
The Diversity of the Mitochondrial Outer Membrane Protein Import Channels: Emerging Targets for Modulation


Abstract
1. Introduction
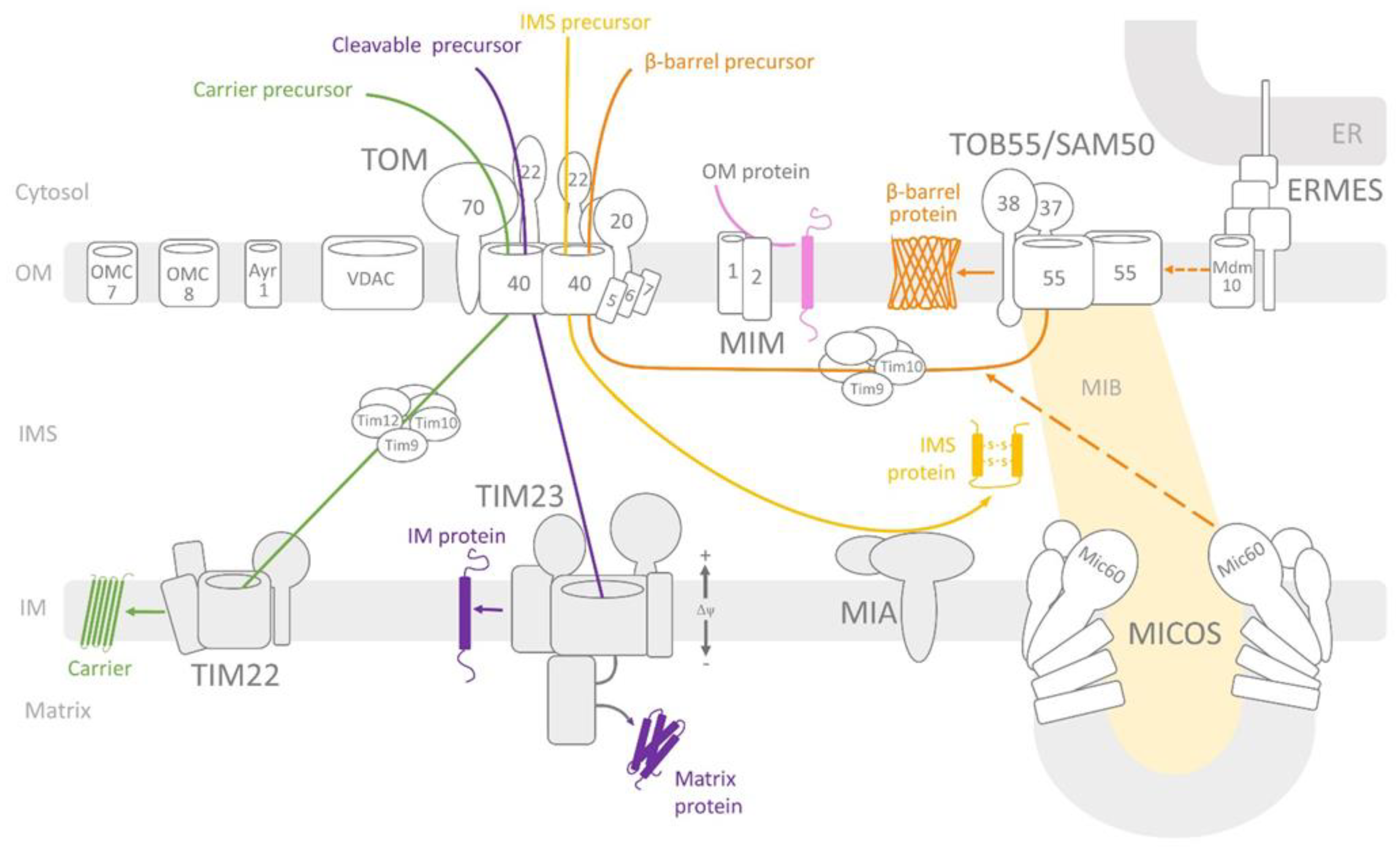
2. Overview of the Mitochondrial Protein Import Machinery
Protein Import across and into the Mitochondrial Outer Membrane
3. Structure and Properties of the Outer Membrane Protein Import Channels
3.1. Tom40 Channel
3.2. Tob55/Sam50 Channel
3.3. Mdm10 Channel
3.4. Mim1 Channel
3.5. Mitochondrial Apoptosis-Induced Channel (MAC)
4. Non-Proteinaceous Molecules Transported by Protein Import Channels
4.1. Transport of RNA
4.2. Transport of Metabolites
4.3. Ayr1 and OMC7 and OMC8: Mitochondrial Outer Membrane Channels for Unknown Molecules
5. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Kutik, S.; Guiard, B.; Meyer, H.E.; Wiedemann, N.; Pfanner, N. Cooperation of translocase complexes in mitochondrial protein import. J. Cell Bio. 2007, 179, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Wiedemann, N.; Pfanner, N. Mitochondrial machineries for protein import and assembly. Annu. Rev. Biochem. 2017, 86, 685–714. [Google Scholar] [CrossRef] [PubMed]
- Pfanner, N.; Warscheid, B.; Wiedemann, N. Mitochondrial proteins: From biogenesis to functional networks. Nat. Rev. Mol. Cell Biol. 2019, 20, 267–284. [Google Scholar] [CrossRef] [PubMed]
- Kruger, V.; Becker, T.; Becker, L.; Montilla-Martinez, M.; Ellenrieder, L.; Vögtle, F.N.; Meyer, H.E.; Ryan, M.T.; Wiedemann, N.; Warscheid, B.; et al. Identification of new channels by systematic analysis of the mitochondrial outer membrane. J. Cell Biol. 2017, 216, 3485–3495. [Google Scholar] [CrossRef] [PubMed]
- Checchetto, V.; Szabo, I. Novel channels of the outer membrane of mitochondria: Recent discoveries change our view. Bioessays 2018, 40, e1700232. [Google Scholar] [CrossRef]
- Endo, T.; Yamano, K. Multiple pathways for mitochondrial protein trafc. Biol. Chem. 2009, 390, 723–730. [Google Scholar] [CrossRef] [PubMed]
- Kreimendahl, S.; Rassow, J. The mitochondrial outer membrane protein Tom70-mediator in protein traffic, membrane contact sites and innate immunity. Int. J. Mol. Sci. 2020, 21, 7262. [Google Scholar] [CrossRef]
- Paschen, S.A.; Waizenegger, T.; Stan, T.; Preuss, M.; Cyrklaff, M.; Hell, K.; Rapaport, D.; Neupert, W. Evolutionary conservation of biogenesis of β-barrel membrane proteins. Nature 2003, 426, 862–866. [Google Scholar] [CrossRef] [PubMed]
- Habib, S.J.; Waizenegger, T.; Niewienda, A.; Paschen, S.A.; Neupert, W.; Rapaport, D. The N-terminal domain of Tob55 has a receptor-like function in the biogenesis of mitochondrial β-barrel proteins. J. Cell Biol. 2007, 176, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Zeth, K. Structure and evolution of mitochondrial outer membrane proteins of β-barrel topology. Biochim. Biophys. Acta 2010, 1797, 1292–1299. [Google Scholar] [CrossRef]
- Misra, R. Assembly of the β-barrel outer membrane proteins in Gram-negative bacteria, mitochondria, and chloroplasts. ISRN Mol. Biol. 2012, 2012, 708203. [Google Scholar] [CrossRef]
- Popov-Celeketic, J.; Waizenegger, T.; Rapaport, D. Mim1 functions in an oligomeric form to facilitate the integration of Tom20 into the mitochondrial outer membrane. J. Mol. Biol. 2008, 376, 671–680. [Google Scholar] [CrossRef]
- Lueder, F.; Lithgow, T. The three domains of the mitochondrial outer membrane protein Mim1 have discrete functions in assembly of the TOM complex. FEBS Lett. 2009, 583, 1475–1480. [Google Scholar] [CrossRef]
- Ellenrieder, L.; Opalinski, L.; Becker, L.; Kruger, V.; Mirus, O.; Straub, S.P.; Ebell, K.; Flinner, N.; Stiller, S.B.; Guiard, B.; et al. Separating mitochondrial protein assembly and endoplasmic reticulum tethering by selective coupling of Mdm10. Nat. Commun. 2016, 7, 13021. [Google Scholar] [CrossRef]
- Chacinska, A.; Koehler, C.M.; Milenkovic, D.; Lithgow, T.; Pfanner, N. Importing mitochondrial proteins: Machineries and mechanisms. Cell 2009, 138, 628–644. [Google Scholar] [CrossRef]
- Truscott, K.N.; Kovermann, P.; Geissler, A.; Merlin, A.; Meijer, M.; Driessen, A.J.; Rassow, J.; Pfanner, N.; Wagner, R. A presequence- and voltage-sensitive channel of the mitochondrial preprotein translocase formed by Tim23. Nat. Struct. Biol. 2001, 8, 1074–1082. [Google Scholar] [CrossRef]
- Sirrenberg, C.; Bauer, M.F.; Guiard, B.; Neupert, W.; Brunner, M. Import of carrier proteins into the mitochondrial inner membrane mediated by Tim22. Nature 1996, 384, 582–585. [Google Scholar] [CrossRef] [PubMed]
- Callegari, S.; Richter, F.; Chojnacka, K.; Jans, D.C.; Lorenzi, I.; Pacheu-Grau, D.; Jakobs, S.; Lenz, C.; Urlaub, H.; Dudek, J.; et al. TIM29 is a subunit of the human carrier translocase required for protein transport. FEBS Lett. 2016, 590, 4147–4158. [Google Scholar] [CrossRef]
