
Could Polyphenols Really Be a Good Radioprotective Strategy?





Abstract
:1. Introduction
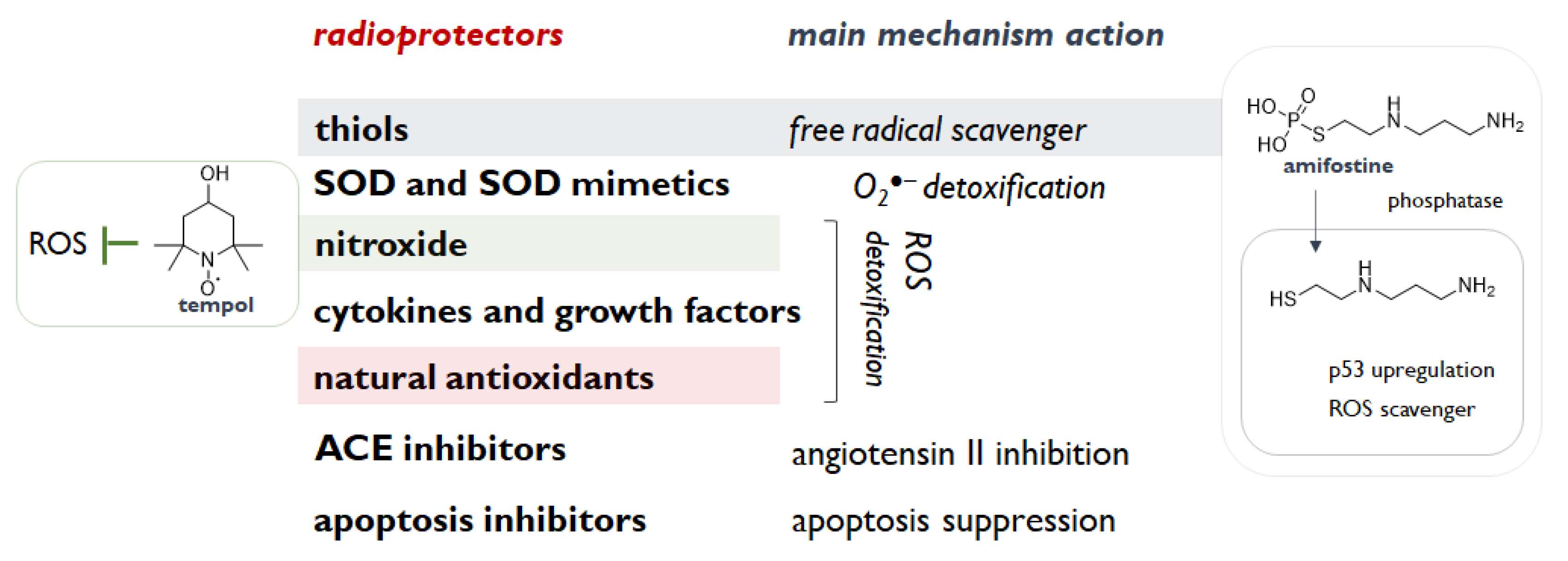
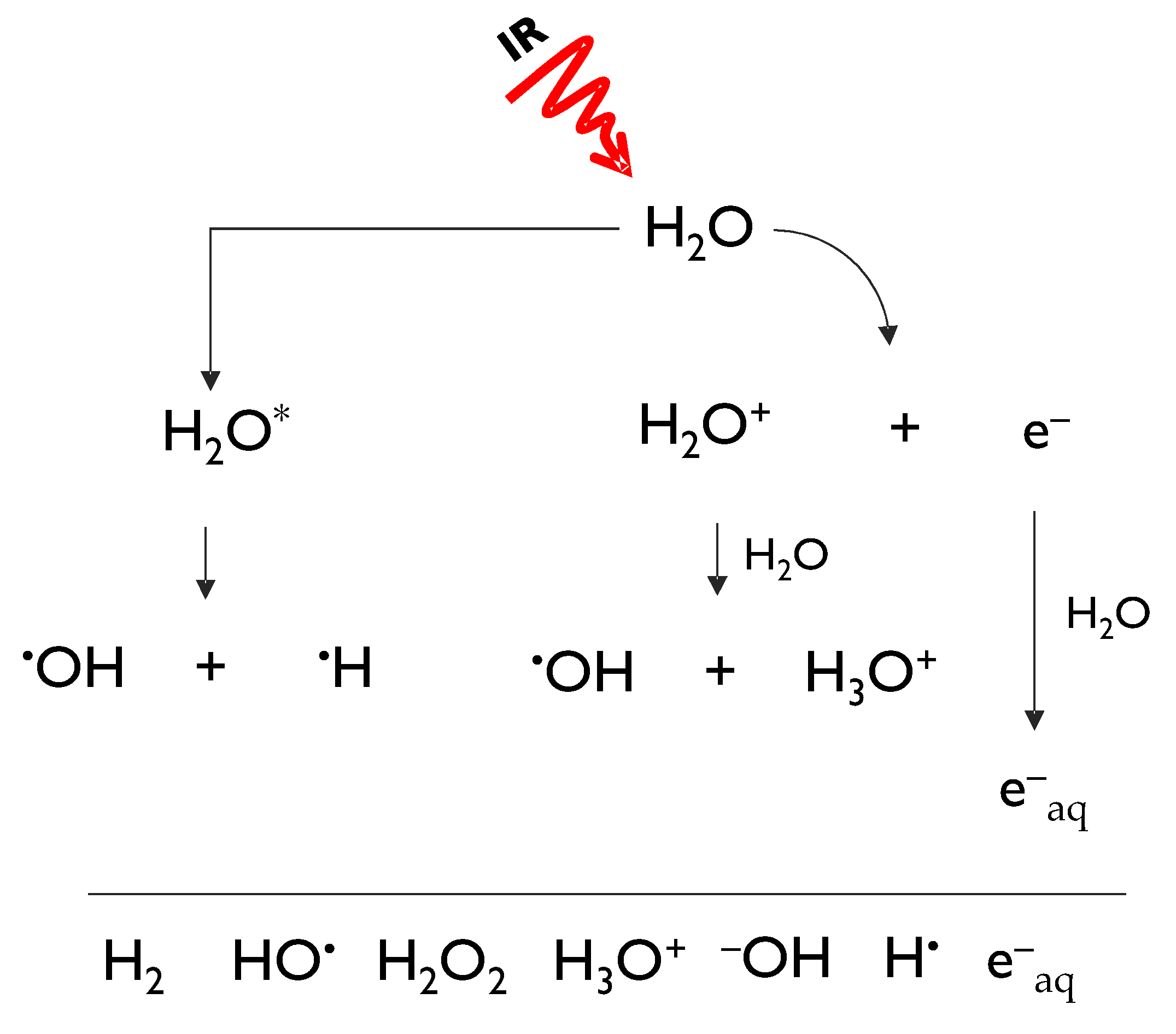
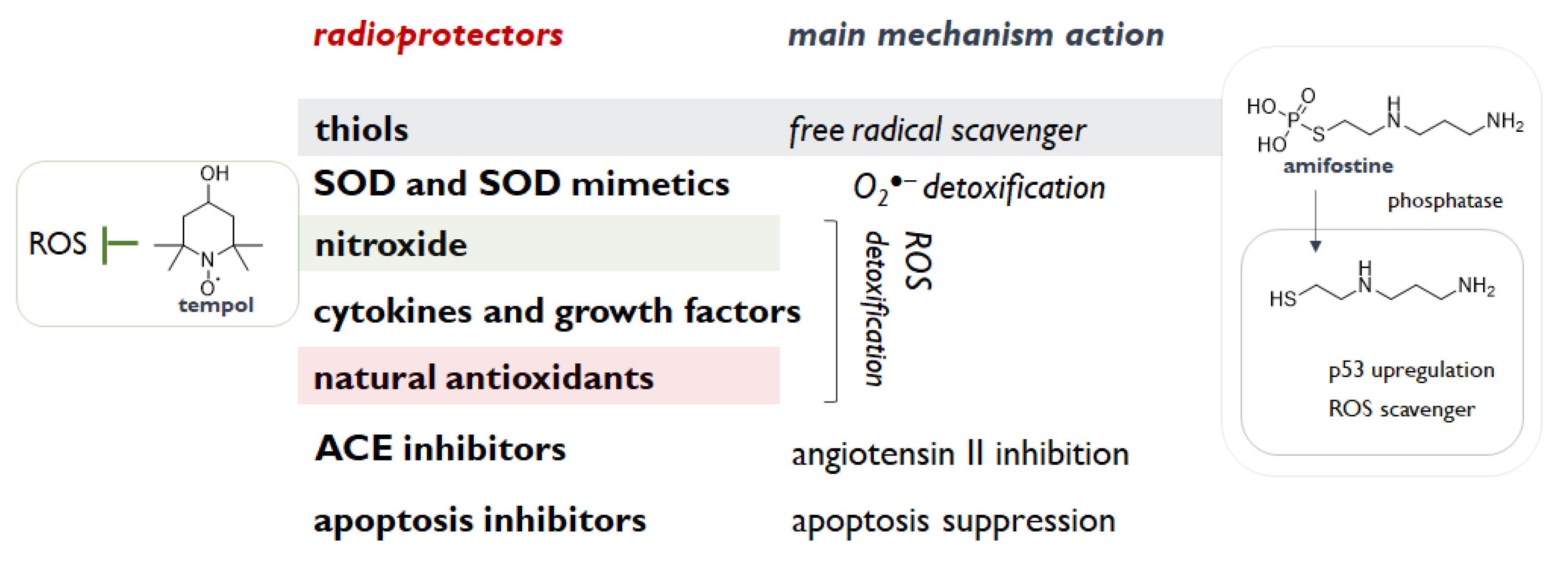
2. Radioprotection: A Valuable Approach to Counteract Radiation Exposure
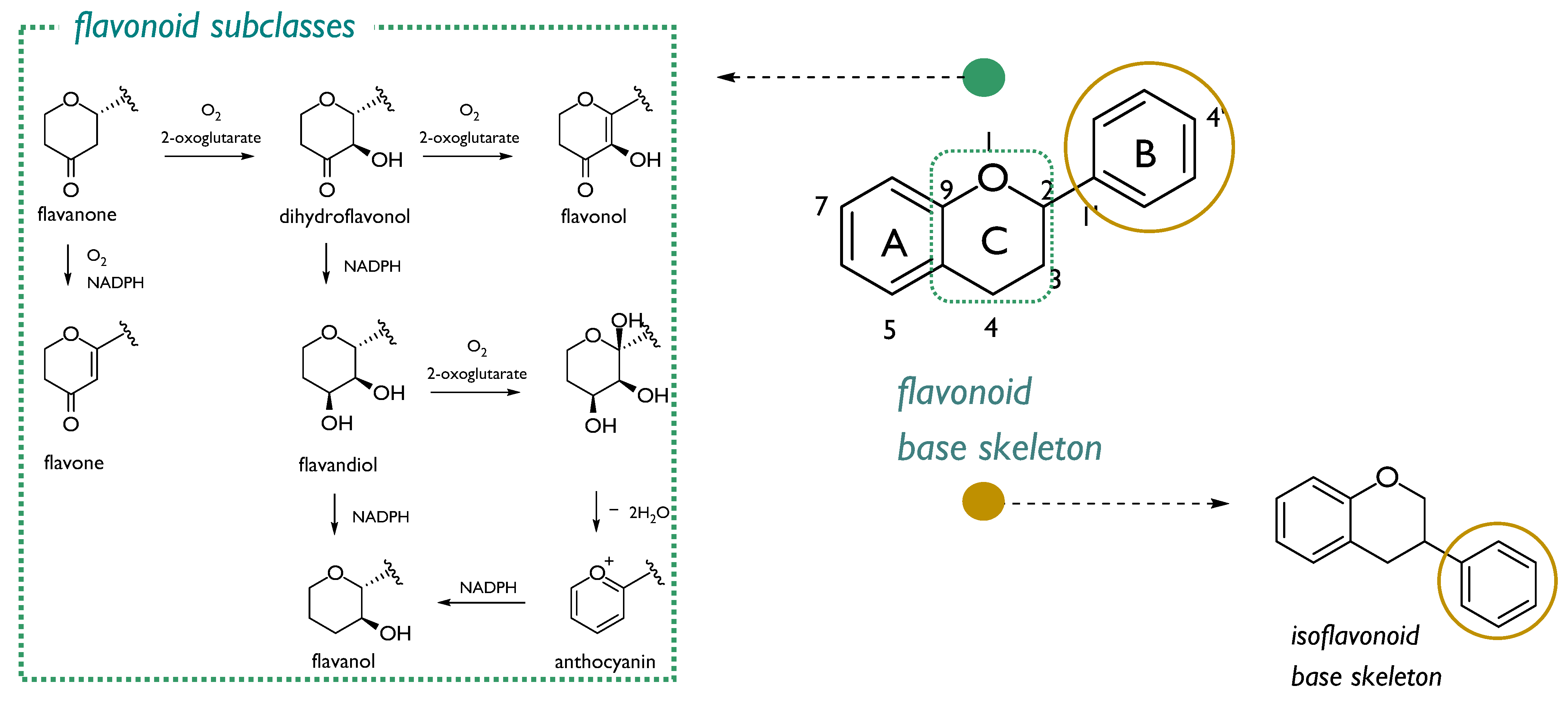
3. Phenols and Polyphenols: Are They a Valuable Radioprotective Strategy?
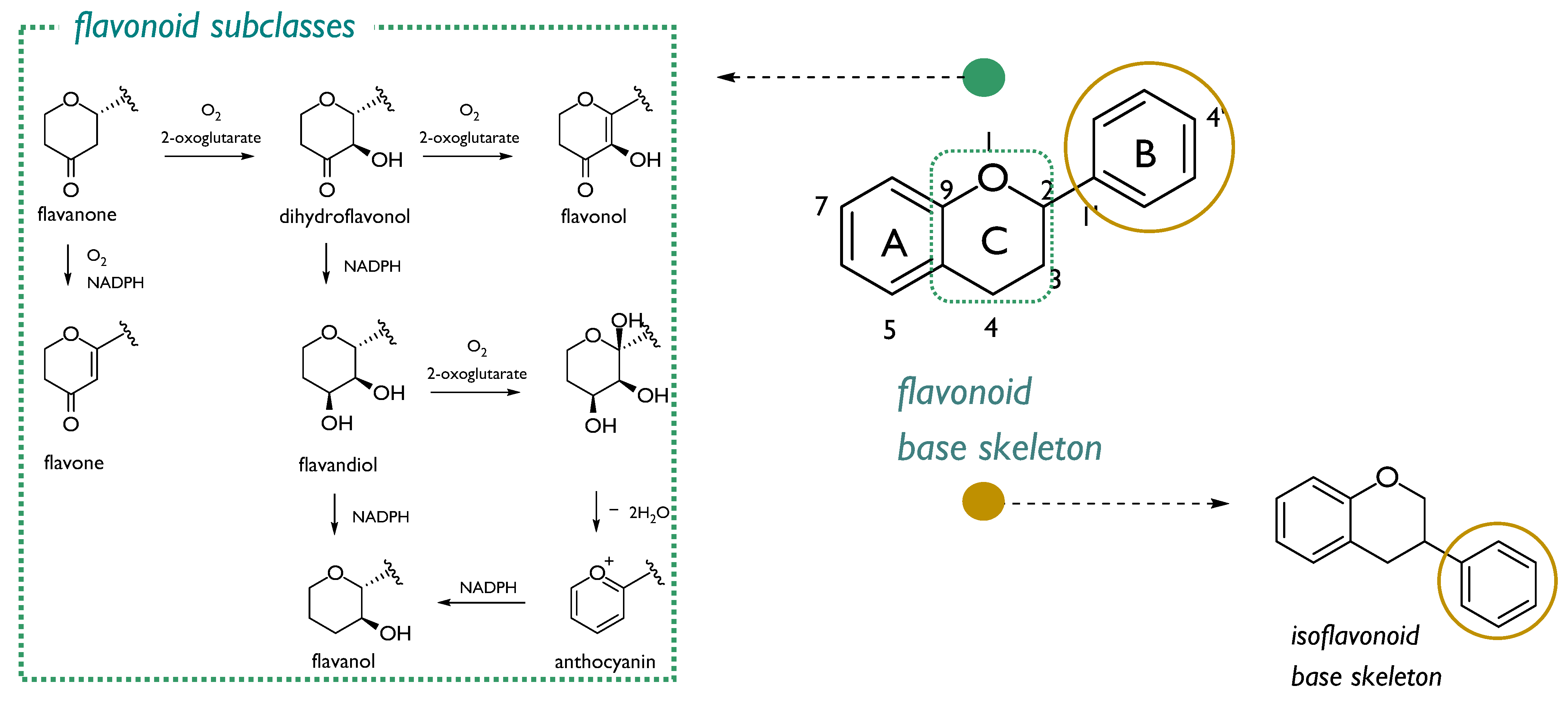
3.1. Flavonoids: The Double-Edged Sword in Radioprotection
3.2. Other Phenols and Polyphenols with Radioprotective Efficacy
4. Bioactive Plant Extracts in Radioprotection: A Still Undervalued Topic
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| ACE | Angiotensin Converting Enzyme |
| Akt | Protein kinase B |
| Bax | Bcl-2-associated X protein |
| Bcl-2 | B-cell lymphoma 2 |
| CAT | Catalase |
| CDK | Cyclin-Dependent Kinase |
| COX-2 | Cyclo-oxygenase-2 |
| DDR | DNA Damage Response |
| DPPH | 2,2-diphenyl-1-picryl hydrazyl |
| DSB | Double Strand Break |
| e−aq | Hydrated electrons |
| EGCG | Epigallocatechin-3-O-gallate |
| ERK | Extracellular signal-regulated kinase |
| ER-β | Estrogen Receptor-β |
| FOXL-2 | Forkhead box L2 protein |
| G-CSF | Granulocyte-Colony Stimulating Factor |
| GM-CSF | Granulocyte Macrophage-Colony Stimulating Factor |
| GPx | Glutathione peroxidase |
| GRP78 | 78-kDa Glucose-Regulated Protein |
| GSH | Reduced Glutathione |
| HO-1 | Heme Oxygenase-1 |
| HO2● | Hydroperoxyl radical |
| HSCs | Hematopoiesis Stem Cells |
| IL-1 | Interleukin-1 |
| IR | Ionizing Radiation |
| LPS | Lipopolysaccharide |
| MAPK | Mitogen-Activated Protein Kinase |
| MKP3 | MAP Kinase Phosphatase 3 |
| NF-κB | Nuclear factor kappa B |
| Nrf2-ARE | NF-E2-related factor 2-Antioxidant Responsive Element |
| O2−● | Superoxide anion radical |
| PSHE | Pterocarpus santalinus hydroalcoholic extract |
| PUMA | p53 Up-regulated Modulator of Apoptosis |
| RNF8 | Ring Finger Protein 8 |
| ROS | Reactive Oxygen Species |
| SIRT1 | Sirtuin 1 |
| SOD | Superoxide Dismutase |
| SSB | Single Strand Break |
| TGF-β | Transforming Growth Factor-β |
| TNF-α | Tumor Necrosis Factor-1 |
| UHPLC-HRMS/MS | Ultra High Performance Liquid Chromatography-High Resolution Tandem Mass Spectrometry |
References
- Belli, M.; Indovina, L. The Response of Living Organisms to Low Radiation Environment and Its Implications in Radiation Protection. Front. Public Health 2020, 8, 601711. [Google Scholar] [CrossRef]
