
Diverse Banana Pseudostems and Rachis Are Distinctive for Edible Carbohydrates and Lignocellulose Saccharification towards High Bioethanol Production under Chemical and Liquid Hot Water Pretreatments

 ,
, 
Abstract
:1. Introduction
2. Results and Discussion
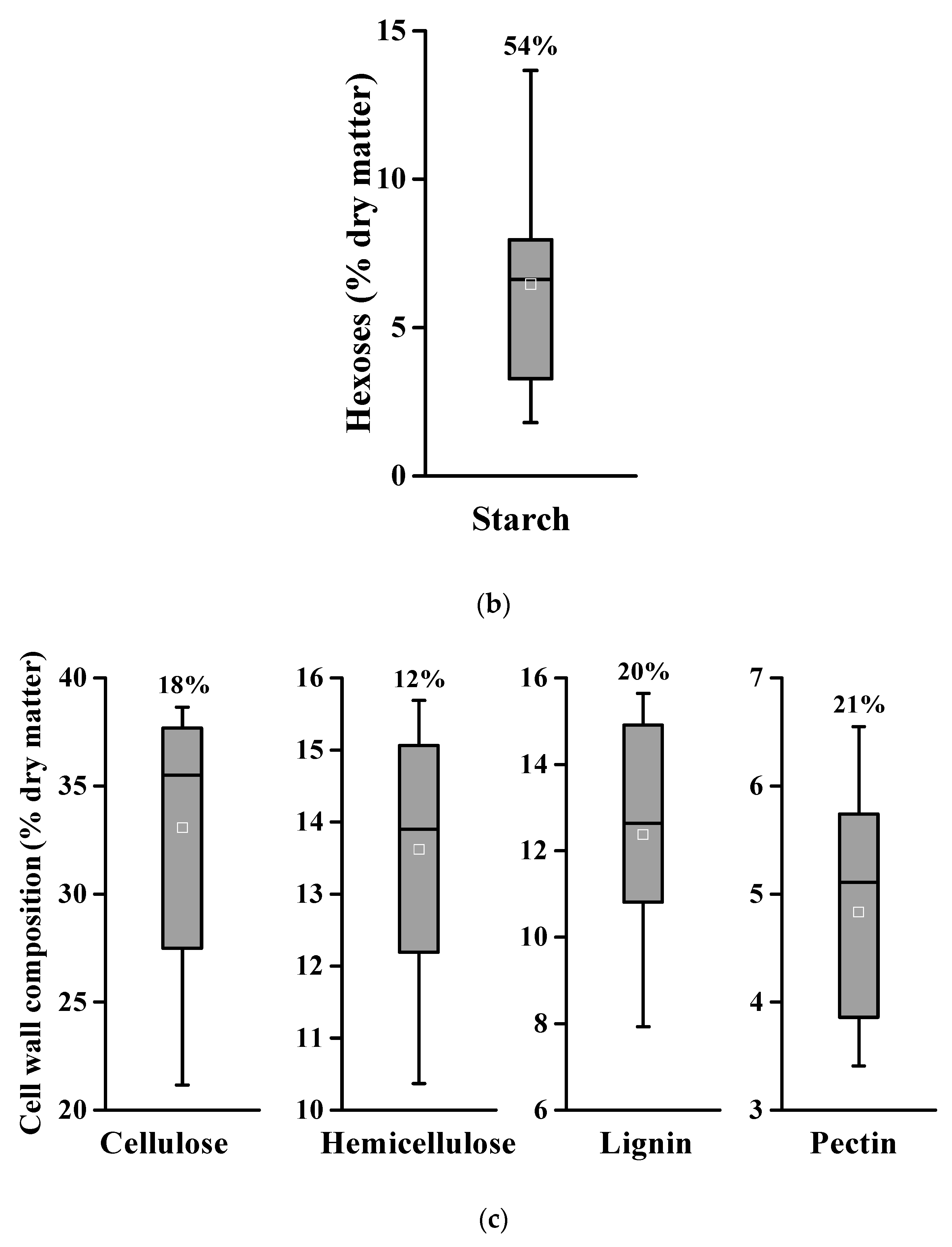
2.1. Large Variations in Edible Carbohydrates and Lignocelluloses among Different Genetic Types of Banana Crops
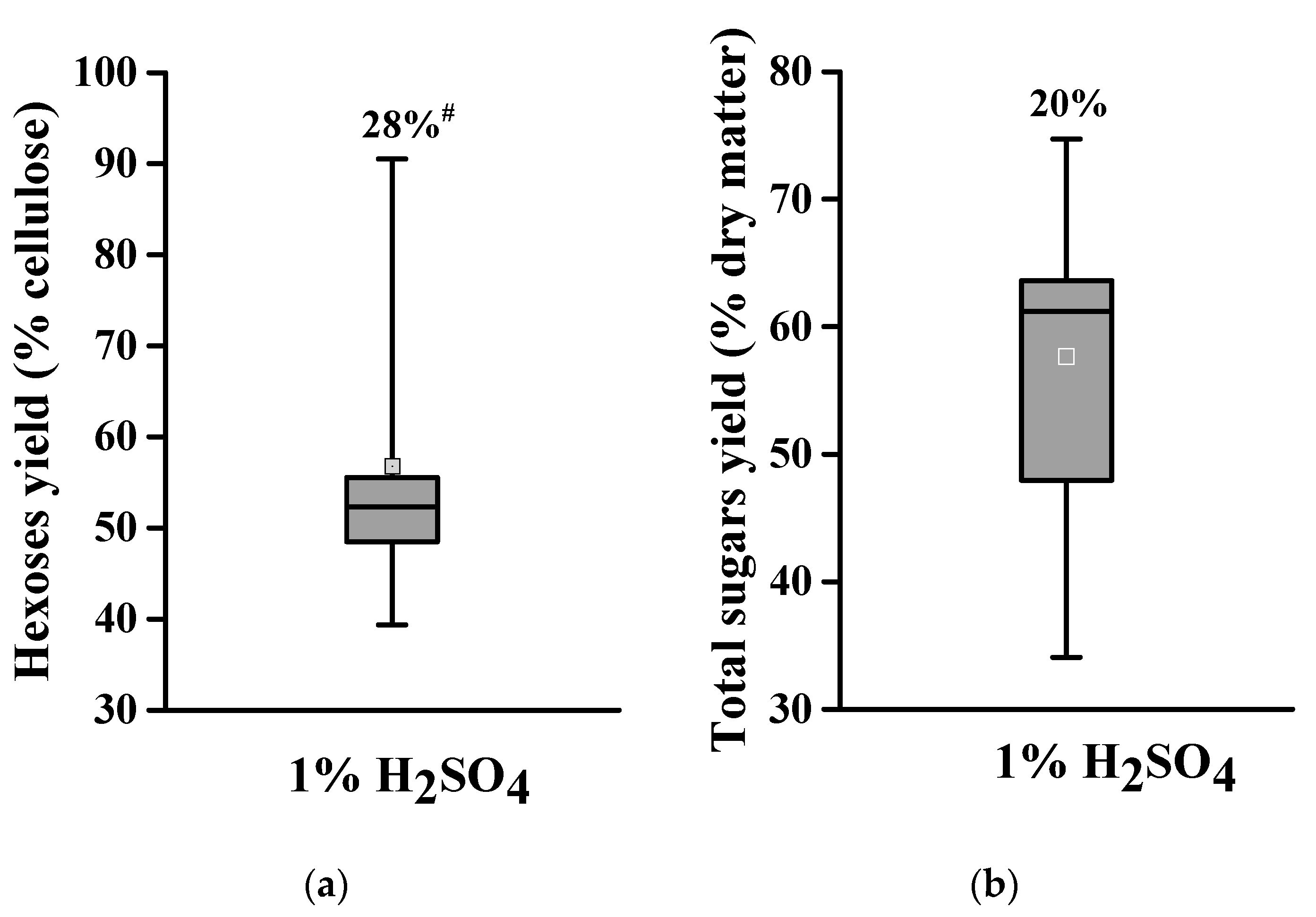
2.2. Acid Pretreatment for Varied Biomass Enzymatic Saccharification among Banana Lignocellulose Samples
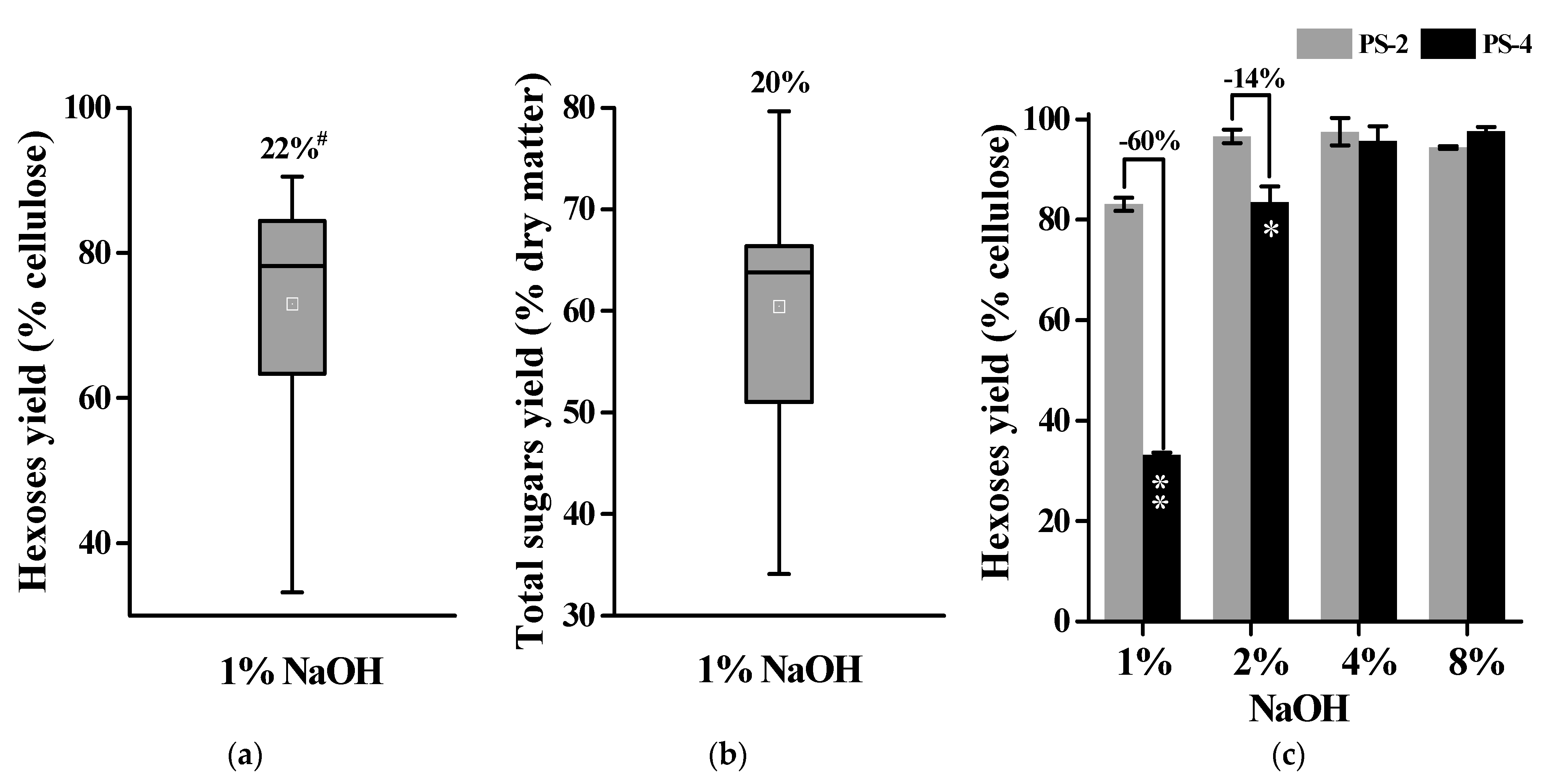
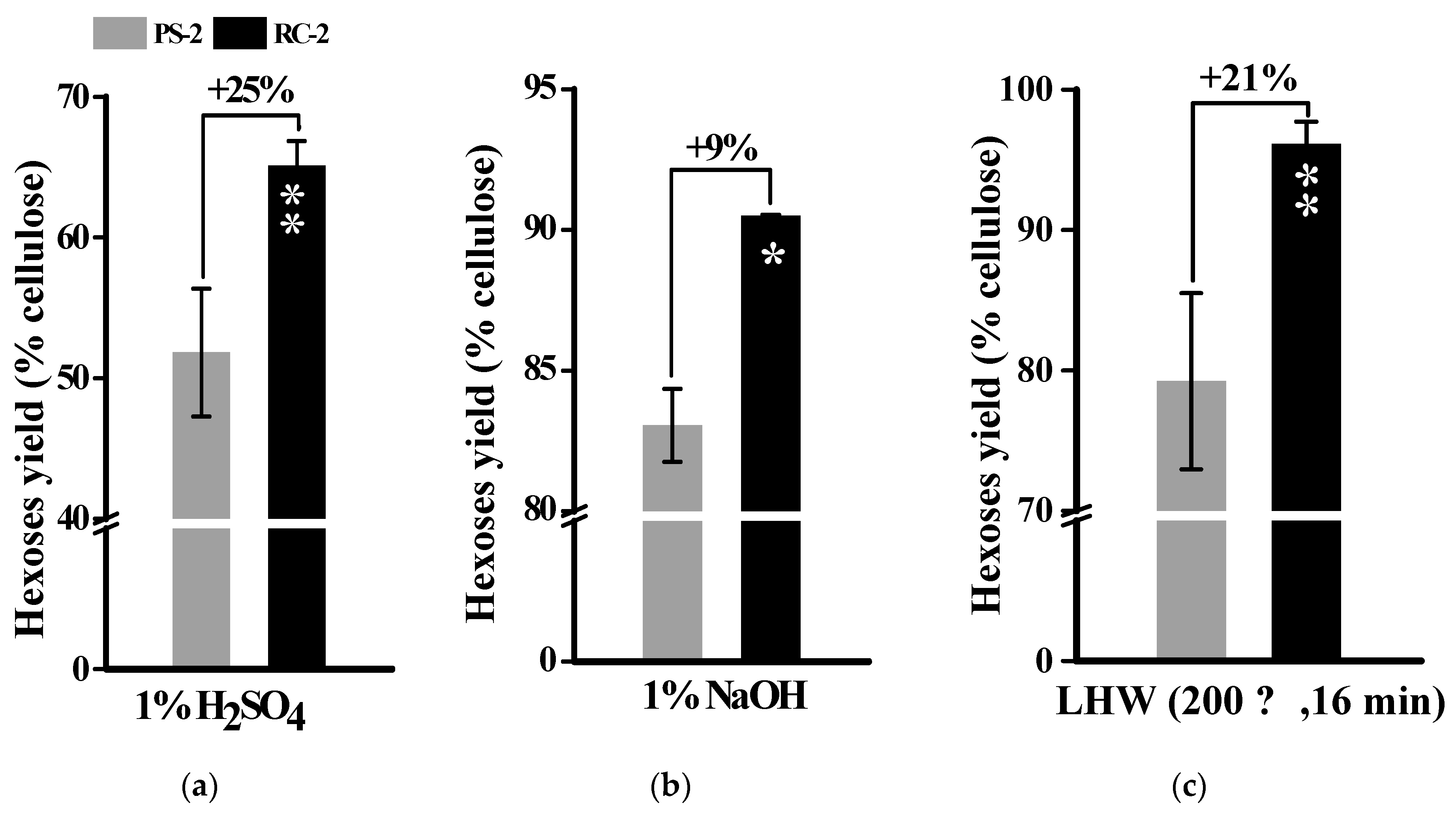
2.3. Alkali Pretreatments for Much Enhanced Biomass Enzymatic Saccharification
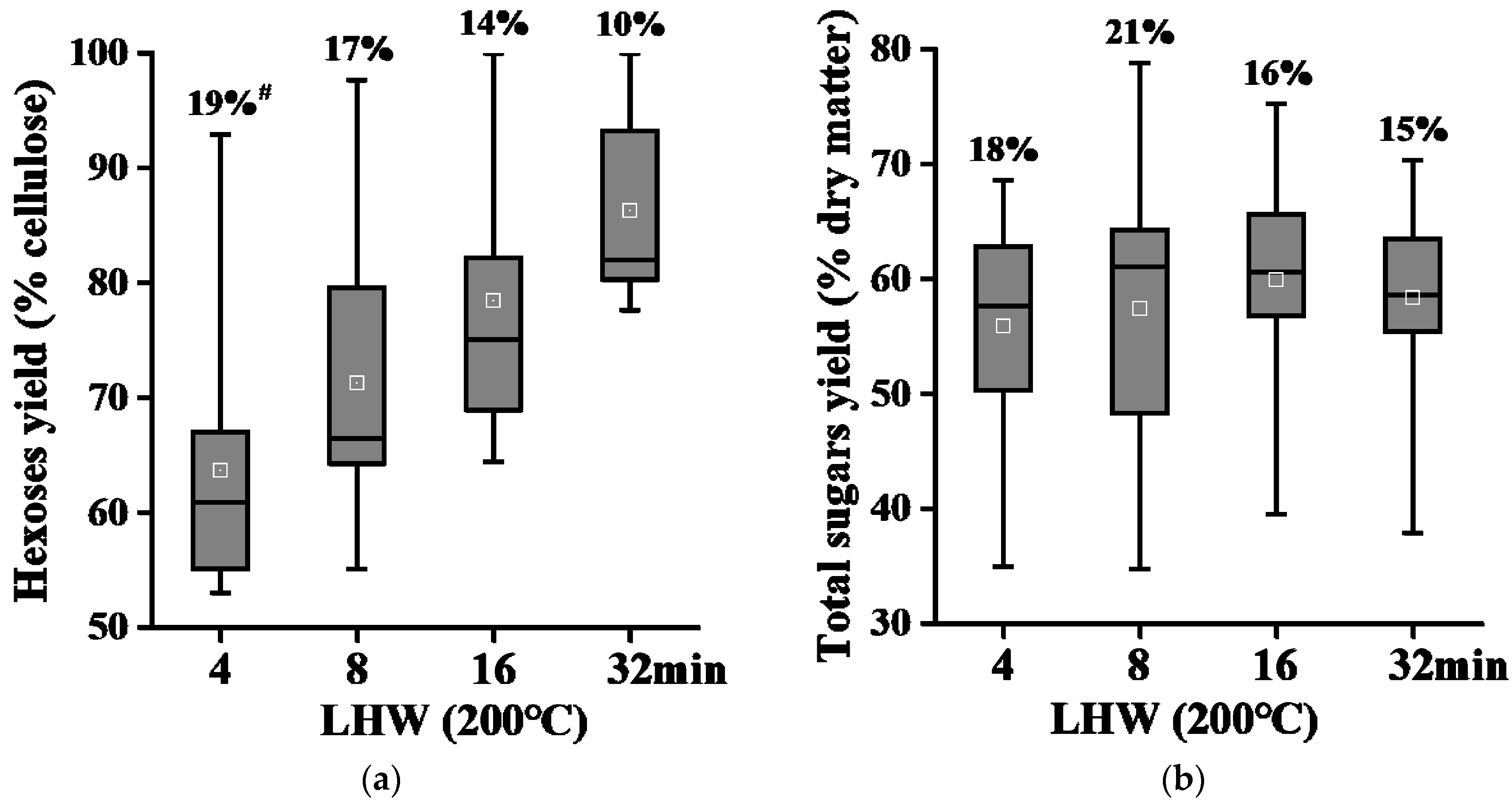
2.4. Liquid Hot Water Pretreatments for Complete Biomass Saccharification in Banana Rachis Samples