- Callegari, S.; Cruz-Zaragoza, L.D.; Rehling, P. From TOM to the TIM23 complex-handing over of a precursor. Biol. Chem. 2020, 401, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, M.; Darden, C.; Gonzalez, F.S.; Singha, U.K.; Quinones, L.; Tripathi, A. Tim17 updates: A comprehensive review of an ancient mitochondrial protein translocator. Biomolecules 2020, 10, 1643. [Google Scholar] [CrossRef] [PubMed]
- Neupert, W. A perspective on transport of proteins into mitochondria: A myriad of open questions. J. Mol. Biol. 2015, 427, 1135–1158. [Google Scholar] [CrossRef] [PubMed]
- Doan, K.N.; Ellenrieder, L.; Becker, T. Mitochondrial porin links protein biogenesis to metabolism. Curr. Genet. 2019, 65, 899–903. [Google Scholar] [CrossRef]
- Sirrenberg, C.; Endres, M.; Folsch, H.; Stuart, R.A.; Neupert, W.; Brunner, M. Carrier protein import into mitochondria mediated by the intermembrane proteins Tim10/Mrs11 and Tim12/Mrs5. Nature 1998, 391, 912–915. [Google Scholar] [CrossRef]
- Adam, A.; Endres, M.; Sirrenberg, C.; Lottspeich, F.; Neupert, W.; Brunner, M. Tim9, a new component of the TIM22.54 translocase in mitochondria. EMBO J. 1999, 18, 313–319. [Google Scholar] [CrossRef]
- Kozjak, V.; Wiedemann, N.; Milenkovic, D.; Lohaus, C.; Meyer, H.E.; Guiard, B.; Meisinger, C.; Pfanner, N. An Essential role of Sam50 in the protein sorting and assembly machinery of the mitochondrial outer membrane. J. Biol. Chem. 2003, 278, 49. [Google Scholar] [CrossRef]
- Wiedemann, N.; Kozjak, V.; Chacinska, A.; Schonfisch, B.; Rospert, S.; Ryan, M.T.; Pfanner, N.; Meisinger, C. Machinery for protein sorting and assembly in the mitochondrial outer membrane. Nature 2003, 424, 565–571. [Google Scholar] [CrossRef]
- Waizenegger, T.; Habib, S.J.; Lech, M.; Mokranjac, D.; Paschen, S.A.; Hell, K.; Neupert, W.; Rapaport, D. Tob38, a novel essential component in the biogenesis of betabarrel proteins of mitochondria. EMBO Rep. 2004, 5, 704709. [Google Scholar] [CrossRef]
- Klein, A.; Israel, L.; Lackey, S.W.K.; Nargang, F.E.; Imhof, A.; Baumeister, W.; Neupert, W.; Thomas, D.R. Characterization of the insertase for β-barrel proteins of the outer mitochondrial membrane. J. Cell Biol. 2012, 99, 599–611. [Google Scholar] [CrossRef] [PubMed]
- Diederichs, K.A.; Ni, X.; Rollauer, S.E.; Botos, I.; Tan, X.; King, M.S.; Kunji, E.R.S.; Jiang, J.; Buchanan, S.K. Structural insight into mitochondrial β-barrel outer membrane protein biogenesis. Nat. Commun. 2020, 11, 3290. [Google Scholar] [CrossRef]
- Kozjak-Pavlovic, V.; Ross, K.; Benlasfer, N.; Kimmig, S.; Karlas, A.; Rudel, T. Conserved roles of Sam50 and metaxins in VDAC biogenesis. EMBO Rep. 2007, 8, 576–582. [Google Scholar] [CrossRef] [PubMed]
- Xiea, J.; Marusichb, M.F.; Soudac, P.; Whiteleggec, J.; Capaldi, R.A. The mitochondrial inner membrane protein Mitofilin exists as a complex with SAM50, metaxins 1 and 2, coiled-coil-helix domain-containing protein 3 and 6 and DnaJC11. FEBS Lett. 2007, 581, 3545–3549. [Google Scholar] [CrossRef]
- Cartron, P.F.; Petit, E.; Bellot, G.; Oliver, L.; Vallette, F.M. Metaxins 1 and 2, two proteins of the mitochondrial protein sorting and assembly machinery, are essential for Bak activation during TNF alpha triggered apoptosis. Cell. Signal. 2014, 26, 1928–1934. [Google Scholar] [CrossRef]
- Buczek, D.; Wojtkowska, M.; Suzuki, Y.; Sonobe, S.; Nishigami, Y.; Antoniewicz, M.; Kmita, H.; Makałowski, W. Protein import complexes in the mitochondrial outer membrane of Amoebozoa representatives. BMC Genom. 2016, 17, 99. [Google Scholar] [CrossRef]
- Wojtkowska, M.; Buczek, D.; Suzuki, Y.; Shabardina, V.; Makałowski, W.; Kmita, H. The emerging picture of the mitochondrial protein import complexes of Amoebozoa supergroup. BMC Genom. 2017, 18, 997. [Google Scholar] [CrossRef]
- Mazur, M.; Wojtkowska, M.; Skalski, M.; Słocińska, M.; Kmita, H. The TOB/SAM complex composition in mitochondria of Dictyostelium discoideum during progression from unicellularity to multicellularity. Acta Biochim. Pol. 2019, 66, 551–557. [Google Scholar] [CrossRef]
- Kornera, C.; Barrerac, M.; Dukanovice, J.; Eydt, K.; Harner, M.; Rabla, R.; Vogelf, F.; Rapaporte, D.; Neupert, W.; Reichert, A.S. The C-terminal domain of Fcj1 is required for formation of crista junctions and interacts with the TOB/SAM complex in mitochondria. Mol. Biol. Cell 2012, 23, 2143–2155. [Google Scholar] [CrossRef]
- Kozjak-Pavlovic, V. The MICOS complex of human mitochondria. Cell Tissue Res. 2017, 367, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Horvath, S.E.; Rampelt, H.; Oeljeklaus, S.; Warscheid, B.; van der Laan, M.; Pfanner, N. Role of membrane contact sites in protein import into mitochondria. Protein Sci. 2015, 24, 277–297. [Google Scholar] [CrossRef] [PubMed]
