
Photodynamic Inactivation Using Natural Bioactive Compound Prevents and Disrupts the Biofilm Produced by Staphylococcus saprophyticus

 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
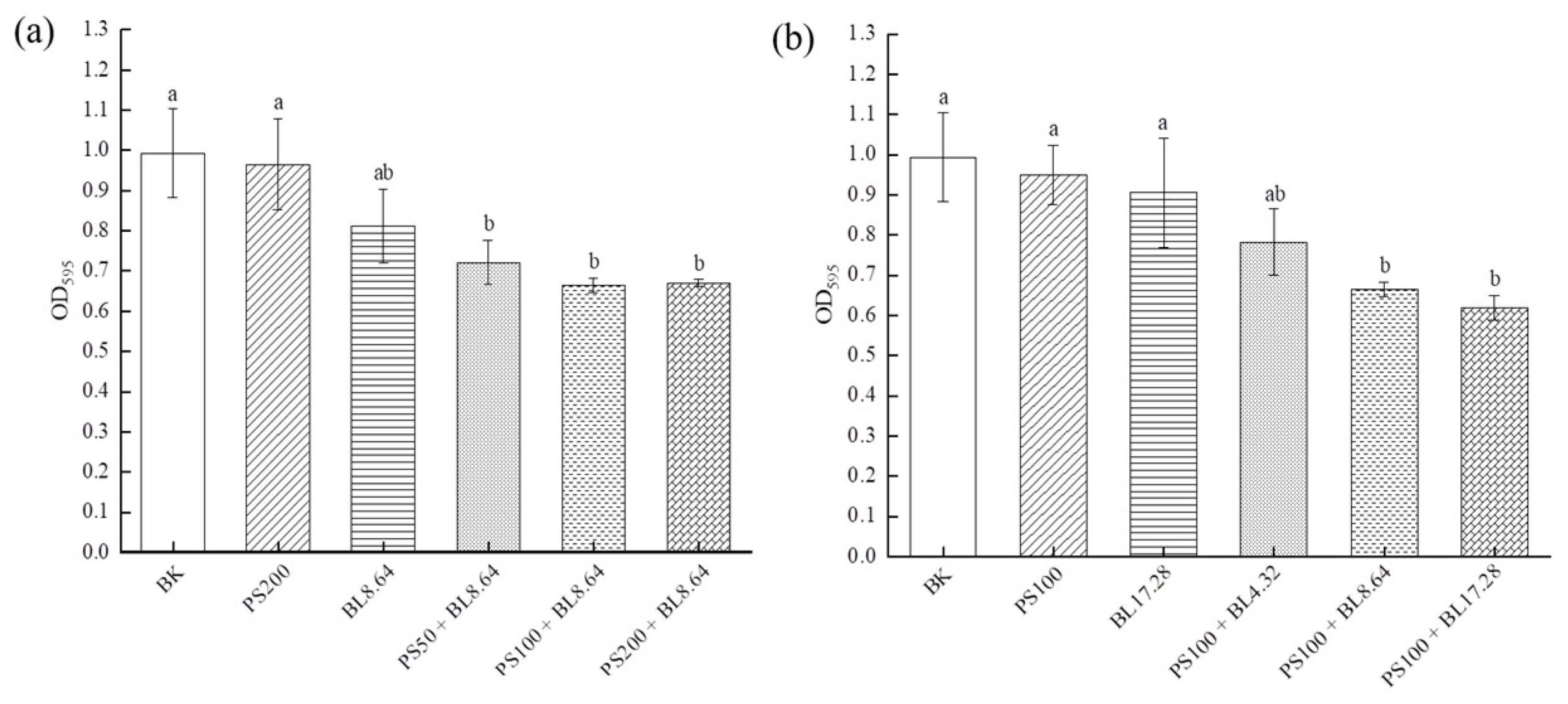
2.1. Effect of PDI on the Prevention of S. saprophyticus Biofilm Formation
2.2. Effect of PDI on the Eradication of Pre-Established S. saprophyticus Biofilm (CFU Assay)
2.3. Effect of PDI on Biofilm Metabolic Activity
2.4. Effect of PDI on Biofilm Membrane Integrity
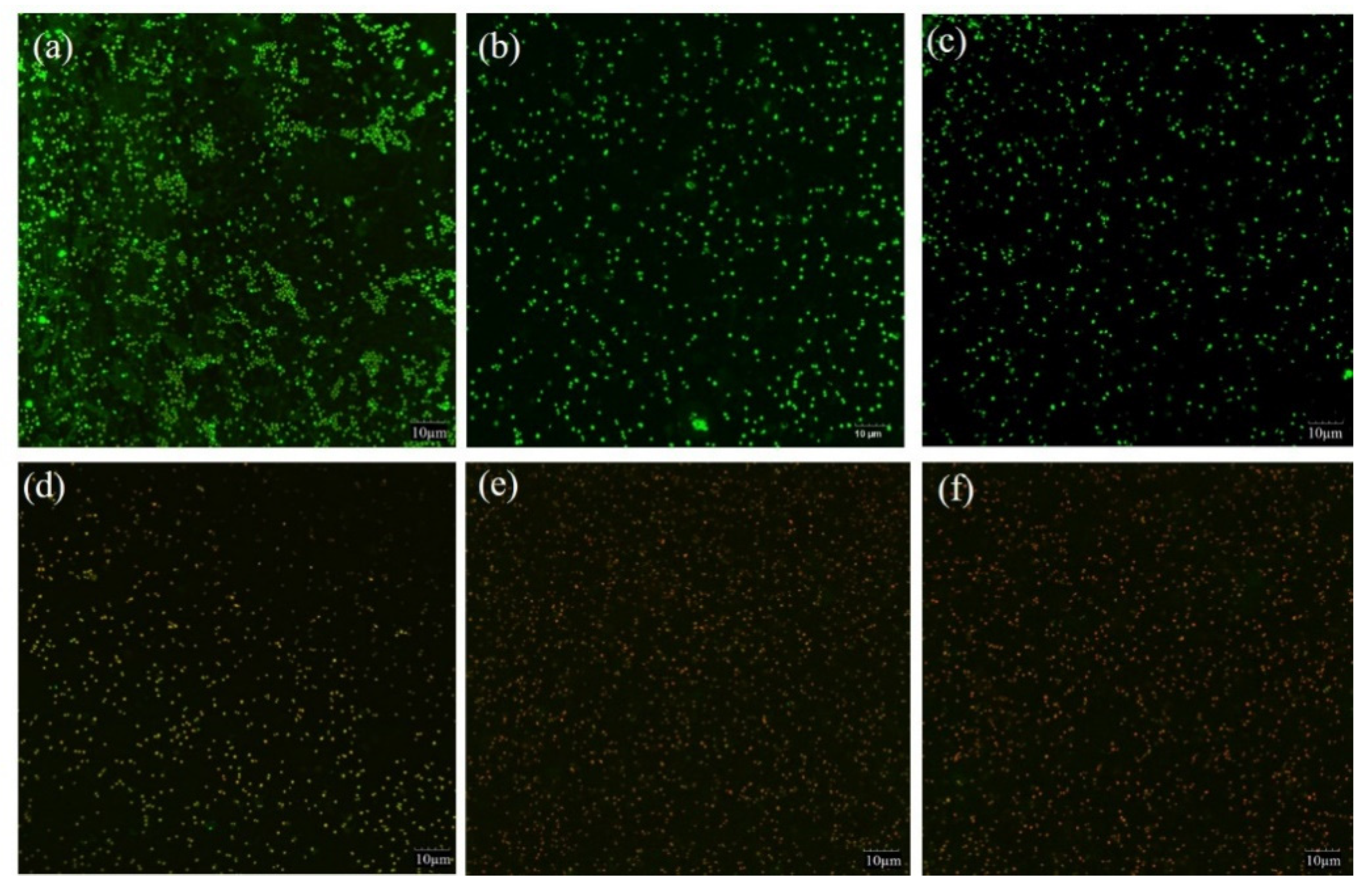
2.5. Morphological Alteration of S. saprophyticus Cells
3. Materials and Methods
3.1. Materials
3.2. Bacterial Strain and Culture Conditions
3.3. Crystal Violet (CV) Staining
3.4. PDI Treatment for Biofilm Prevention
3.5. PDI Treatment for Biofilm Eradication
3.6. Quantification of Biofilm Culturable Cells
3.7. MTT Assay
3.8. CLSM Analysis
3.9. Scanning Electron Microscope (SEM) Analysis for Biofilm Morphology
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Mohamed, E.A.; Marcos, Q.B.; Karola, B.; Sonia, C.A.; Mohamed, A.B.; Rania, M.K.; Abdallah, M.M.; Pilar, C.M.; Jorge, B.V. Molecular characterisation and typing the methicillin resistance of Staphylococcus spp. isolated from raw milk and cheeses in northwest Spain: A mini survey. Int. Dairy J. 2019, 89, 68–76. [Google Scholar]
- Harakeh, S.; Yassine, H.; Hajjar, S.; El-Fadel, M. Isolates of Staphylococcus aureus and saprophyticus resistant to antimicrobials isolated from the Lebanese aquatic environment. Mar. Pollut. Bull. 2006, 52, 912–919. [Google Scholar] [CrossRef]
- Casaes Nunes, R.S.; Pires de Souza, C.; Pereira, K.S.; Del Aguila, E.M.; Flosi Paschoalin, V.M. Identification and molecular phylogeny of coagulasenegative staphylococci isolates from Minas Frescal cheese in southeastern Brazil: Superantigenic toxin production and antibiotic resistance. J. Dairy Sci. 2016, 99, 2641–2653. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Qiao, M.; Zhou, Y.; Du, H.; Bai, J.; Yuan, W.; Liu, J.; Wang, D.; Hu, Y.; Wu, Y. In vitro synergistic effect of baicalin with azithromycin against Staphylococcus saprophyticus isolated from francolins with ophthalmia. Poult. Sci. 2019, 98, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Martins, K.B.; Ferreira, A.M.; Pereira, V.C.; Pinheiro, L.; Oliveira, A.; Cunha, M.L.R.S. In vitro effects of antimicrobial agents on planktonic and biofilm forms of Staphylococcus saprophyticus isolated from patients with urinary tract infections. Front. Microbiol. 