- Goodhead, D.T. An Assessment of the Role of Microdosimetry in Radiobiology. Radiat. Res. 1982, 91, 45. [Google Scholar] [CrossRef]
- Jeggo, P.; Löbrich, M. Radiation-induced DNA damage responses. Radiat. Prot. Dosim. 2006, 122, 124–127. [Google Scholar] [CrossRef] [PubMed]
- Durante, M.; Manti, L. Human response to high-background radiation environments on Earth and in space. Adv. Space Res. 2008, 42, 999–1007. [Google Scholar] [CrossRef]
- Saha, G.B. Physics and Radiobiology of Nuclear Medicine; Springer Science & Business Media: Berlin, Heidelberg, Germany, 2012. [Google Scholar]
- Varanda, E.A.; Tavares, D.C. Radioprotection: Mechanisms and Radioprotective Agents Including Honeybee Venom. J. Venom. Anim. Toxins 1998, 4, 5–21. [Google Scholar] [CrossRef]
- Lomax, M.E.; Folkes, L.K.; O’Neill, P. Biological consequences of radiation-induced DNA damage: Relevance to radiotherapy. Clin. Oncol. 2013, 25, 578–585. [Google Scholar] [CrossRef] [Green Version]
- Manti, L.; Braselmann, H.; Calabrese, M.L.; Massa, R.; Pugliese, M.; Scampoli, P.; Sicignano, G.; Grossi, G. Effects of Modulated Microwave Radiation at Cellular Telephone Frequency (1.95 GHz) on X-ray-Induced Chromosome Aberrations in Human Lymphocytes In Vitro. Radiat. Res. 2008, 169, 575–583. [Google Scholar] [CrossRef]
- Wong, K.; Delaney, G.P.; Barton, M.B.; Information, P.E.K.F.C. Evidence-based optimal number of radiotherapy fractions for cancer: A useful tool to estimate radiotherapy demand. Radiother. Oncol. 2015, 119, 145–149. [Google Scholar] [CrossRef]
- Johnke, R.M.; Sattler, J.A.; Allison, R.R. Radioprotective agents for radiation therapy: Future trends. Futur. Oncol. 2014, 10, 2345–2357. [Google Scholar] [CrossRef]
- Andreassen, C.N.; Grau, C.; Lindegaard, J.C. Chemical radioprotection: A critical review of amifostine as a cytoprotector in radiotherapy. Semin. Radiat. Oncol. 2003, 13, 62–72. [Google Scholar] [CrossRef]
- Obrador, E.; Salvador, R.; Villaescusa, J.; Soriano, J.; Estrela, J.; Montoro, A. Radioprotection and Radiomitigation: From the Bench to Clinical Practice. Biomedicines 2020, 8, 461. [Google Scholar] [CrossRef]
- Mun, G.-I.; Kim, S.; Choi, E.; Kim, C.S.; Lee, Y.-S. Pharmacology of natural radioprotectors. Arch. Pharmacal Res. 2018, 41, 1033–1050. [Google Scholar] [CrossRef] [Green Version]
- Commission Regulation (EU) No 182/2013 of 1 March 2013 (EU commission N° 182 2013). Available online: https://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2013:061:0002:0005:EN:PDF (accessed on 5 March 2013).