- Dekker, P.J.; Martin, F.; Maarse, A.C.; Bömer, U.; Muller, H.; Guiard, B.; Meijer, M.; Rassow, J.; Pfanner, N. The Tim core complex defines the number of mitochondrial translocation contact sites and can hold arrested preproteins in the absence of matrix Hsp70-Tim44. EMBO J. 1997, 16, 5408–5419. [Google Scholar] [CrossRef] [PubMed]
- Hill, K.; Model, K.; Ryan, M.T.; Dietmeier, K.; Martin, F.; Wagner, R.; Pfanner, N. Tom40 forms the hydrophilic channel of the mitochondrial import pore for preproteins. Nature 1998, 395, 516–521. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Okazawa, Y.; Komiya, T.; Saeki, K.; Mekada, E.; Kitada, S.; Ito, A.; Mihara, K. Characterization of rat TOM40, a central component of the preprotein translocase of the mitochondrial outer membrane. J. Biol. Chem. 2000, 275, 37930–37936. [Google Scholar] [CrossRef] [PubMed]
- Meisinger, C.; Ryan, M.T.; Hill, K.; Model, K.; Lim, J.H.; Sickmann, A.; Müller, H.; Meyer, H.E.; Wagner, R.; Pfanner, N. Protein import channel of the outer mitochondrial membrane: A highly stable Tom40-Tom22 core structure differentially interacts with preproteins, small tom proteins, and import receptors. Mol. Cell. Biol. 2001, 21, 2337–2348. [Google Scholar] [CrossRef] [PubMed]
- Kreimendahl, S.; Schwichtenberg, J.; Günnewig, K.; Brandherm, L.; Rassow, J. The selectivity filter of the mitochondrial protein import machinery. BMC Biol. 2020, 18, 156. [Google Scholar] [CrossRef] [PubMed]
- Harner, M.; Neupert, W.; Deponteb, M. Lateral release of proteins from the TOM complex into the outer membrane of mitochondria. EMBO J. 2011, 30, 3232–3241. [Google Scholar] [CrossRef]
- Wojtkowska, M.; Jakalski, M.; Pienkowska, J.R.; Stobienia, O.; Karachitos, A.; Przytycka, T.M.; Weiner, J., 3rd; Kmita, H.; Makalowski, W. Phylogenetic analysis of mitochondrial outer membrane β-barrel channels. Genome Biol. Evol. 2012, 4, 110–125. [Google Scholar] [CrossRef]
- Morgenstern, M.; Stiller, S.B.; Lubbert, P.; Peikert, C.D.; Dannenmaier, S.; Drepper, F.; Weill, U.; Hoß, P.; Feuerstein, R.; Gebert, M.; et al. Definition of a high- confidence mitochondrial proteome at quantitative scale. Cell Rep. 2017, 19, 2836–2852. [Google Scholar] [CrossRef]
- Ahting, U.; Thun, C.; Hegerl, R.; Typke, D.; Nargang, F.E.; Neupert, W.; Nussberger, S. The TOM core complex: The general protein import pore of the outer membrane of mitochondria. J. Cell Biol. 1999, 147, 959–968. [Google Scholar] [CrossRef]
- Shiota, T.; Imai, K.; Qiu, J.; Hewitt, V.L.; Tan, K.; Shen, H.H.; Sakiyama, N.; Fukasawa, Y.; Hayat, S.; Kamiya, M.; et al. Molecular architecture of the active mitochondrial protein gate. Science 2015, 349, 1544–1548. [Google Scholar] [CrossRef]
- Model, K.; Meisinger, C.; Prinz, T.; Wiedemann, N.; Truscott, K.N.; Pfanner, N.; Ryan, M.T. Multistep assembly of the protein import channel of the mitochondrial outer membrane. Nat. Struct. Biol. 2001, 8, 361–370. [Google Scholar] [CrossRef]
- Gold, V.A.; Brandt, T.; Cavellini, L.; Cohen, M.M.; Ieva, R.; van der Laan, M. Analysis of mitochondrial membrane protein complexes by electron cryo-tomography. Methods Mol. Biol. 2017, 1567, 315–336. [Google Scholar] [PubMed]
- Bausewein, T.; Naveed, H.; Liang, J.; Nussberger, S. The structure of the TOM core complex in the mitochondrial outer membrane. Biol. Chem. 2020, 401, 687–697. [Google Scholar] [CrossRef]
- Araiso, Y.; Tsutsumi, A.; Qiu, J.; Imai, K.; Shiota, T.; Song, J.; Lindau, C.; Wenz, L.S.; Sakaue, H.; Yunoki, K.; et al. Structure of the mitochondrial import gate reveals distinct preprotein paths. Nature 2019, 575, 395–401. [Google Scholar] [CrossRef]
- Wang, W.; Chen, X.; Zhang, L.; Yi, J.; Ma, Q.; Yin, J.; Zhuo, W.; Gu, J.; Yang, M. Atomic structure of human TOM core complex. Cell Discov. 2020, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tucker, K.; Park, E. Cryo-EM structure of the mitochondrial protein-import channel TOM complex at near-atomic resolution. Nat. Struct. Mol. Biol. 2019, 26, 1158–1166. [Google Scholar] [CrossRef] [PubMed]
- Becker, T.; Horvath, S.E.; Bottinger, L.; Gebert, N.; Daum, G.; Pfanner, N. Role of phosphatidylethanolamine in the biogenesis of mitochondrial outer membrane proteins. J. Biol. Chem. 2013, 288, 16451–16459. [Google Scholar] [CrossRef] [PubMed]
- Bausewein, T.; Mills, D.J.; Langer, J.D.; Nitschke, B.; Nussberger, S.; Kuhlbrandt, W. Cryo-EM structure of the TOM core complex from Neurospora crassa. Cell 2017, 170, 693–700. [Google Scholar] [CrossRef]
- Kuszak, A.J.; Jacobs, D.; Gurnev, P.A.; Shiota, T.; Louis, J.M.; Lithgow, T.; Bezrukov, S.M.; Rostovtseva, T.K.; Buchanan, S.K. Evidence of distinct channel conformations and substrate binding affinities for the mitochondrial outer membrane protein translocase pore Tom40. J. Biol. Chem. 2015, 290, 26204–26217. [Google Scholar] [CrossRef]
- Becker, L.; Bannwarth, M.; Meisinger, C.; Hill, K.; Model, K.; Krimmer, T.; Casadio, R.; Truscott, K.N.; Schulz, G.E.; Pfanner, N.; et al. Preprotein translocase of the outer mitochondrial membrane: Reconstituted Tom40 forms a characteristic TOM pore. J. Mol. Biol. 2005, 353, 1011–1020. [Google Scholar] [CrossRef]