2019, 10, 40. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, Y.D.; Lamarche, D.; Chever, P.; Haine, D.; Messier, S.; Jacques, M. Characterization of the ability of coagulase-negative staphylococci isolated from the milk of Canadian farms to form biofilms. J. Dairy Sci. 2013, 96, 234–246. [Google Scholar] [CrossRef]
- Shirtliff, M.E.; Mader, J.T.; Camper, A.K. Molecular interactions in biofilms. Chem. Biol. 2000, 9, 859–871. [Google Scholar] [CrossRef]
- Goldberg, J. Biofilms and antibiotic resistance: A genetic linkage. Trends Microbiol. 2002, 10, 264. [Google Scholar] [CrossRef]
- Szabados, F.; Mohner, A.; Kleine, B.; Gatermann, S.G. Staphylococcus saprophyticus surface-associated protein (Ssp) is associated with lifespan reduction in Caenorhabditis elegans. Virulence 2013, 4, 604–611. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wassenaar, T.M.; Ussery, D.; Nielsen, L.N.; Ingmer, H. Review and phylogenetic analysis of qac genes that reduce susceptibility to quaternary ammonium compounds in Staphylococcus species. Eur. J. Microbiol. Immunol. 2015, 5, 44–61. [Google Scholar] [CrossRef] [PubMed]
- Von Eiff, C.; Peters, G.; Heilmann, C. Pathogenesis of infections due to coagulase-negative staphylococci. Lancet Infect Dis. 2002, 2, 677–685. [Google Scholar] [CrossRef]
- Castro, K.A.; Moura, N.M.; Fernandes, A.; Faustino, M.A.; Simões, M.M.; Cavaleiro, J.A.; Nakagaki, S.; Almeida, A.; Cunha, Â.; Silvestre, A.J. Control of Listeria innocua biofilms by biocompatible photodynamic antifouling chitosan based materials. Dyes Pigments 2017, 137, 265–276. [Google Scholar] [CrossRef]
- Gundogan, N.; Citak, S.; Yucel, N.; Devren, A. A note on the incidence and antibiotic resistance of Staphylococcus aureus isolated from meat and chicken samples. Meat Sci. 2005, 69, 807–810. [Google Scholar] [CrossRef]
- Chang, S.C.; Hsieh, S.M.; Chen, M.L.; Sheng, W.H.; Chen, Y.C. Oral fusidic acid fails to eradicate methicillin-resistant Staphylococcus aureus colonization and results in emergence of fusidic acid-resistant strains. Diagn. Microbiol. Infect Dis. 2000, 36, 131–136. [Google Scholar] [CrossRef]