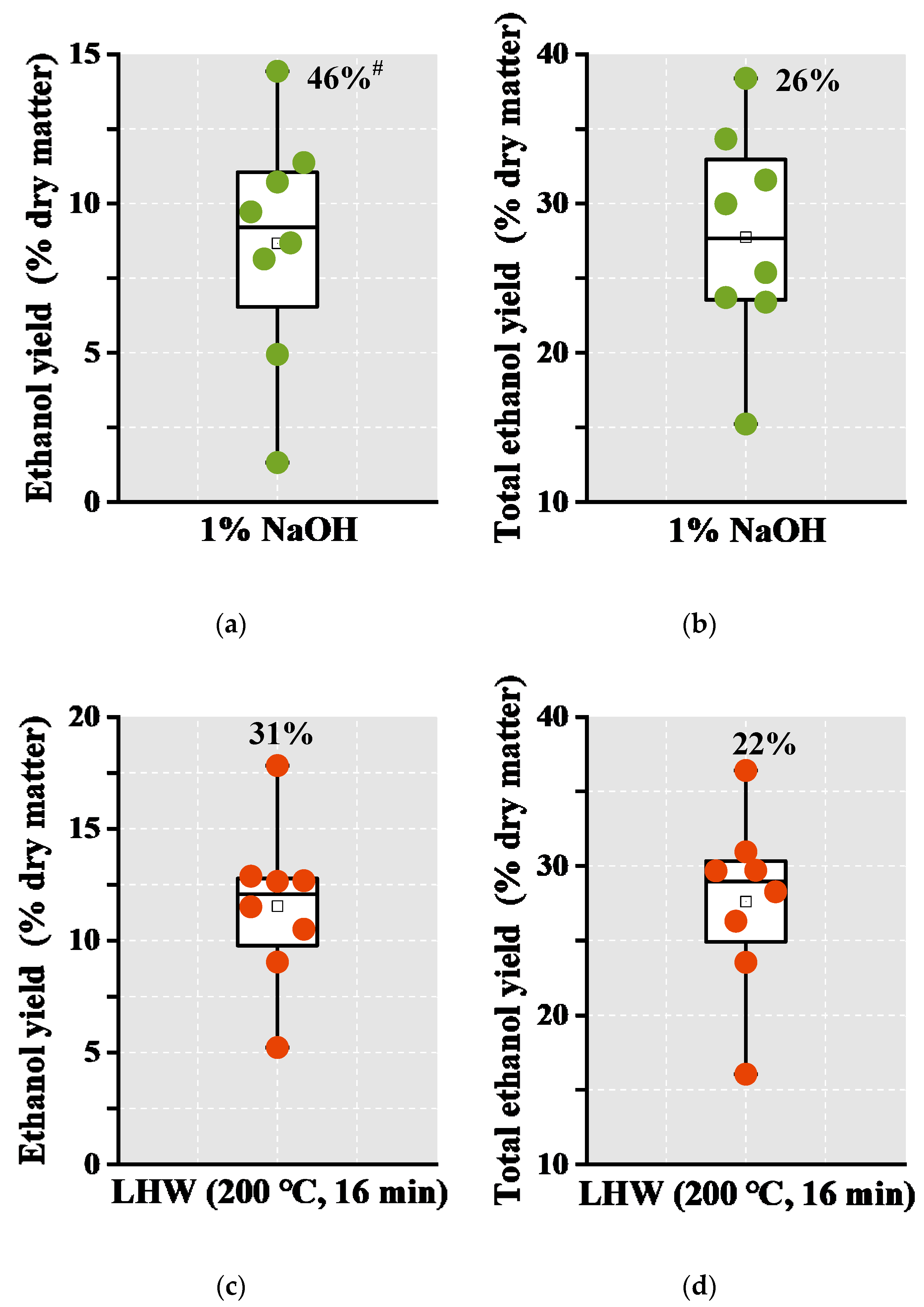
2.5. Remarkably High Bioethanol Production in the Desirable Banana Samples
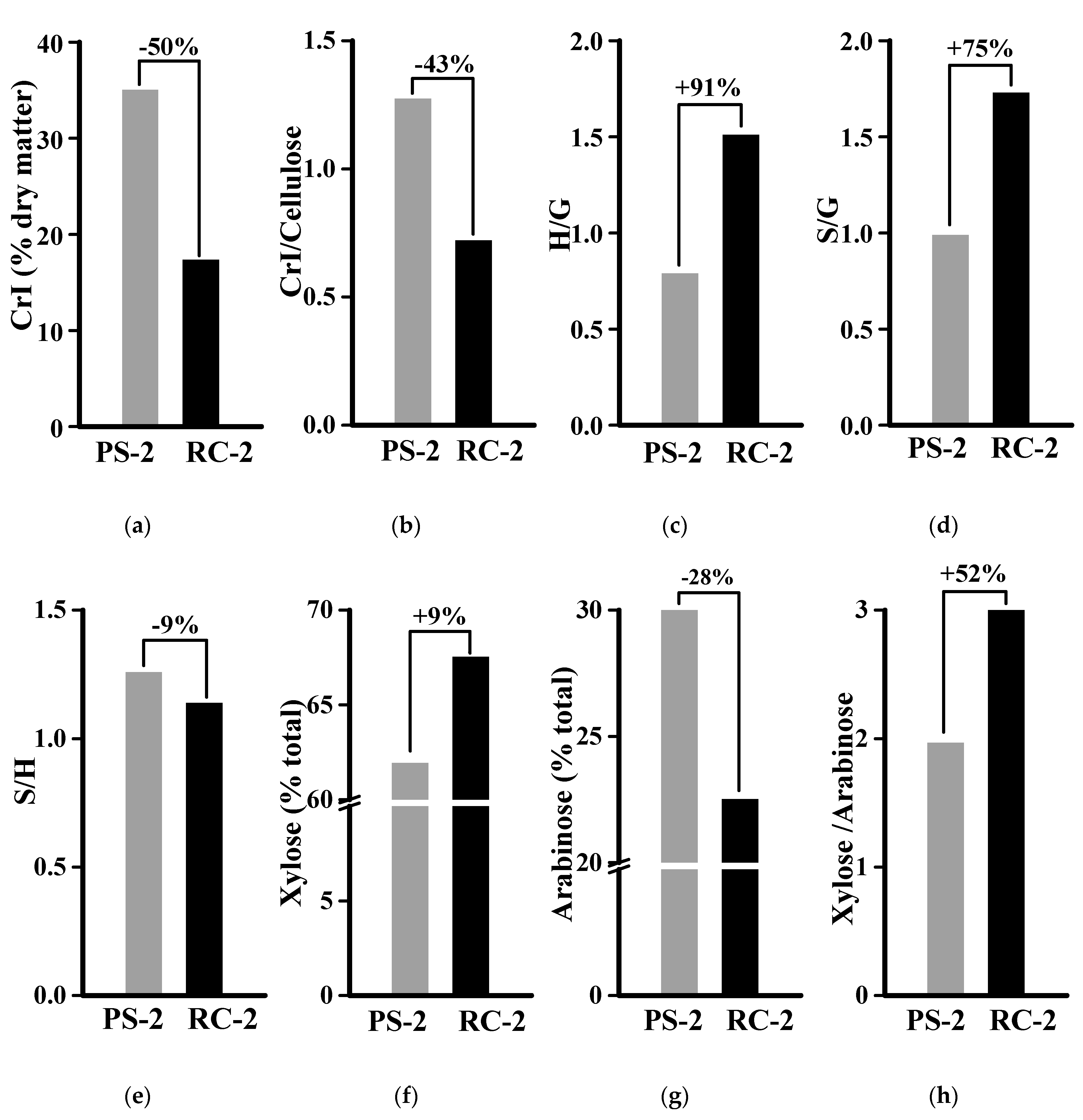
2.6. Characteristic Lignocellulose Features for Distinct Biomass Enzymatic Saccharification in the Desirable Banana Sample
3. Material and Methods
3.1. Banana Samples Collection
3.2. Plant Cell Wall Fractionation
3.3. Colorimetric Assay of Hexoses and Pentoses and Uronic Acids
3.4. Total Lignin and Monolignol Detection
3.5. Hemicellulose Monosaccharide Determination
3.6. Soluble Sugars Extraction and Assay
3.7. Detection of Cellulose Crystalline Index
3.8. Biomass Pretreatments
3.9. Enzymatic Hydrolysis of Pretreated Biomass Residues
3.10. Yeast Fermentation and Ethanol Measurement
3.11. Statistics