- Wojtkowska, M.; Buczek, D.; Stobienia, O.; Karachitos, A.; Antoniewicz, M.; Slocinska, M.; Makalowski, W.; Kmita, H. The TOM complex of amoebozoans: The cases of the amoeba Acanthamoeba castellanii and the slime mold Dictyostelium discoideum. Protist 2015, 166, 349–362. [Google Scholar] [CrossRef]
- Poynor, M.; Eckert, R.; Nussberger, S. Dynamics of the preprotein translocation channel of the outer membrane of mitochondria. Biophys. J. 2008, 95, 1511–1522. [Google Scholar] [CrossRef] [PubMed]
- Stan, T.; Ahting, U.; Dembowski, M.; Kunkele, K.P.; Nussberger, S.; Neupert, W.; Rapaport, D. Recognition of preproteins by the isolated TOM complex of mitochondria. EMBO J. 2000, 19, 4895–4902. [Google Scholar] [CrossRef]
- Abe, Y.; Shodai, T.; Muto, T.; Mihara, K.; Torii, H.; Nishikawa, S.; Endo, T.; Kohda, D. Structural basis of presequence recognition by the mitochondrial protein import receptor Tom20. Cell 2000, 100, 551–560. [Google Scholar] [CrossRef]
- Niemann, M.; Harsman, A.; Mani, J.; Peikert, C.D.; Oeljeklaus, S.; Warscheid, B.; Wagner, R.; Schneider, A. tRNAs and proteins use the same import channel for translocation across the mitochondrial outer membrane of trypanosomes. Proc. Natl. Acad. Sci. USA 2017, 114, E7679–E7687. [Google Scholar] [CrossRef]
- Pickrell, A.M.; Youle, R.J. The roles of PINK1, parkin, and mitochondrial fidelity in Parkinson’s disease. Neuron 2015, 85, 257–273. [Google Scholar] [CrossRef]
- Hansson Petersen, C.A.; Alikhani, N.; Behbahani, H.; Wiehager, B.; Pavlov, P.F.; Alafuzoff, I.; Leinonen, V.; Ito, A.; Winblad, B.; Glaser, E.; et al. The amyloid beta-peptide is imported into mitochondria via the TOM import machinery and localized to mitochondrial cristae. Proc. Natl. Acad. Sci. USA 2008, 105, 13145–13150. [Google Scholar] [CrossRef]
- Salinas, T.; Duchene, A.M.; Delage, L.; Nilsson, S.; Glaser, E.; Zaepfel, M.; Marechal-Drouard, L. The voltage-dependent anion channel, a major component of the tRNA import machinery in plant mitochondria. Proc. Natl. Acad. Sci. USA 2006, 103, 18362–18367. [Google Scholar] [CrossRef]
- Salinas, T.; El Farouk-Ameqrane, S.; Ubrig, E.; Sauter, C.; Duchene, A.M.; Marechal-Drouard, L. Molecular basis for the differential interaction of plant mitochondrial VDAC proteins with tRNAs. Nucleic Acids Res. 2014, 42, 9937–9948. [Google Scholar] [CrossRef]
- Antos, N.; Budzinska, M.; Kmita, H. An interplay between the TOM complex and porin isoforms in the yeast Saccharomyces cerevisiae mitochondria. FEBS Lett. 2001, 500, 12–16. [Google Scholar] [CrossRef]
- Kmita, H.; Budzińska, M. Involvement of the TOM complex in external NADH transport into yeast mitochondria depleted of mitochondrial porin1. Biochim. Biophys. Acta 2000, 1509, 86–94. [Google Scholar] [CrossRef][Green Version]
- Di Maio, R.; Barrett, P.J.; Hoffman, E.K.; Barrett, C.W.; Zharikov, A.; Borah, A.; Hu, X.; McCoy, J.; Chu, C.T.; Burton, E.A.; et al. α-Synuclein binds to TOM20 and inhibits mitochondrial protein import in Parkinson’s disease. Sci. Transl. Med. 2016, 8, 342ra78. [Google Scholar] [CrossRef] [PubMed]
- Kutik, S.; Stojanovski, D.; Becker, L.; Becker, T.; Meinecke, M.; Kruger, V.; Prinz, C.; Meisinger, C.; Guiard, B.; Wagner, R.; et al. Dissecting membrane insertion of mitochondrial β-barrel proteins. Cell 2008, 132, 1011–1024. [Google Scholar] [CrossRef]
- Chiusolo, V.; Jacquemin, G.; Bassoy, E.Y.; Vinet, L.; Liguori, L.; Walch, M.; Kozjak-Pavlovic, V.; Martinvalet, D. Granzyme B enters the mitochondria in a Sam50-, Tim22- and mtHsp70-dependent manner to induce apoptosis. Cell Death Differ. 2017, 24, 747–758. [Google Scholar] [CrossRef]
- Galganska, H.; Budzinska, M.; Wojtkowska, M.; Kmita, H. Redox regulation of protein expression in Saccharomyces cerevisiae mitochondria: Possible role of VDAC. Arch. Biochem. Biophys. 2008, 479, 39–45. [Google Scholar] [CrossRef]
- Galganska, H.; Karachitos, A.; Wojtkowska, M.; Stobienia, O.; Budzinska, M.; Kmita, H. Communication between mitochondria and nucleus: Putative role for VDAC in reduction/oxidation mechanism. Biochim. Biophys. Acta 2010, 1797, 1276–1280. [Google Scholar] [CrossRef] [PubMed]
- Dejean, L.M.; Ryu, S.Y.; Martinez-Caballero, S.; Teijido, O.; Peixoto, P.M.; Kinnally, K.W. MAC and Bcl-2 family proteins conspire in a deadly plot. Biochim. Biophys. Acta 2010, 1797, 1231–1238. [Google Scholar] [CrossRef] [PubMed]
- Pavlov, E.V.; Priault, M.; Pietkiewicz, D.; Cheng, E.H.Y.; Antonsson, B.; Manon, S.; Korsmeyer, S.J.; Mannella, C.A.; Kinnally, K.W. A novel, high conductance channel of mitochondria linked to apoptosis in mammalian cells and Bax expression in yeast. J. Cell Biol. 2001, 155, 725–732. [Google Scholar] [CrossRef]