- Patyar, R.R.; Patyar, S. Role of drugs in the prevention and amelioration of radiation induced toxic effects. Eur. J. Pharmacol. 2018, 819, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Sueiro-Benavides, R.A.; Leiro-Vidal, J.M.; Salas-Sánchez, A.A.; Rodríguez-González, J.A.; Ares-Pena, F.J.; López-Martín, M.E. Radiofrequency at 2.45 GHz increases toxicity, pro-inflammatory and pre-apoptotic activity caused by black carbon in the RAW 264.7 macrophage cell line. Sci. Total Environ. 2020, 765, 142681. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Snyder, A.R.; Morgan, W.F. Radiation-induced genomic instability and its implications for radiation carcinogenesis. Oncogene 2003, 22, 5848–5854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohan, S.; Gupta, D. Role of Nrf2-antioxidant in radioprotection by root extract of Inula racemosa. Int. J. Radiat. Biol. 2019, 95, 1122–1134. [Google Scholar] [CrossRef]
- Nair, C.K.K.; Parida, D.K.; Nomura, T. Radioprotectors in Radiotherapy. J. Radiat. Res. 2001, 42, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.B.; Anver, M.R.; Sowers, A.L.; Rosenberg, P.; Figueroa, M.; Thetford, A.; Krishna, M.C.; Albert, P.S.; Cook, J.A. The Antioxidant Tempol Reduces Carcinogenesis and Enhances Survival in Mice When Administered after Nonlethal Total Body Radiation. Cancer Res. 2012, 72, 4846–4855. [Google Scholar] [CrossRef] [Green Version]
- Rosen, E.M.; Day, R.; Singh, V.K. New Approaches to Radiation Protection. Front. Oncol. 2015, 4, 381. [Google Scholar] [CrossRef] [Green Version]
- Li, M. The role of P53 up-regulated modulator of apoptosis (PUMA) in ovarian development, cardiovascular and neurodegenerative diseases. Apoptosis 2021, 26, 235–247. [Google Scholar] [CrossRef]
- Brown, S.L.; Kolozsvary, A.; Liu, J.; Jenrow, K.A.; Ryu, S.; Kim, J.H. Antioxidant Diet Supplementation Starting 24 Hours after Exposure Reduces Radiation Lethality. Radiat. Res. 2010, 173, 462–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farhood, B.; Goradel, N.H.; Mortezaee, K.; Khanlarkhani, N.; Najafi, M.; Sahebkar, A. Melatonin and cancer: From the promotion of genomic stability to use in cancer treatment. J. Cell. Physiol. 2019, 234, 5613–5627. [Google Scholar] [CrossRef] [PubMed]
- Hazra, B.; Ghosh, S.; Kumar, A.; Pandey, B.N. The Prospective Role of Plant Products in Radiotherapy of Cancer: A Current Overview. Front. Pharmacol. 2012, 2, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvaruso, M.; Pucci, G.; Musso, R.; Bravatà, V.; Cammarata, F.P.; Russo, G.; Forte, G.I.; Minafra, L. Nutraceutical Compounds as Sensitizers for Cancer Treatment in Radiation Therapy. Int. J. Mol. Sci. 2019, 20, 5267. [Google Scholar] [CrossRef] [Green Version]
- Piccolella, S.; Pacifico, S. Plant-Derived Polyphenols. Adv. Mol. Toxicol. 2015, 9, 161–214. [Google Scholar] [CrossRef]
- Kemertelidze, E.P.; Tsitsishvili, V.G.; Alaniya, M.D.; Sagareishvili, T.G. Structure-function analysis of the radioprotective and antioxidant activity of flavonoids. Chem. Nat. Compd. 2000, 36, 54–59. [Google Scholar] [CrossRef]
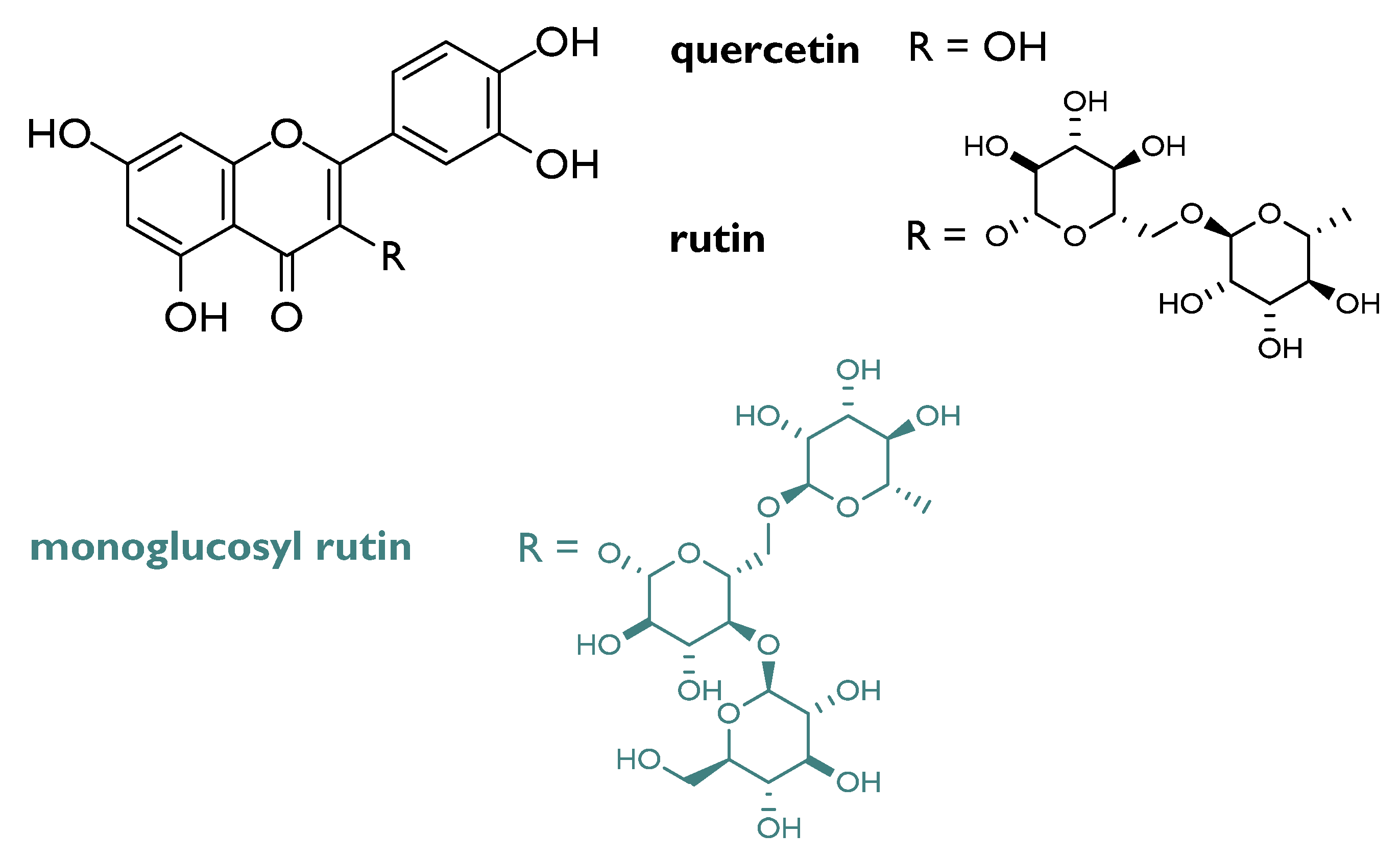
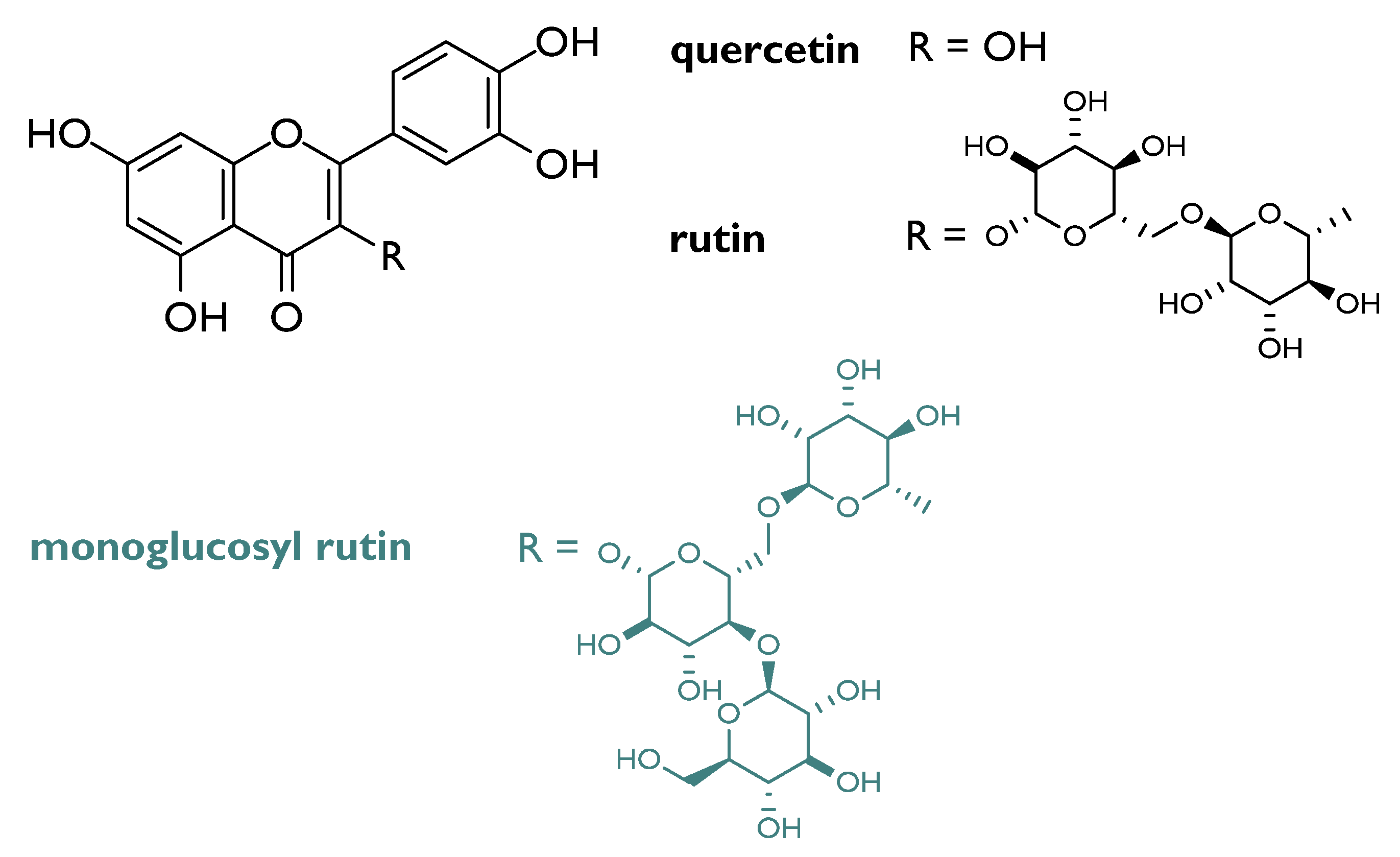
- Yashavarddhan, M.H.; Shukla, S.K.; Chaudhary, P.; Srivastava, N.N.; Joshi, J.; Suar, M.; Gupta, M.L. Targeting DNA Repair through Podophyllotoxin and Rutin Formulation in Hematopoietic Radioprotection: An In Silico, In Vitro, and In Vivo Study. Front. Pharmacol. 2017, 8, 750. [Google Scholar] [CrossRef] [Green Version]
- Dutta, A.; Gupta, M.L.; Kalita, B. The combination of the active principles of Podophyllum hexandrumsupports early recovery of the gastrointestinal system via activation of Nrf2-HO-1 signaling and the hematopoietic system, leading to effective whole-body survival in lethally irradiated mice. Free. Radic. Res. 2015, 49, 317–330. [Google Scholar] [CrossRef] [PubMed]
- Dutta, A.; Verma, S.; Sankhwar, S.; Flora, S.; Gupta, M.L. Bioavailability, antioxidant and non toxic properties of a radioprotective formulation prepared from isolated compounds of Podophyllum hexandrum: A study in mouse model. Cell. Mol. Boil. 2012, 58. [Google Scholar]
- Sunada, S.; Fujisawa, H.; Cartwright, I.M.; Maeda, J.; Brents, C.A.; Mizuno, K.; Aizawa, Y.; Kato, T.A.; Uesaka, M. Monoglucosyl-rutin as a potential radioprotector in mammalian cells. Mol. Med. Rep. 2014, 10, 10–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.; Haskins, J.S.; Su, C.; Allum, A.; Haskins, A.H.; Salinas, V.A.; Sunada, S.; Inoue, T.; Aizawa, Y.; Uesaka, M.; et al. In vitro screening of radioprotective properties in the novel glucosylated flavonoids. Int. J. Mol. Med. 2016, 38, 1525–1530. [Google Scholar] [CrossRef]
- Benković, V.; Kopjar, N.; Kneževic, A.H.; Đikić, D.; Bašić, I.; Ramić, S.; Viculin, T.; Knežević, F.; Orolić, N. Evaluation of Radioprotective Effects of Propolis and Quercetin on Human White Blood Cells in Vitro. Biol. Pharm. Bull. 2008, 31, 1778–1785. [Google Scholar] [CrossRef] [Green Version]
- Benkovic, V.; Knezevic, A.H.; Dikic, D.; Lisicic, D.; Orsolic, N.; Basic, I.; Kosalec, I.; Kopjar, N. Radioprotective effects of propolis and quercetin in γ-irradiated mice evaluated by the alkaline comet assay. Phytomedicine 2008, 15, 851–858. [Google Scholar] [CrossRef]
