, 26, 5354. https://doi.org/10.3390/molecules26175354
Stollenwerk TM, Pollock S, Hillard CJ. Contribution of the Adenosine 2A Receptor to Behavioral Effects of Tetrahydrocannabinol, Cannabidiol and PECS-101. Molecules. 2021; 26(17):5354. https://doi.org/10.3390/molecules26175354
Chicago/Turabian StyleStollenwerk, Todd M., Samantha Pollock, and Cecilia J. Hillard. 2021. "Contribution of the Adenosine 2A Receptor to Behavioral Effects of Tetrahydrocannabinol, Cannabidiol and PECS-101" Molecules 26, no. 17: 5354. https://doi.org/10.3390/molecules26175354
APA StyleStollenwerk, T. M., Pollock, S., & Hillard, C. J. (2021). Contribution of the Adenosine 2A Receptor to Behavioral Effects of Tetrahydrocannabinol, Cannabidiol and PECS-101. Molecules, 26(17), 5354. https://doi.org/10.3390/molecules26175354