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guerrero, A.B.; Ballesteros, I.; Ballesteros, M. The potential of agricultural banana waste for bioethanol production. Fuel 2018, 213, 176–185. [Google Scholar] [CrossRef]
- James, D.; Jean-Yves, P.; Benjamin, D.; Robert, H. Modifying bananas: From transgenics to organics? Sustainability 2017, 9, 333. [Google Scholar]
- Redondo-Gómez, C.; Quesada, M.R.; Astúa, S.; Zamora, J.; Vega-Baudrit, J.R. Biorefinery of biomass of agroindustrial banana waste to obtain high-value biopolymers. Molecules 2020, 25, 3829. [Google Scholar] [CrossRef]
- Antonopoulou, G. Designing Efficient Processes for Sustainable Bioethanol and Bio-Hydrogen Production from Grass Lawn Waste. Molecules 2020, 25, 2889. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, A.B.; Aguado, P.L.; Sánchez, J.; Curt, M.D. GIS-Based Assessment of Banana Residual Biomass Potential for Ethanol Production and Power Generation: A Case Study. Waste Biomass Valorizat. 2016, 7, 405–415. [Google Scholar] [CrossRef] [Green Version]
- Stcker, M. Biofuels and biomass-to-liquid fuels in the biorefinery: Catalytic conversion of lignocellulosic biomass using porous materials. Angew. Chem. Int. Ed. 2008, 47, 9200–9211. [Google Scholar] [CrossRef] [PubMed]
- Scully, M.J.; Norris, G.A.; Falconi, T.; Macintosh, D.L. Carbon intensity of corn ethanol in the united states: State of the science. Environ. Res. Lett. 2021, 16, 4. [Google Scholar] [CrossRef]
- Kotarska, K.; Dziemianowicz, W.; Wierczyńska, A. Study on the sequential combination of bioethanol and biogas production from corn straw. Molecules 2019, 24, 4558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morais, S.; Morag, E.; Barak, Y.; Goldman, D.; Hadar, Y.; Lamed, R.; Shoham, Y.; Wilson, D.B.; Bayer, E.A. Deconstruction