- Khayyal, M.T.; Abdel-Naby, D.H.; El-Ghazaly, M.A. Propolis extract protects against radiation-induced intestinal mucositis through anti-apoptotic mechanisms. Environ. Sci. Pollut. Res. 2019, 26, 24672–24682. [Google Scholar] [CrossRef]
- Karapetsas, A.; Voulgaridou, G.-P.; Konialis, M.; Tsochantaridis, I.; Kynigopoulos, S.; Lambropoulou, M.; Stavropoulou, M.-I.; Stathopoulou, K.; Aligiannis, N.; Bozidis, P.; et al. Propolis Extracts Inhibit UV-Induced Photodamage in Human Experimental In Vitro Skin Models. Antioxidants 2019, 8, 125. [Google Scholar] [CrossRef] [Green Version]
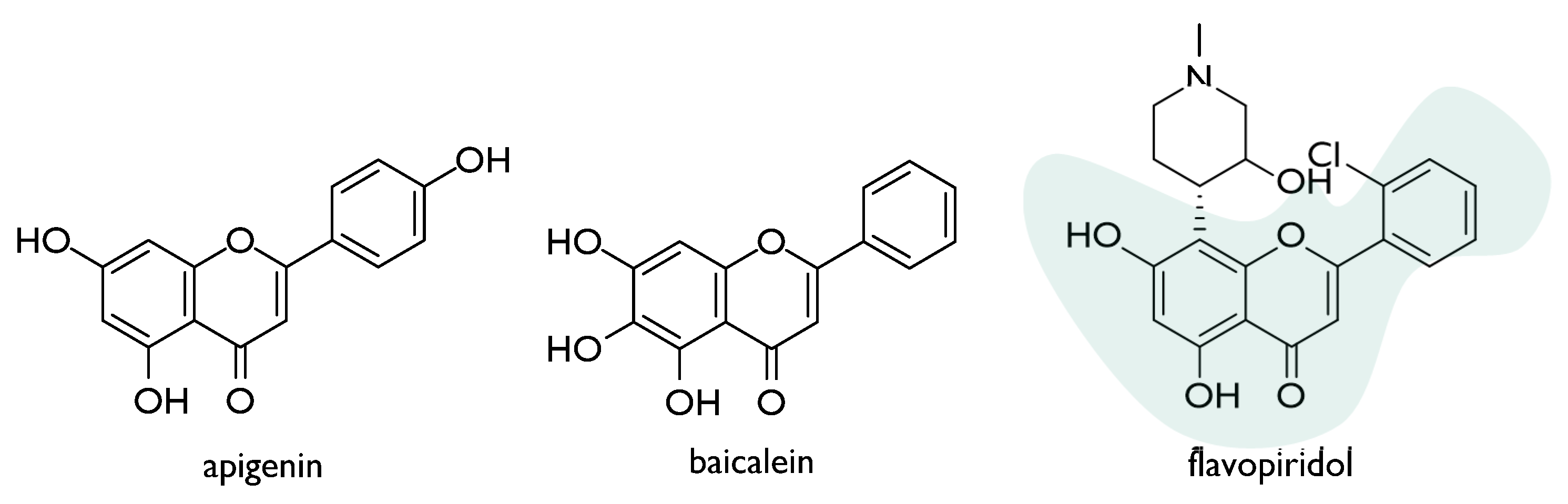
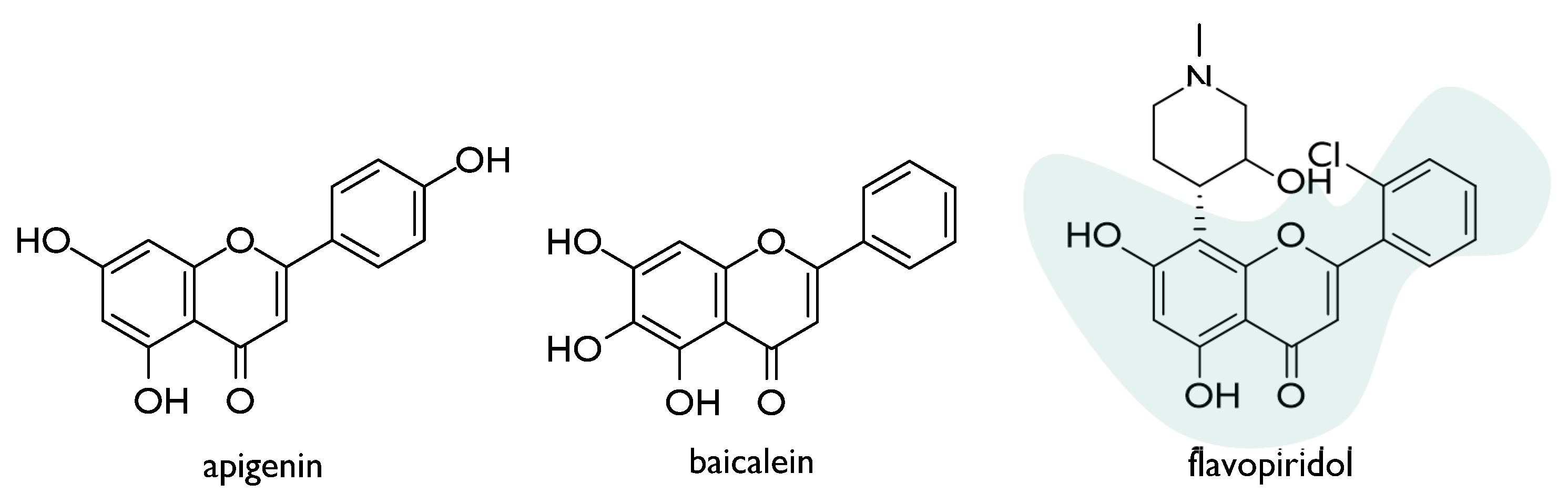
- Rithidech, K.N.; Tungjai, M.; Whorton, E.B. Protective effect of apigenin on radiation-induced chromosomal damage in human lymphocytes. Mutat. Res. Toxicol. Environ. Mutagen. 2005, 585, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Prasad, R.; Thayalan, K.; Begum, N. Apigenin protects gamma-radiation induced oxidative stress, hematological changes and animal survival in whole body irradiated Swiss albino mice. Int. J. Nutr. Pharmacol. Neurol. Dis. 2012, 2, 45–52. [Google Scholar] [CrossRef]
- Begum, N.; Prasad, N.R.; Kanimozhi, G.; Agilan, B. Apigenin prevents gamma radiation-induced gastrointestinal damages by modulating inflammatory and apoptotic signalling mediators. Nat. Prod. Res. 2021, 1–5. [Google Scholar] [CrossRef]
- Manna, K.; Khan, A.; Biswas, S.; Das, U.; Sengupta, A.; Mukherjee, D.; Chakraborty, A.; Dey, S. Naringin ameliorates radiation-induced hepatic damage through modulation of Nrf2 and NF-κB pathways. RSC Adv. 2016, 6, 23058–23073. [Google Scholar] [CrossRef]
- Saha, S.; Buttari, B.; Panieri, E.; Profumo, E.; Saso, L. An Overview of Nrf2 Signaling Pathway and Its Role in Inflammation. Molecules 2020, 25, 5474. [Google Scholar] [CrossRef] [PubMed]
- Patwardhan, R.; Sharma, D.; Checker, R.; Sandur, S.K. Mitigation of radiation-induced hematopoietic injury via regulation of cellular MAPK/phosphatase levels and increasing hematopoietic stem cells. Free. Radic. Biol. Med. 2014, 68, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sano, S.; Yura, Y.; Ke, Z.; Sano, M.; Oshima, K.; Ogawa, H.; Horitani, K.; Min, K.-D.; Miura-Yura, E.; et al. Tet2-mediated clonal hematopoiesis in nonconditioned mice accelerates age-associated cardiac dysfunction. JCI Insight 2020, 5, e135204. [Google Scholar] [CrossRef] [Green Version]
- Gandhi, N.M. Baicalein protects mice against radiation-induced DNA damages and genotoxicity. Mol. Cell. Biochem. 2013, 379, 277–281. [Google Scholar] [CrossRef]
- Lee, E.K.; Kim, J.M.; Choi, J.; Jung, K.J.; Kim, D.H.; Chung, S.W.; Ha, Y.M.; Yu, B.P.; Chung, H.Y. Modulation of NF-κB and FOXOs by baicalein attenuates the radiation-induced inflammatory process in mouse kidney. Free. Radic. Res. 2011, 45, 507–517. [Google Scholar] [CrossRef]
- Davari, H.; Haddad, F.; Moghimi, A.; Rahimi, M.F.; Ghavamnasiri, M.R. Study of Radioprotective Effect of Green Tea against Gamma Irradiation Using Micronucleus Assay on Binucleated Human Lymphocytes. Iran. J. Basic Med. Sci. 2012, 15, 1026–1031. [Google Scholar]
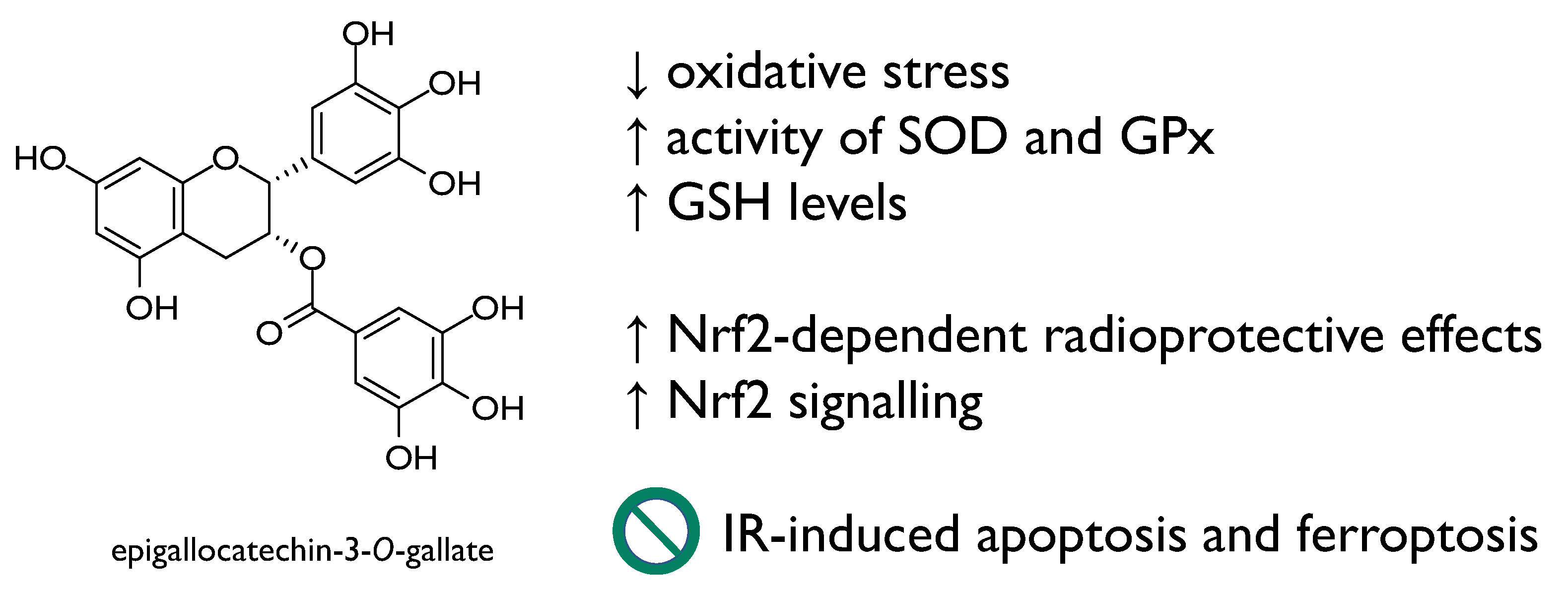
- Morley, N.; Clifford, T.; Salter, L.; Campbell, S.; Gould, D.; Curnow, A. The green tea polyphenol (-)-epigallocatechin gallate and green tea can protect human cellular DNA from ultraviolet and visible radiation-induced damage. Photodermatol. Photoimmunol. Photomed. 2005, 21, 15–22. [Google Scholar] [CrossRef]
- Xie, L.-W.; Cai, S.; Zhao, T.-S.; Li, M.; Tian, Y. Green Tea Derivative (−)-Epigallocatechin-3-Gallate (EGCG) Confers Protec-tion against Ionizing Radiation-Induced Intestinal Epithelial Cell Death Both In Vitro and In Vivo. Free Radic. Biol. Med. 2020, 161, 175–186. [Google Scholar] [CrossRef]
- Yi, J.; Chen, C.; Liu, X.; Kang, Q.; Hao, L.; Huang, J.; Lu, J. Radioprotection of EGCG based on immunoregulatory effect and antioxidant activity against 60Coγ radiation-induced injury in mice. Food Chem. Toxicol. 2020, 135, 111051. [Google Scholar] [CrossRef] [PubMed]
- Fountain, M.D.; McLellan, L.A.; Smith, N.L.; Loughery, B.F.; Rakowski, J.T.; Tse, H.Y.; Hillman, G.G. Isoflavone-mediated radioprotection involves regulation of early endothelial cell death and inflammatory signaling in Radiation-Induced lung injury. Int. J. Radiat. Biol. 2019, 96, 245–256. [Google Scholar] [CrossRef]
- Song, L.; Ma, L.; Cong, F.; Shen, X.; Jing, P.; Ying, X.; Zhou, H.; Jiang, J.; Fu, Y.; Yan, H. Radioprotective effects of genistein on HL-7702 cells via the inhibition of apoptosis and DNA damage. Cancer Lett. 2015, 366, 100–111. [Google Scholar] [CrossRef]
- Yan, H.; Jiang, J.; Du, A.; Gao, J.; Zhang, D.; Song, L. Genistein Enhances Radiosensitivity of Human Hepatocellular Carcinoma Cells by Inducing G2/M Arrest and Apoptosis. Radiat. Res. 2020, 193, 286. [Google Scholar] [CrossRef] [PubMed]
- Landauer, M.R.; Srinivasan, V.; Seed, T.M. Genistein treatment protects mice from ionizing radiation injury. J. Appl. Toxicol. 2003, 23, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Landauer, M.R.; Harvey, A.J.; Kaytor, M.D.; Day, R.M. Mechanism and therapeutic window of a genistein nanosuspension to protect against hematopoietic-acute radiation syndrome. J. Radiat. Res. 2019, 60, 308–317. [Google Scholar] [CrossRef]