- Leonel, L.D.C.; Carvalho, M.L.; Da Silva, B.M.; Zamuner, S.; Alberto-Silva, C.; Costa, M.S. Photodynamic Antimicrobial Chemotherapy (PACT) using methylene blue inhibits the viability of the biofilm produced by Candida albicans. Photodiagnosis Photodyn. Ther. 2019, 26, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Baltazar, L.M.; Ray, A.; Santos, D.A.; Cisalpino, P.S.; Friedman, A.J.; Nosanchuk, J.D. Antimicrobial photodynamic therapy: An effective alternative approach to control fungal infections. Front. Microbiol. 2015, 6, 202. [Google Scholar] [CrossRef] [PubMed]
- Tortik, N.; Spaeth, A.; Plaetzer, K. Photodynamic decontamination of foodstuff from Staphylococcus aureus based on novel formulations of curcumin. Photochem. Photobiol. Sci. 2014, 13, 1402–1409. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Jia, Y.T.; Zhang, M. Effect of curcumin on the quality properties of millet fresh noodle and its inhibitory mechanism against the isolated spoilage bacteria. Food Sci. Nutr. 2020, 8, 1451–1460. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, A.P.D.; Pavarina, A.C.; Dovigo, L.N.; Brunetti, I.L.; Bagnato, V.S.; Vergani, C.E.; de Souza Costa, C.A. Phototoxic effect of curcumin on methicillin-resistant Staphylococcus aureus and L929 fibroblasts. Lasers Med. Sci. 2013, 28, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Araújo, N.; Fontana, C.; Bagnato, V.; Gerbi, M. Photodynamic antimicrobial therapy of curcumin in biofilms and carious dentine. Lasers Med. Sci. 2014, 29, 629–635. [Google Scholar] [CrossRef] [PubMed]
- Dovigo, L.N.; Carmello, J.C.; de Souza Costa, C.A.; Vergani, C.E.; Brunetti, I.L.; Bagnato, V.S.; Pavarina, A.C. Curcumin-mediated photodynamic inactivation of Candida albicans in a murine model of oral candidiasis. Med. Mycol. 2013, 51, 243–251. [Google Scholar] [CrossRef]
- Hegge, A.B.; Bruzell, E.; Kristensen, S.; Tønnesen, H. Photoinactivation of Staphylococcus epidermidis biofilms and suspensions by the hydrophobic photosensitizer curcumin-effect of selected nanocarrier: Studies on curcumin and curcuminoides XLVII. Eur. J. Pharm. Sci. 2012, 47, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Paschoal, M.A.; Tonon, C.C.; Spolidório, D.M.; Bagnato, V.S.; Giusti, J.S.; Santos-Pinto, L. Photodynamic potential of curcumin and blue LED against Streptococcus mutans in a planktonic culture. Photodiagn. Photodyn. Ther. 2013, 10, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Sanitá, P.V.; Pavarina, A.C.; Dovigo, L.N.; Ribeiro, A.P.D.; Andrade, M.C.; Mima, E.G.D.O. Curcumin-mediated anti-microbial photodynamic therapy against Candida dubliniensis biofilms. Lasers Med. Sci. 2018, 33, 709–717. [Google Scholar] [CrossRef]