- Walther, D.M.; Rapaport, D. Biogenesis of mitochondrial outer membrane proteins. Biochim. Biophys. Acta 2009, 1793, 42–51. [Google Scholar] [CrossRef]
- Sutton, V.R.; Davis, J.E.; Cancilla, M.; Johnstone, R.W.; Ruefli, A.A.; Sedelies, K.; Browne, K.A.; Trapani, J.A. Initiation of apoptosis by granzyme B requires direct cleavage of bid, but not direct granzyme B-mediated caspase activation. J. Exp. Med. 2000, 192, 1403–1414. [Google Scholar] [CrossRef] [PubMed]
- Goping, I.S.; Barry, M.; Liston, P.; Sawchuk, T.; Constantinescu, G.; Michalak, K.M.; Shostak, I.; Roberts, D.L.; Hunter, A.M.; Korneluk, R.; et al. Granzyme B-induced apoptosis requires both direct caspase activation and relief of caspase inhibition. Immunity 2003, 18, 355–365. [Google Scholar] [CrossRef]
- Qiu, J.; Wenz, L.S.; Zerbes, R.M.; Oeljeklaus, S.; Bohnert, M.; Stroud, D.A.; Wirth, C.; Ellenrieder, L.; Thornton, N.; Kutik, S.; et al. Coupling of mitochondrial import and export translocases by receptor-mediated supercomplex formation. Cell 2013, 154, 596–608. [Google Scholar] [CrossRef]
- Wenz, L.S.; Ellenrieder, L.; Qiu, J.; Bohnert, M.; Zufall, N.; van der Laan, M.; Pfanner, N.; Wiedemann, N.; Becker, T. Sam37 is crucial for formation of the mitochondrial TOM–SAM supercomplex, thereby promoting β-barrel biogenesis. J. Cell Biol. 2015, 210, 1047–1054. [Google Scholar] [CrossRef] [PubMed]
- Rampelt, H.; Bohnert, M.; Zerbes, R.M.; Horvath, S.E.; Warscheid, B.; Pfanner, N.; van der Laan, M. Mic10, a core subunit of the mitochondrial contact site and cristae organizing system, interacts with the dimeric F1F0-ATP synthase. J. Mol. Biol. 2017, 429, 1162–1170. [Google Scholar] [CrossRef] [PubMed]
- Yamano, K.; Tanaka-Yamano, S.; Endo, T. Mdm10 as a dynamic constituent of the TOB/SAM complex directs coordinated assembly of Tom40. EMBO Rep. 2010, 11, 187–193. [Google Scholar] [CrossRef]
- Becker, T.; Guiard, B.; Thornton, N.; Zufall, N.; Stroud, D.A.; Wiedemann, N.; Pfanner, N. Assembly of the mitochondrial protein import channel: Role of Tom5 in two-stage interaction of Tom40 with the SAM complex. Mol. Biol. Cell. 2010, 21, 3106–3113. [Google Scholar] [CrossRef]
- Ott, C.; Ross, K.; Straub, S.; Thiede, B.; Gotz, M.; Goosmann, C.; Krischke, M.; Mueller, M.J.; Krohne, G.; Rudel, T.; et al. Sam50 functions in mitochondrial intermembrane space bridging and biogenesis of respiratory complexes. Mol. Cell. Biol. 2012, 32, 1173–1188. [Google Scholar] [CrossRef] [PubMed]
- Ott, C.; Dorsch, E.; Fraunholz, M.; Straub, S.; Kozjak-Pavlovic, V. Detailed analysis of the human mitochondrial contact site complex indicate a hierarchy of subunits. PLoS ONE 2015, 10, e0120213. [Google Scholar] [CrossRef]
- Ding, C.; Wu, Z.; Huang, L.; Wang, Y.; Xue, J.; Chen, S.; Deng, Z.; Wang, L.; Song, Z.; Chen, S. Mitofilin and CHCHD6 physically interact with Sam50 to sustain cristae structure. Sci. Rep. Nov. 2015, 5, 1–11. [Google Scholar] [CrossRef]
- Huynen, M.A.; Mühlmeister, M.; Gotthardt, K.; Guerrero-Castillo, S.; Brandt, U. Evolution and structural organization of the mitochondrial contact site (MICOS) complex and the mitochondrial intermembrane space bridging (MIB) complex. Biochim. Biophys. Acta 2016, 1863, 91–101. [Google Scholar] [CrossRef]
- Tang, J.; Zhang, K.; Dong, J.; Yan, C.; Hu, C.; Ji, H.; Chen, L.; Chen, S.; Zhao, H.; Song, Z. Sam50-Mic19-Mic60 axis determines mitochondrial cristae architecture by mediating mitochondrial outer and inner membrane contact. Cell Death Differ. 2020, 27, 146–160. [Google Scholar] [CrossRef] [PubMed]
- Meisinger, C.; Rissler, M.; Chacinska, A.; Sanjua Szklarz, L.K.; Milenkovic, D.; Kozjak, V.; Schonfisch, B.; Lohaus, C.; Meyer, H.E.; Yaffe, M.P.; et al. The Mitochondrial Morphology Protein Mdm10 Functions in Assembly of the Preprotein Translocase of the Outer Membrane. Dev. Cell 2004, 7, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Thornton, N.; Stroud, D.A.; Milenkovic, D.; Guiard, B.; Pfanner, N.; Becker, T. Two modular forms of the mitochondrial sorting and assembly machinery are involved in biogenesis of a-helical outer membrane proteins. J. Mol. Biol. 2010, 396, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Takeda, H.; Tsutsumi, A.; Nishizawa, T.; Lindau, C.; Busto, J.V.; Wenz, L.S.; Ellenrieder, L.; Imai, K.; Straub, S.P.; Mossmann, W.; et al. Mitochondrial sorting and assembly machinery operates by β-barrel switching. Nature 2021, 590, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Blachly-Dyson, E.; Forte, M. VDAC channels. IUBMB Life 2001, 52, 113–118. [Google Scholar] [PubMed]
- Hohr, A.I.C.; Lindau, C.; Wirth, C.; Qiu, J.; Stroud, D.A.; Kutik, S.; Guiard, B.; Hunte, C.; Becker, T.; Pfanner, N.; et al. Membrane protein insertion through a mitochondrial beta-barrel gate. Science 2018, 359, eaah6834. [Google Scholar] [CrossRef] [PubMed]