of lignocellulose into soluble sugars by native and designer cellulosomes. mBio 2012, 3, e00508-12. [Google Scholar] [CrossRef] [Green Version]
- Passoth, V.; Sandgren, M. Biofuel production from straw hydrolysates: Current achievements and perspectives. Appl. Microbiol. Biotechnol. 2019, 103, 5105–5116. [Google Scholar] [CrossRef] [Green Version]
- Phitsuwan, P.; Sakka, K.; Ratanakhanokchai, K. Improvement of lignocellulosic biomass in planta: A review of feedstocks, biomass recalcitrance, and strategic manipulation of ideal plants designed for ethanol production and processability. Biomass Bioenergy 2013, 58, 390–405. [Google Scholar] [CrossRef]
- Brancourt-Hulmel, M.; Arnoult, S.; Cézard, L.; Hage, F.E.; Reymond, M. A comparative study of maize and Miscanthus re-garding cell-wall composition and stem anatomy for conversion into bioethanol and polymer composites. BioEnerg. Res. 2021. [Google Scholar] [CrossRef]
- Mansfield, S.D.; Meder, R. Cellulose hydrolysis—The role of monocomponent cellulases in crystalline cellulose degradation. Cellulose 2003, 10, 159–169. [Google Scholar] [CrossRef]
- Wang, Y.; Fan, C.; Hu, H.; Li, Y.; Sun, D.; Wang, Y.; Peng, L. Genetic modification of plant cell walls to enhance biomass yield and biofuel production in bioenergy crops. Biotechnol. Adv. 2016, 34, 997–1017. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Zhang, M.; Guo, K.; Hu, Z.; Zhang, R.; Feng, Y.; Yi, X.; Zou, W.; Wang, L.; Wu, C. High-level hemicellulosic arabinose predominately affects lignocellulose crystallinity for genetically enhancing both plant lodging resistance and biomass en-zymatic digestibility in rice mutants. Plant Biotechnol. J. 2014, 13, 