- Haddad, Y.H.; Said, R.S.; Kamel, R.; Morsy, E.M.E.; El-Demerdash, E. Phytoestrogen genistein hinders ovarian oxidative damage and apoptotic cell death-induced by ionizing radiation: Co-operative role of ER-β, TGF-β, and FOXL. Sci. Rep. 2020, 10, 13551. [Google Scholar] [CrossRef] [PubMed]
- Hillman, G.G. Soy Isoflavones Protect Normal Tissues While Enhancing Radiation Responses. Semin. Radiat. Oncol. 2018, 29, 62–71. [Google Scholar] [CrossRef]
- Hara, T.; Omura-Minamisawa, M.; Kang, Y.; Cheng, C.; Inoue, T. Flavopiridol Potentiates the Cytotoxic Effects of Radiation in Radioresistant Tumor Cells in Which p53 is Mutated or Bcl-2 is Overexpressed. Int. J. Radiat. Oncol. 2008, 71, 1485–1495. [Google Scholar] [CrossRef]
- Hu, Y.; Ma, Y.; Wu, S.; Chen, T.; He, Y.; Sun, J.; Jiao, R.; Jiang, X.; Huang, Y.; Deng, L.; et al. Protective Effect of Cyanidin-3-O-Glucoside against Ultraviolet B Radiation-Induced Cell Damage in Human HaCaT Keratinocytes. Front. Pharmacol. 2016, 7, 301. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Hu, Y.; Jiang, X.; Chen, T.; Ma, Y.; Wu, S.; Sun, J.; Jiao, R.; Li, X.; Deng, L.; et al. Cyanidin-3-O-glucoside inhibits the UVB-induced ROS/COX-2 pathway in HaCaT cells. J. Photochem. Photobiol. B Biol. 2017, 177, 24–31. [Google Scholar] [CrossRef]
- Liu, Z.; Hu, Y.; Li, X.; Mei, Z.; Wu, S.; He, Y.; Jiang, X.; Sun, J.; Xiao, J.; Deng, L.; et al. Nanoencapsulation of Cyanidin-3-O-glucoside Enhances Protection Against UVB-Induced Epidermal Damage through Regulation of p53-Mediated Apoptosis in Mice. J. Agric. Food Chem. 2018, 66, 5359–5367. [Google Scholar] [CrossRef]
- Maurya, D.K.; Devasagayam, T.P.A. Indian medicinal herbs and ayurvedic formulations as potential radioprotectors. In Herbal Radiomodulators: Applications in Medicine, Homeland Defence and Space; CABI: Wallingford, UK, 2008; pp. 25–46. [Google Scholar]
- Li, M.; Gu, M.-M.; Lang, Y.; Shi, J.; Chen, B.P.; Guan, H.; Yu, L.; Zhou, P.-K.; Shang, Z.-F. The vanillin derivative VND3207 protects intestine against radiation injury by modulating p53/NOXA signaling pathway and restoring the balance of gut microbiota. Free. Radic. Biol. Med. 2019, 145, 223–236. [Google Scholar] [CrossRef]
- Li, M.; Lang, Y.; Gu, M.-M.; Shi, J.; Chen, B.P.; Yu, L.; Zhou, P.-K.; Shang, Z.-F. Vanillin derivative VND3207 activates DNA-PKcs conferring protection against radiation-induced intestinal epithelial cells injury in vitro and in vivo. Toxicol. Appl. Pharmacol. 2019, 387, 114855. [Google Scholar] [CrossRef]
- Sim, H.-J.; Bhattarai, G.; Lee, J.; Lee, J.-C.; Kook, S.-H. The Long-lasting Radioprotective Effect of Caffeic Acid in Mice Exposed to Total Body Irradiation by Modulating Reactive Oxygen Species Generation and Hematopoietic Stem Cell Senescence-Accompanied Long-term Residual Bone Marrow Injury. Aging Dis. 2019, 10, 1320–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurya, D.K.; Salvi, V.P.; Nair, C.K.K. Radiation protection of DNA by ferulic acid under in vitro and in vivo conditions. Mol. Cell. Biochem. 2005, 280, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Cinkilic, N.; Cetintas, S.K.; Zorlu, T.; Vatan, O.; Yilmaz, D.; Cavas, T.; Tunc, S.; Ozkan, L.; Bilaloglu, R. Radioprotection by two phenolic compounds: Chlorogenic and quinic acid, on X-ray induced DNA damage in human blood lymphocytes in vitro. Food Chem. Toxicol. 2013, 53, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Hosseinimehr, S.J.; Zakaryaee, V.; Ahmadi, A.; Akhlaghpoor, S. Radioprotective effects of chlorogenic acid against mortality induced by gamma irradiation in mice. Methods Find. Exp. Clin. Pharmacol. 2008, 30, 13–16. [Google Scholar] [CrossRef] [PubMed]
- Taysi, S.; Demir, E.; Cinar, K.; Tarakcioglu, M. The Radioprotective Effects of Propolis and Caffeic Acid Phenethyl Ester on Radiation-Induced Oxidative/nitrosative Stress in Brain Tissue. Free. Radic. Biol. Med. 2016, 100, S111. [Google Scholar] [CrossRef]
- Anjaly, K. Radio-Modulatory Potential of Caffeic Acid Phenethyl Ester: A Therapeutic Perspective. Anti-Cancer Agents Med. Chem. 2018, 18, 468–475. [Google Scholar] [CrossRef]
- Xu, W.; Yang, F.; Zhang, Y.; Shen, X. Protective effects of rosmarinic acid against radiation-induced damage to the hematopoietic system in mice. J. Radiat. Res. 2016, 57, 356–362. [Google Scholar] [CrossRef] [Green Version]
- Del Baño, M.J.; Castillo, J.; Benavente-García, O.; Lorente, J.; Martín-Gil, R.; Acevedo, C.; Alcaraz, M. Radioprotective−Antimutagenic Effects of Rosemary Phenolics against Chromosomal Damage Induced in Human Lymphocytes by γ-rays. J. Agric. Food Chem. 2006, 54, 2064–2068. [Google Scholar] [CrossRef]
- Kalman, D.S.; Hewlings, S.J. The Effects of Morus alba and Acacia catechu on Quality of Life and Overall Function in Adults with Osteoarthritis of the Knee. J. Nutr. Metab. 2017, 2017, 4893104. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.C.; Kinniry, P.A.; Arguiri, E.; Serota, M.; Kanterakis, S.; Chatterjee, S.; Solomides, C.C.; Javvadi, P.; Koumenis, C.; Cengel, K.A.; et al. Dietary Curcumin Increases Antioxidant Defenses in Lung, Ameliorates Radiation-Induced Pulmonary Fibrosis, and Improves Survival in Mice. Radiat. Res. 2010, 173, 590–601. [Google Scholar] [CrossRef] [Green Version]