- Huang, J.; Chen, B.; Li, H.; Zeng, Q.H.; Wang, J.J.; Liu, H.; Pan, Y.; Zhao, Y. Enhanced antibacterial and antibiofilm functions of the curcumin-mediated photodynamic inactivation against Listeria monocytogenes. Food Control 2020, 108, 106886. [Google Scholar] [CrossRef]
- Chen, B.; Huang, J.; Li, H.; Zeng, Q.H.; Wang, J.J.; Liu, H.; Pan, Y.; Zhao, Y. Eradication of planktonic Vibrio parahaemolyticus and its sessile biofilm by curcumin-mediated photodynamic inactivation. Food Control 2020, 113, 107181. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Jia, Y.T.; Li, W.Y.; Zhang, M. Antimicrobial photodynamic inactivation with curcumin against Staphylococcus saprophyticus, in vitro and on fresh dough sheet. LWT Food Sci. Technol. 2021, 147, 111567. [Google Scholar] [CrossRef]
- Mah, T.F.; O’Toole, G.A. Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol. 2001, 9, 34–39. [Google Scholar] [CrossRef]
- Galdiero, E.; de Alteriis, E.; De Natale, A.; D’Alterio, A.; Siciliano, A.; Guida, M.; Lombardi, L.; Falanga, A.; Galdiero, S. Eradication of Candida albicans persister cell biofilm by the membranotropic peptide gH625. Sci. Rep. 2020, 10, 5780. [Google Scholar] [CrossRef] [PubMed]
- Lima, S.L.; Colombo, A.L.; de Almeida Junior, J.N. Fungal cell wall: Emerging antifungals and drug resistance. Front. Microbiol. 2019, 10, 2573. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, C.G.D.S.; Sanitá, P.V.; Ribeiro, A.P.D.; Dias, L.M.; Jorge, J.H.; Pavarina, A.C. Antimicrobial photodynamic therapy effectiveness against susceptible and methicillin-resistant Staphylococcus aureus biofilms. Photodiagnosis Photodyn. Ther. 2020, 30, 101760. [Google Scholar] [CrossRef] [PubMed]
- Quishida, C.C.; Mima, E.G.O.; Jorge, J.H.; Vergani, C.E.; Bagnato, V.S.; Pavarina, A.C. Photodynamic inactivation of a multispecies biofilm using curcumin and LED light. Lasers Med. Sci. 2016, 31, 997–1009. [Google Scholar] [CrossRef] [PubMed]