- Lionello, S.; Marzaro, G.; Martinvalet, D. SAM50, a side door to the mitochondria: The case of cytotoxic proteases. Pharmacol. Res. 2020, 160, 105196. [Google Scholar] [CrossRef]
- Boldogh, I.R.; Nowakowski, D.W.; Yang, H.C.; Chung, H.; Karmon, S.; Royes, P.; Pon, L.A. A Protein Complex Containing Mdm10p, Mdm12p, and Mmm1p Links Mitochondrial Membranes and DNA to the Cytoskeleton-based Segregation Machinery. Mol. Biol. Cell 2003, 14, 4618–4627. [Google Scholar] [CrossRef]
- Kornmann, K.; Currie, E.; Collins, S.R.; Schuldiner, M.; Nunnari, J.; Weissman, J.S.; Walter, P. An ER-mitochondria tethering complex revealed by a synthetic biology screen. Science 2009, 325, 477–481. [Google Scholar] [CrossRef]
- Flinner, N.; Ellenrieder, L.; Stiller, S.B.; Becker, T.; Schleiff, E.; Mirus, O. Mdm10 is an ancient eukaryotic porin co-occurring with theERMES complex. Biochim. Biophys. Acta 2013, 1833, 3314–3325. [Google Scholar] [CrossRef]
- Meisinger, C.; Wiedemann, N.; Rissler, M.; Strub, A.; Milenkovic, D.; Schonfisch, B.; Muller, H.; Kozjak, V.; Pfanner, N. Mitochondrial Protein Sorting. Differentiation of β-barrel assembly by Tom7-mediated segregation of Mdm10. J. Biol. Chem. 2006, 281, 22819–22826. [Google Scholar] [CrossRef]
- Becker, T.; Pfannschmidt, S.; Guiard, B.; Stojanovski, D.; Milenkovic, D.; Kutik, S.; Pfanner, N.; Meisinger, C.; Wiedemann, N. Biogenesis of the mitochondrial TOM complex. Mim1 promotes insertion and assembly of signal-anchored receptors. J. Biol. Chem. 2008, 283, 120–127. [Google Scholar] [CrossRef]
- Becker, T.; Wenz, L.S.; Kruger, V.; Lehmann, W.; Müller, J.M.; Goroncy, L.; Zufall, N.; Lithgow, T.; Guiard, B.; Chacinska, A.; et al. The mitochondrial import protein Mim1 promotes biogenesis of multispanning outer membrane proteins. J. Cell Biol. 2011, 194, 387–395. [Google Scholar] [CrossRef]
- Papic, D.; Krumpe, K.; Dukanovic, J.; Dimmer, K.S.; Rapaport, D. Multispan mitochondrial outer membrane protein Ugo1 follows a unique Mim1-dependent import pathway. J. Cell Biol. 2011, 194, 397–405. [Google Scholar] [CrossRef]
- Martensson, C.U.; Priesnitz, C.; Song, J.; Ellenrieder, L.; Doan, K.N.; Boos, F.; Floerchinger, A.; Zufall, N.; Oeljeklaus, S.; Warscheid, B.; et al. Mitochondrial protein translocation associated degradation. Nature 2019, 569, 679–683. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, P.M.; Lue, J.K.; Ryu, S.Y.; Wroble, B.N.; Sible, J.C.; Kinnally, K.W. Mitochondrial apoptosis-induced channel (MAC) function triggers a Bax/Bak-dependent bystander effect. Am. J. Pathol. 2011, 178, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Jeandard, D.; Smirnova, A.; Tarassov, I.; Barrey, E.; Smirnov, A.; Entelis, N. Import of non-coding RNAs into human mitochondria: A critical review and emerging approaches. Cells 2019, 8, 286. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, A.; Wallet, C.; Iqbal, R.K.; Gualberto, J.M.; Lotfi, F. Organellar non-coding RNAs: Emerging regulation mechanisms. Biochimie 2015, 117, 48–62. [Google Scholar] [CrossRef]
- Verechshagina, N.; Nikitchina, N.; Yamada, Y.; Harashima, H.; Tanaka, M.; Orishchenko, K.; Mazunin, I. Future of human mitochondrial DNA editing technologies. Mitochondrial DNA Part A DNA Mapp. Seq. Anal. 2019, 30, 214–221. [Google Scholar] [CrossRef]
- Kim, K.M.; Noh, J.H.; Abdelmohsen, K.; Gorospe, M. Mitochondrial noncoding RNA transport. BMB Rep. 2017, 50, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Campo, S.; Gilbert, K.B.; Carrington, J.C. Small RNA-based antiviral defense in the phytopathogenic fungus colletotrichum higginsianum. PLoS Pathog. 2016, 12, e1005640. [Google Scholar] [CrossRef]
- Kamenski, P.; Krasheninnikov, I.; Tarassov, I. 40 Years of Studying RNA Import into Mitochondria: From Basic Mechanisms to Gene Therapy Strategies. Mol. Biol. (Mosk.) 2019, 53, 924–932. [Google Scholar] [CrossRef]
- Ramamonjisoa, D.; Kauffmann, S.; Choisne, N.; Marechal-Drouard, L.; Green, G.; Wintz, H.; Small, I.; Dietrich, A. Structure and expression of several bean (Phaseolus vulgaris) nuclear transfer RNA genes: Relevance to the process of tRNA import into plant mitochondria. Plant Mol. Biol. 1998, 36, 613–625. [Google Scholar] [CrossRef]
- Rusconi, C.P.; Cech, T.R. The anticodon is the signal sequence for mitochondrial import of glutamine tRNA in Tetrahymena. Genes Dev. 1996, 10, 28702880. [Google Scholar] [CrossRef]
- Hancock, K.; Hajduk, S.L. The mitochondrial tRNAs of Trypanosoma brucei are nuclear encoded. J. Biol. Chem. 1990, 265, 19208–19215. [Google Scholar] [CrossRef]
- Shi, X.; Chen, D.H.; Suyama, Y. A nuclear tRNA gene cluster in the protozoan Leishmania tarentolae and differential distribution of nuclear encoded tRNAs between the cytosol and mitochondria. Mol. Biochem. Parasitol. 1994, 65, 2337. [Google Scholar] [CrossRef]