- Akpolat, M.; Kanter, M.; Uzal, M.C. Protective effects of curcumin against gamma radiation-induced ileal mucosal damage. Arch. Toxicol. 2008, 83, 609–617. [Google Scholar] [CrossRef] [Green Version]
- Seong, K.M.; Yu, M.; Lee, K.-S.; Park, S.; Jin, Y.W.; Min, K.-J. Curcumin Mitigates Accelerated Aging after Irradiation in Drosophila by Reducing Oxidative Stress. BioMed Res. Int. 2015, 2015, 425380. [Google Scholar] [CrossRef] [PubMed]
- Verma, V. Relationship and interactions of curcumin with radiation therapy. World J. Clin. Oncol. 2016, 7, 275–283. [Google Scholar] [CrossRef]
- Shabeeb, D.; Musa, A.E.; Ali, H.S.A.; Najafi, M. Curcumin Protects Against Radiotherapy-Induced Oxidative Injury to the Skin. Drug Des. Dev. Ther. 2020, 14, 3159–3163. [Google Scholar] [CrossRef]
- Anand, P.; Kunnumakkara, A.B.; Newman, R.A.; Aggarwal, B.B. Bioavailability of Curcumin: Problems and Promises. Mol. Pharm. 2007, 4, 807–818. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.-H.; Pham, N.-D.; Dong, B.; Nguyen, T.-H.-N.; Bui, C.B.; Hadinoto, K. Radioprotective activity of curcumin-encapsulated liposomes against genotoxicity caused by Gamma Cobalt-60 irradiation in human blood cells. Int. J. Radiat. Biol. 2017, 93, 1267–1273. [Google Scholar] [CrossRef] [PubMed]
- Nosrati, H.; Danafar, H.; Rezaeejam, H.; Gholipour, N.; Rahimi-Nasrabadi, M. Evaluation radioprotective effect of curcumin conjugated albumin nanoparticles. Bioorganic Chem. 2020, 100, 103891. [Google Scholar] [CrossRef]
- Sasidharan, S.; Chen, Y.; Saravanan, D.; Sundram, K.M.; Latha, L.Y. Extraction, isolation and characterization of bioactive compounds from plants’ extracts. AJTCAM 2011, 8, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jagetia, G.C.; Shirwaikar, A.; Rao, S.K.; Bhilegaonkar, P.M. Evaluation of the radioprotective effect of Ageratum conyzoides Linn. extract in mice exposed to different doses of gamma radiation. J. Pharm. Pharmacol. 2010, 55, 1151–1158. [Google Scholar] [CrossRef] [PubMed]
- Samarth, R.M.; Samarth, M. Protection against Radiation-induced Testicular Damage in Swiss Albino Mice byMentha piperita(Linn.). Basic Clin. Pharmacol. Toxicol. 2009, 104, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Weiss, E.I.; Shaniztki, B.; Dotan, M.; Ganeshkumar, N.; Kolenbrander, P.E.; Metzger, Z. Attachment of Fusobacterium nucleatum PK1594 to mammalian cells and its coaggregation with periodontopathogenic bacteria are mediated by the same galactose-binding adhesin. Oral Microbiol. Immunol. 2000, 15, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Bump, E.A.; Brown, J. Role of glutathione in the radiation response of mammalian cells invitro and in vivo. Pharmacol. Ther. 1990, 47, 117–136. [Google Scholar] [CrossRef]
- Murray, A.P.; Rodriguez, S.; Frontera, M.A.; Tomas, M.A.; Mulet, M.C. Antioxidant Metabolites from Limonium brasiliense (Boiss.) Kuntze. Z. Naturforsch. C 2004, 59, 477–480. [Google Scholar] [CrossRef] [PubMed]
- Piccolella, S.; Crescente, G.; Nocera, P.; Pacifico, F.; Manti, L.; Pacifico, S. Ultrasound-assisted aqueous extraction, LC-MS/MS analysis and radiomodulating capability of autochthonous Italian sweet cherry fruits. Food Funct. 2018, 9, 1840–1849. [Google Scholar] [CrossRef] [PubMed]
- Castillo, J.J.; Alcaraz, M.; Benavente-García, O. Antioxidant and Radioprotective Effects of Olive Leaf Extract. In Olives and Olive Oil in Health and Disease Prevention; Elsevier BV: Amsterdam, The Netherlands, 2010; pp. 951–958. [Google Scholar]
- Xu, T.; Xiao, D. Oleuropein enhances radiation sensitivity of nasopharyngeal carcinoma by downregulating PDRG1 through HIF1α-repressed microRNA-519d. J. Exp. Clin. Cancer Res. 2017, 36, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, H.-Y.; Yang, J.-J.; Lin, C.-C. Effects of oleanolic acid and ursolic acid on inhibiting tumor growth and enhancing the recovery of hematopoietic system postirradiation in mice. Cancer Lett. 1997, 111, 7–13. [Google Scholar] [CrossRef]
- Wang, H.; Sim, M.-K.; Loke, W.K.; Chinnathambi, A.; Alharbi, S.A.; Tang, F.R.; Sethi, G. Potential Protective Effects of Ursolic Acid against Gamma Irradiation-Induced Damage Are Mediated through the Modulation of Diverse Inflammatory Mediators. Front. Pharmacol. 2017, 8, 352. [Google Scholar] [CrossRef] [Green Version]
- Chaudhary, P.; Shukla, S.; Kumar, I.P.; Namita, I.; Afrin, F.; Sharma, R.K. Radioprotective properties of apple polyphenols: An in vitro study. Mol. Cell. Biochem. 2006, 288, 37–46. [Google Scholar] [CrossRef]
- Haggag, M.G.; Kazem, H.H. Protective Role of Emodin in Reducing the Gamma Rays Induced Hazardous Effects On The Tongue of Diabetic or Normoglycaemic Mice. Isot. Radiat. Res. 2013, 45, 359–373. [Google Scholar]
- Kumar, M.; Samarth, R.; Kumar, M.; Selvan, S.R.; Saharan, B.; Kumar, A. Protective Effect of Adhatoda vascia Nees against Radiation-Induced Damage at Cellular, Biochemical and Chromosomal Levels in Swiss Albino Mice. Evidence-Based Complement. Altern. Med. 2007, 4, 343–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, S.; Singh, M. Histological Alterations in the Spleen of Gamma-Irradiated Mice Induced by Adhatoda vasica Leaf Extract. Int. J. Sci. Res. 2016, 5, 1216–1219. [Google Scholar]