- Stepanovic, S.; Vukovic, D.; Dakic, I.; Savic, B.; Svabic-Vlahovic, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Vassena, C.; Fenu, S.; Giuliani, F.; Fantetti, L.; Roncucci, G.; Simonutti, G.; Romanò, C.L.; De Francesco, R.; Drago, L. Photodynamic antibacterial and antibiofilm activity of RLP068/Cl against Staphylococcus aureus and Pseudomonas aeruginosa forming biofilms on prosthetic material. Int. J. Antimicrob. Agents 2014, 44, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Vilela, S.F.G.; Junqueira, J.C.; Barbosa, J.O.; Majewski, M.; Munin, E.; Jorge, A.O.C. Photodynamic inactivation of Staphylococcus aureus and Escherichia coli biofilms by malachite green and phenothiazine dyes: An in vitro study. Arch. Oral Biol. 2012, 57, 704–710. [Google Scholar] [CrossRef]
- Liu, J.; Wang, Y.; Hao, Y.M.; Wang, Z.W.; Yang, Z.H.; Wang, Z.Y.; Wang, J. 5-heptadecylresorcinol attenuates oxidative damage and mitochondria-mediated apoptosis through activation of SIRT3/FOXO3a signaling pathway in neurocytes. Food Funct. 2020, 11, 2535–2542. [Google Scholar] [CrossRef] [PubMed]
- Garcez, A.S.; Núnez, S.C.; Azambuja Jr, N.; Fregnani, E.R.; Rodriguez, H.M.; Hamblin, M.R.; Suzuki, H.; Ribeiro, M.S. Effects of photodynamic therapy on Gram-positive and Gram-negative bacterial biofilms by bioluminescence imaging and scanning electron microscopic analysis. Photomed. Laser. Surg. 2013, 31, 519–525. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, W.; Wang, Z.; Li, Q.; Jia, Y.; Song, S.; Ma, Z.; Liu, J.; Wang, J. Photodynamic Inactivation Using Natural Bioactive Compound Prevents and Disrupts the Biofilm Produced by Staphylococcus saprophyticus. Molecules 2021, 26, 4713. https://doi.org/10.3390/molecules26164713
Yang W, Wang Z, Li Q, Jia Y, Song S, Ma Z, Liu J, Wang J. Photodynamic Inactivation Using Natural Bioactive Compound Prevents and Disrupts the Biofilm Produced by Staphylococcus saprophyticus. Molecules. 2021; 26(16):4713. https://doi.org/10.3390/molecules26164713
Chicago/Turabian StyleYang, Wei, Ziyuan Wang, Qing Li, Yating Jia, Shuimiao Song, Zichu Ma, Jie Liu, and Jing Wang. 2021. "Photodynamic Inactivation Using Natural Bioactive Compound Prevents and Disrupts the Biofilm Produced by Staphylococcus saprophyticus" Molecules 26, no. 16: 4713. https://doi.org/10.3390/molecules26164713
APA StyleYang, W., Wang, Z., Li, Q., Jia, Y., Song, S., Ma, Z., Liu, J., & Wang, J. (2021). Photodynamic Inactivation Using Natural Bioactive Compound Prevents and Disrupts the Biofilm Produced by Staphylococcus saprophyticus. Molecules, 26(16), 4713. https://doi.org/10.3390/molecules26164713