- Kamenski, P.; Smirnova, E.; Kolesnikova, O.; Krasheninnikov, I.A.; Martin, R.P.; Entelis, N.; Tarassov, I. tRNA mitochondrial import in yeast: Mapping of the import determinants in the carrier protein, the precursor of mitochondrial lysyl-tRNA synthetase. Mitochondrion 2010, 10, 284–293. [Google Scholar] [CrossRef]
- Tarassov, I.; Entelis, N.; Martin, R.P. An intact protein translocating machinery is required f or mitochondrial import of a yeast cytoplasmic tRNA. J. Mol. Biol. 1995, 245, 315–323. [Google Scholar] [CrossRef]
- Vyssokikh, M.Y.; Schirtz, T.; Kolesnikova, O.; Entelis, N.; Antonenko, Y.N.; Rokitskaya, T.I.; Tarassov, I. Isoform porin 2 is involved in tRNA(Lys) transport from cytosol to mitochondria in yeast. BBA Bioenerg. 2012, 1817, 124–125. [Google Scholar] [CrossRef][Green Version]
- Harsman, A.; Schneider, A. Mitochondrial protein import in trypanosomes: Expect the unexpected. Traffic 2017, 18, 96–109. [Google Scholar] [CrossRef] [PubMed]
- Shikha, S.; Huot, J.L.; Schneider, A.; Niemann, M. tRNA import across the mitochondrial inner membrane in T. brucei requires TIM subunits but is independent of protein import. Nucleic Acids Res. 2020, 48, 12269–12281. [Google Scholar] [CrossRef] [PubMed]
- Cosma, M.P.; Panizza, S.; Nasmyth, K. Cdk1 Triggers association of RNA polymerase to cell cycle promoters only after recruitment of the mediator by SBF. Mol. Cell 2016, 7, 1213–1220. [Google Scholar] [CrossRef]
- Singha, U.K.; Hamilton, V.; Chaudhuri, M. Tim62, a novel mitochondrial protein in Trypanosoma brucei, is essential for assembly and stability of the TbTim17 protein complex. J. Biol. Chem. 2015, 290, 23226–23239. [Google Scholar] [CrossRef] [PubMed]
- Singha, U.K.; Tripathi, A.; Smith, J.T., Jr.; Quinones, L.; Saha, A.; Singha, T.; Chaudhuri, M. Novel IM-associated protein Tim54 plays a role in the mitochondrial import of internal signal-containing proteins in Trypanosoma brucei. Biol. Cell 2021, 113, 39–57. [Google Scholar] [CrossRef] [PubMed]
- Salinas-Giege, T.; Giege, R.; Giege, P. tRNA biology in mitochondria. Int. J. Mol. Sci. 2015, 16, 4518–4559. [Google Scholar] [CrossRef] [PubMed]
- Bouzaidi-Tiali, N.; Aeby, E.; Charriere, F.; Pusnik, M.; Schneider, A. Elongation factor 1a mediates the specificity of mitochondrial tRNA import in T. brucei. EMBO J. 2007, 26, 4302–4312. [Google Scholar] [CrossRef]
- Entelis, N.; Brandina, I.; Kamenski, P.; Krasheninnikov, I.A.; Martin, R.P.; Tarassov, I. A glycolytic enzyme, enolase, is recruited as a cofactor of tRNA targeting toward mitochondria in Saccharomyces cerevisiae. Genes Dev. 2006, 20, 1609–1620. [Google Scholar] [CrossRef]
- Benz, R. Permeation of hydrophilic solutes through mitochondrial outer membranes: Review on mitochondrial porins. Biochim. Biophys. Acta 1994, 1197, 167–196. [Google Scholar] [CrossRef]
- Colombini, M. VDAC: The channel at the interface between mitochondria and the cytosol. Mol. Cell. Biochem. 2004, 256-257, 107–115. [Google Scholar] [CrossRef]
- Shoshan-Barmatz, V.; De Pinto, V.; Zweckstetter, M.; Raviv, Z.; Keinan, N.; Arbel, N. VDAC, a multi-functional mitochondrial protein regulating cell life and death. Mol. Aspects Med. 2010, 31, 227–285. [Google Scholar] [CrossRef]
- Mannella, C.A. VDAC-A primal perspective. Int. J. Mol. Sci. 2021, 22, 1685. [Google Scholar] [CrossRef]
- De Pinto, V. Renaissance of VDAC: New insights on a protein family at the interface between mitochondria and cytosol biomolecules. Biomolecules 2021, 11, 107. [Google Scholar] [CrossRef]
- Colombini, M. VDAC structure, selectivity, and dynamics. Biochim. Biophys. Acta 2012, 1818, 1457–1465. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Antunes, F.; Canali, R.; Rettori, D.; Cadenas, E. Voltage-dependent anion channels control the release of the superoxide anion from mitochondria to cytosol. J. Biol. Chem. 2003, 278, 5557–5563. [Google Scholar] [CrossRef] [PubMed]
- Homble, F.; Krammer, E.-M.; Prevost, M. Plant VDAC: Facts and speculations. Biochim. Biophys. Acta Biomembr. 2012, 1818, 1486–1501. [Google Scholar] [CrossRef] [PubMed]
- Kmita, H.; Stobienia, O.; Michejda, J. The access of metabolites into yeast mitochondria in the presence and absence of the voltage dependent anion selective channel (YVDAC1). Acta Biochim. Pol. 1999, 46, 991–1000. [Google Scholar] [CrossRef]
- Budzinska, M.; Gałgańska, H.; Karachitos, A.; Wojtkowska, M.; Kmita, H. The TOM complex is involved in the release of superoxide anion from mitochondria. J. Bioenerg. Biomembr. 2009, 41, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Blachly-Dyson, E.; Song, J.; Wolfgang, W.J.; Colombini, M.; Forte, M. Multicopy suppressors of phenotypes resulting from the absence of yeast VDAC encode a VDAC-like protein. Mol. Cell. Biol. 1997, 17, 5727–5738. [Google Scholar] [CrossRef]