- Krishna, A.; Kumar, A. Evaluation of Radioprotective Effects of Rajgira (Amaranthus paniculatus) Extract in Swiss Albino Mice. J. Radiat. Res. 2005, 46, 233–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, R.K.; Bhatia, A.L.; Sisodia, R. Modulation of Radiation Induced Biochemical Changes in Testis of Swiss Albino Mice by Amaranthus Paniculatus Linn. Asian J. Exp. Sci. 2004, 18, 63–74. [Google Scholar]
- Adamczak, A.; Ożarowski, M. Radioprotective Effects of Plants from the Lamiaceae Family. Anti-Cancer Agents Med. Chem. 2020, 20, 1–23. [Google Scholar] [CrossRef]
- Kumar, A.; Samarth, R.M.; Yasmeen, S.; Sharma, A.; Sugahara, T.; Terado, T.; Kimura, H. Anticancer and radioprotective potentials of Mentha piperita. BioFactors 2004, 22, 87–91. [Google Scholar] [CrossRef]
- Samarth, R.M.; Samarth, M.; Matsumoto, Y. Medicinally important aromatic plants with radioprotective activity. Futur. Sci. OA 2017, 3, FSO247. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.-S.; Endoh, D.; Ishida, Y.; Takahashi, H.; Ozawa, S.; Hayashi, M.; Yabuki, A.; Yamato, O. Radioprotective Effect of Alk(en)yl Thiosulfates Derived from Allium Vegetables against DNA Damage Caused by X-ray Irradiation in Cultured Cells: Antiradiation Potential of Onions and Garlic. Sci. World J. 2012, 2012, 846750. [Google Scholar] [CrossRef] [Green Version]
- Bulle, S.; Reddy, V.D.; Padmavathi, P.; Maturu, P.; Varadacharyulu, N.C. Modulatory role of Pterocarpus santalinus against alcohol-induced liver oxidative/nitrosative damage in rats. Biomed. Pharmacother. 2016, 83, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.E.H.; Kumar, S.S.; Balaji, M.; Maurya, D.K.; Kesavulu, M. Pterocarpus santalinus L. extract mitigates gamma radiation-inflicted derangements in BALB/c mice by Nrf2 upregulation. Biomed. Pharmacother. 2021, 141, 111801. [Google Scholar] [CrossRef]
- Szejk-Arendt, M.; Czubak-Prowizor, K.; Macieja, A.; Poplawski, T.; Olejnik, A.K.; Pawlaczyk-Graja, I.; Gancarz, R.; Zbikowska, H.M. Polyphenolic-polysaccharide conjugates from medicinal plants of Rosaceae/Asteraceae family protect human lymphocytes but not myeloid leukemia K562 cells against radiation-induced death. Int. J. Biol. Macromol. 2020, 156, 1445–1454. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Wang, Z.; Ma, F.; Yang, X.; Cheng, C.; Yao, L. Protective Effect of Anthocyanin from Lonicera caerulea var. edulis on Radiation-Induced Damage in Mice. Int. J. Mol. Sci. 2012, 13, 11773–11782. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Studied Model | Dose | Main Protective Effects | Ref. |
|---|---|---|---|---|
| Apigenin | Human lymphocytes | Up to 25 μg/mL | Protection from 137Cs gamma rays-induced chromosome aberrations | [38] |
| Swiss albino mice | 15 mg/kg BW i.p. for 7 consecutive days | Immunostimulatory effect and mitigation of radiation-induced hematological alterations | [39] | |
| Swiss albino mice | 15 mg/kg BW i.p. for 7 days | Restoration of intestinal crypt-villus architecture Inhibition of the radiation-induced activation of NF-κB expression in the gastrointestinal tissue | [40] | |
| Baicalein | Swiss and C57BL/6 male mice | 10 mg/kg BW for 3 days | Activation of the target molecules ERK and Nrf-2 both in vitro and in vivo | [43] |
| Murine T cell lymphoma cells (EL4) | 5–100 μM | |||
| Swiss albino mice | 150 mg/kg BW | Protection from DNA damage Reduction of radiation-induced damage to mice bone marrow cells | [45] | |
| Human white blood cells | Up to 50 μM | Dose dependent inhibition of DNA strand breaks | ||
| C57BL/6 mice | 5 mg/kg BW for 3 days | Protection against NFκB-mediated inflammatory response through MAPKs and the Akt pathway Up-regulation of FOXO activation, catalase and SOD activities | [46] | |
| Caffeic acid | Mouse colon cancer (CT-26), human liver cancer (HepG2) and human breast cancer (MCF-7) cells lines | 1–3 mM | Induction of apoptosis of colon cancer cell | [65] |
| C57BL/6 mice | 20 mg/kg BW 5 times (every three days) via oral gavage before and 1 after irradiation | Amelioration of ROS production and premature senescence of hematopoietic stem cells | ||
| Caffeic acid phenethyl esters | Sprague-Dawley rats | 10 µmol/kg i.p. 30 min before irradiation | Prevention of oxidative and nitrosative damages induced by radiation | [69] |
| Chlorogenic acid | Human blood lymphocytes | 0.5, 1, 2 and 4 μg/ml | Prevention of genomic instability induced by X-ray irradiation | [67] |
| Mice | 100, 200 and 400 mg/kg BW 1 or 24 h before irradiation | Increase of animal survival (with 200 mg/kg dose 1 h before irradiation with high doses of γ-radiation) | [68] | |
| Curcumin | C57BL/6 mice | standard mouse chow (5015) with 1% or 5% curcumin w/w | Amelioration of radiation-induced pulmonary fibrosis Increase of mouse survival with no impairment of tumor cell killing by radiation. | [74] |
| Wistar albino rats | 100 mg/kg BW orally (by intra gastric intubation) | Protection against intestinal damage | [75] | |
| Wistar rats | 150 mg/kg BW 1 day before irradiation to 3 days after orally | Prevention of radiotherapy-induced oxidative injury to the skin, through enhancing CAT, SOD, and GSH-Px | [78] | |
| Human blood cells | curcumin-encapsulated liposomes (up to 1046.5 µg/mL) | Reduction in the micronuclei frequency No genotoxicity | [80] | |
| Human foreskin fibroblast cells (HHF-2) | conjugated albumin based nanoparticles (50 µg/mL) | Increase in cell viability | [81] | |