- Lee, A.C.; Xu, X.; Blachly-Dyson, E.; Forte, M.; Colombini, M. The role of yeast VDAC genes on the permeability of the mitochondrial outer membrane. J. Membr. Biol. 1998, 161, 173–181. [Google Scholar] [CrossRef]
- Guardiani, C.; Magri, A.; Karachitos, A.; Di Rosa, M.C.; Reina, S.; Bodrenko, I.; Messina, A.; Kmita, H.; Ceccarelli, M.; De Pinto, V. yVDAC2, the second mitochondrial porin isoform of Saccharomyces cerevisiae. Biochim. Biophys. Acta Bioenerg. 2018, 1859, 270–279. [Google Scholar] [CrossRef]
- Magri, A.; Di Rosa, M.C.; Orlandi, I.; Guarino, F.; Reina, S.; Guarnaccia, M.; Morello, G.; Spampinato, A.; Cavallaro, S.; Messina, A.; et al. Deletion of Voltage-Dependent Anion Channel 1 knocks mitochondria down triggering metabolic rewiring in yeast. Cell. Mol. Life Sci. 2020, 77, 3195–3213. [Google Scholar] [CrossRef]
- Kmita, H.; Antos, N.; Wojtkowska, M.; Hryniewiecka, L. Processes underlying the upregulation of Tom proteins in S. cerevisiae mitochondria depleted of the VDAC channel. J. Bioenerg. Biomembr. 2004, 36, 187–193. [Google Scholar] [CrossRef]
- Athenstaedt, K.; Daum, G. 1-Acyldihydroxyacetone-phosphate reductase (Ayr1p) of the yeast Saccharomyces cerevisiae encoded by the open reading frame YIL124w is a major component of lipid particles. J. Biol. Chem. 2000, 275, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Natter, K.; Leitner, P.; Faschinger, A.; Wolinski, H.; McCraith, S.; Fields, S.; Kohlwein, S.D. The spatial organization of lipid synthesis in the yeast Saccharomyces cerevisiae derived from large scale green fluorescent protein tagging and high resolution microscopy. Mol. Cell. Proteom. 2005, 4, 662–672. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Cruz, M.S.; Simmen, T. Over six decades of discovery and characterization of the architecture at mitochondria-associated membranes (MAMs). Adv. Exp. Med. Biol. 2017, 997, 13–31. [Google Scholar] [PubMed]
- Ploier, B.; Scharwey, M.; Koch, B.; Schmidt, C.; Schatte, J.; Rechberger, G.; Kollroser, M.; Hermetter, A.; Daum, G. Screening for hydrolytic enzymes reveals Ayr1p as a novel triacylglycerol lipase in Saccharomyces cerevisiae. J. Biol. Chem. 2013, 288, 36061–36072. [Google Scholar] [CrossRef]


{kind=link}
| Protein Name | Conductance ** | Reversal Potential (mV) *** | PK+/PCl− | Studied Organism | Constitutively Open | Molecules Transported across Channel | Channel Activity Modulators | References |
|---|---|---|---|---|---|---|---|---|
| Tom40 | 370 ± 8 pS a | 40 a | 8 a | S. cerevisiaea | Yes | Mitochondria proteins c RNA d Pink 1 e Aβ f | PC i α-Syn j | [58] a [59] b [6] c [63] d [64] e [65] f [66,67] g [68] h [69] h [58] i [70] j |
| 390 ± 10 pS a | N. crassaa | |||||||
| TOM complex: 740 ± 18 pS (purified complex) a 760 ± 12 pS (complex in OMVs) a | S. cerevisiaea | Yes | TOM complex: RNA g Metabolites h | signal peptides of: CoxIV a preSu9 a,b | ||||
| 625 npS (purified complex) b | A. castellaniib | |||||||
| 575 pS (purified complex) b | D. discoideumb | |||||||
| Tob55/ Sam50 | 640 pS k | 30 k | 4 k | S. cerevisiaek | Yes | β barrel proteins l Granzymes (a and b) m Caspase-3 m Suggested: metabolites n | Unknown | [71] k [30] l [72] m [73,74] n |
| Mdm10 | 480 pS o | 21.5 o | 2.8 o | S. cerevisiaeo | Yes | Unknown | Tom22 o | [14] o |
| Mim1 | 580 pS p | 53 p | 23.5 p | S. cerevisiaep | N/D | α helical outer membrane proteins r | Mim2 p | [4] p [12] r |
| MAC | 1500–5000 pS s | voltage-independent s | 3 t | Mammalian s | No | apoptotic cofactors e.g.,: Cyt c s Smac/DIABLO s AIF s | Bcl-2 s Bcl-xL s Dibucaine s Trifluoperazine s Propranolol (and its derivatives) s | [75] s [76] t |
| OMC7 | 570 pS p | −12.5 p | 0.55 p | S. cerevisiaep | N/D | Suggested: RNA and/or metabolites | Unknown | [4] p |
| OMC8 | 550 pS p | −15.5 p | 0.48 p | S. cerevisiaep | N/D | Suggested: RNA and/or metabolites | Unknown | [4] p |
| Ayr1 | 1470 pS p | 30 p | 4.5 p | S. cerevisiaep | N/D | Suggested: proteins | NADPH r | [4] p |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazur, M.; Kmita, H.; Wojtkowska, M. The Diversity of the Mitochondrial Outer Membrane Protein Import Channels: Emerging Targets for Modulation. Molecules 2021, 26, 4087. https://doi.org/10.3390/molecules26134087
Mazur M, Kmita H, Wojtkowska M. The Diversity of the Mitochondrial Outer Membrane Protein Import Channels: Emerging Targets for Modulation. Molecules. 2021; 26(13):4087. https://doi.org/10.3390/molecules26134087
Chicago/Turabian StyleMazur, Monika, Hanna Kmita, and Małgorzata Wojtkowska. 2021. "The Diversity of the Mitochondrial Outer Membrane Protein Import Channels: Emerging Targets for Modulation" Molecules 26, no. 13: 4087. https://doi.org/10.3390/molecules26134087
APA StyleMazur, M., Kmita, H., & Wojtkowska, M. (2021). The Diversity of the Mitochondrial Outer Membrane Protein Import Channels: Emerging Targets for Modulation. Molecules, 26(13), 4087. https://doi.org/10.3390/molecules26134087