| Balb/C mice | 125, 250, 500 and 1000 mg/kg via tail vain | Increase in the survival rate | ||
| Cyanidin-3-O-glucoside (C3G) | HaCaT keratinocytes | 80, 160 and 200 μM | Suppression of COX-2 expression by interaction with the MAPK and Akt signalling pathways | [60] |
| Kunming mice | 125 µM, 250 µM and 500 µM chitosan-C3G nanoparticles; 500 µM C3G | Reduction of UVB-induced epidermal damage through the p53-mediated apoptosis signalling pathway Higher efficiency of nanoparticles respect to C3G at the same dose | [61] | |
| HaCaT keratinocytes | 80, 160 and 200 μM | Intracellular decrease of ROS and of the phospho-p53 and phospho-ATM/ATR levels Expression of anti-apoptotic protein B-cell lymphoma 2 | [59] | |
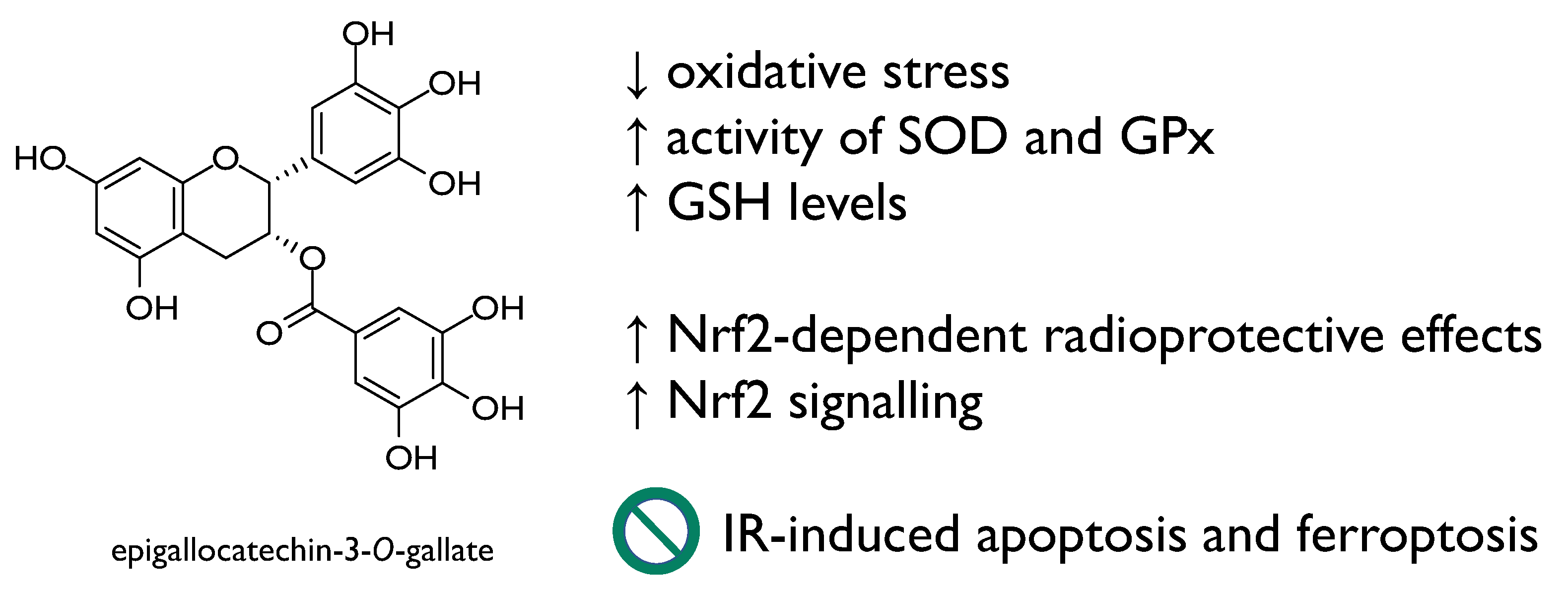
| Epigallocatechin-3-O-gallate | C57 BL/6 J mice | 12.5 or 25 mg/kg BW for 5 days | Reduction of IR-induced cell death in intestinal epithelial cells through Nrf2 Repression of IR-induced apoptosis and ferroptosis Amelioration of intestinal injury induced by total body irradiation | [49] |
| Kunming mice | 6.25, 12.5 and 25 mg/kg BW for 30 days | Prevention of the immune system damage Enhancement of the activity of enzymatic antioxidants (e.g., SOD, GSH-Px) | [50] | |
| Normal fetal lung fibroblasts (MRC5) and adult skin fibroblasts (84BR) Normal human epidermal keratinocytes (NHEK) | Up to 1 mM 250 μM | Prevention against ultraviolet radiation-induced DNA damage | [48] | |
| Ferulic acid | Swiss mice | 50, 75 and 100 mg/kg BW i.p. 1 h before irradiation | Faster repair of DNA | [66] |
| Flavopiridol * | A172/mp53 and HeLa/bcl-2 cells (radioresistant through genetic alteration) | 0.0125–0.125 μM | Radio-sensitization via inhibition of sublethal damage and DNA double-strand breakage repair | [58] |
| Genistein | Human embryo liver cells (L-02) | 1–20 μM | Low concentration: protection against radiation damage High concentration: radiosensitizing features | [52] |
| Huh-7, Hep3B and Hep G2 human HCC cells L-02 cells | 0–40 μM | Low dose (5 μM): HCC cell sensitivity enhancement to X rays; no significant toxicity to L-02 cells | [53] | |
| CD2F1 mice | 100, 200, or 400 mg/kg BW s.c. | Protection against acute radiation injury (administered 24 h before irradiation) Hypothesized indirect mechanism (e.g., cytokine release) | [54] | |
| CD2F1 mice | 150 mg/kg BW i.m. | Selective binding to estrogen receptor β | [55] | |
| Sprague-Dawley rats | 5 mg/kg BW i.p. for 7 days | Up-regulation of ER-β and FOXL-2 Downregulation of TGF-β expression Reversion of radiotherapy-induced premature ovarian failure | [56] | |
| Monoglucosyl rutin * | CHO 10B2 cells | 0.001–0.1% | Increase in survival of IR-treated cells at doses greater than 2 Gy | [32] |
| Naringin | Swiss albino mice | 75 mg/kg BW for 3 days | Reversion of the liver IR-induced redox-imbalance | [41] |
| Quercetin | CBA mice | 100 mg/kg BW i.p. for 3 days | Protection of mice white blood cells from lethal effects and DNA damage before γ-irradiation | [35] |
| Human white blood cells | 50 µM | Protection from DNA damage after γ-irradiation | [34] | |
| Quercetin glycosylated derivatives | Cell-free systems | 10 µM | Reduction of DNA double-strand breaks | [33] |
| Rosmarinic acid | Mice | 100, 200 and 400 mg/kg BW (oral) for 10 days (3 before and 7 after irradiation) | Promotion of the recovery of peripheral blood cells Enhancement of 30-day survival rates | [71] |
| Rutin + podophyllotoxin | C57BL/6 mice | 2.5mg/kg BW 1 h before irradiation | ROS levels reduction Protection of cellular macromolecules Activation of antioxidant signaling pathway | [30] |
| HaCat cells | 0.025–0.4 µg/mL culture medium | Reduction of the cellular damage by scavenging free-radicals, cell-cycle arrest and DNA repair enhancement in the hematopoietic system | [29] | |
| Strain ‘A’ mice | 2.5mg/kg BW i.m. 1 h before irradiation | |||
| VND3207 vanillin derivative * | Human intestinal epithelial cells (HIEC) | 40 µM | Promotion of intestinal repair after radiation injury by regulation of the DNA-PKcs pathway | [64] |
| C57BL/6 J, NOD-SCID (Prkdcscid/scid) and BALB/c mice | 100 mg/kg BW i.g. 30 min before irradiation | |||
| C57BL/6J mice | 100 mg/kg BW (oral gavage) 30 min before/after irradiation | Intestinal repair after radiation injury by reduction of ROS-induced DNA damage and modulation of activated p53 levels in intestinal epithelial cells | [63] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faramarzi, S.; Piccolella, S.; Manti, L.; Pacifico, S. Could Polyphenols Really Be a Good Radioprotective Strategy? Molecules 2021, 26, 4969. https://doi.org/10.3390/molecules26164969
Faramarzi S, Piccolella S, Manti L, Pacifico S. Could Polyphenols Really Be a Good Radioprotective Strategy? Molecules. 2021; 26(16):4969. https://doi.org/10.3390/molecules26164969
Chicago/Turabian StyleFaramarzi, Shadab, Simona Piccolella, Lorenzo Manti, and Severina Pacifico. 2021. "Could Polyphenols Really Be a Good Radioprotective Strategy?" Molecules 26, no. 16: 4969. https://doi.org/10.3390/molecules26164969
APA StyleFaramarzi, S., Piccolella, S., Manti, L., & Pacifico, S. (2021). Could Polyphenols Really Be a Good Radioprotective Strategy? Molecules, 26(16), 4969. https://doi.org/10.3390/molecules26164969