514–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, N.; Zhang, W.; Ren, S.; Liu, F.; Zhao, C.; Liao, H.; Xu, Z.; Huang, J.; Li, Q.; Tu, Y. Hemicelluloses negatively affect ligno-cellulose crystallinity for high biomass digestibility under NaOH and H2SO4 pretreatments in Miscanthus. Biotechnol. Biofuels 2012, 5, 58. [Google Scholar] [CrossRef] [Green Version]
- Si, S.; Chen, Y.; Fan, C.; Hu, H.; Li, Y.; Huang, J.; Liao, H.; Hao, B.; Li, Q.; Peng, L.; et al. Lignin extraction distinctively enhances biomass enzymatic saccharification in hemicelluloses-rich Miscanthus species under various alkali and acid pretreatments. Bioresour. Technol. 2015, 183, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, Y.; Takata, N.; Sakamoto, S.; Nakagawa-Izumi, A.; Taniguchi, T.; Ralph, J.; Mitsuda, N.; Kajita, S. Change in lignin structure, but not in lignin content, in transgenic poplar overexpressing the rice master regulator of secondary cell wall biosynthesis. Physiol. Plant. 2018, 163, 170–182. [Google Scholar]
- Xu, C.; Zhu, J.; Yu, H.; Yu, H.; Yang, Y.; Fu, Q.; Zhan, D.; Wang, Y.; Wang, H.; Zhang, Y.; et al. Recyclable cascading of arsenic phytoremediation and lead removal coupled with high bioethanol production using desirable rice straws. Biochem. Eng. J. 2021, 168, 107950. [Google Scholar] [CrossRef]
- Alam, A.; Wang, Y.; Liu, F.; Kang, H.; Tang, S.-w.; Wang, Y.; Cai, Q.; Wang, H.; Peng, H.; Li, Q.; et al. Modeling of optimal green liquor pretreatment for enhanced biomass saccharification and delignification by distinct alteration of wall polymer features and biomass porosity in Miscanthus. Renew. Energ. 2020, 159, 1128–1138. [Google Scholar] [CrossRef]
- Boonsombuti, A.; Trisinsub, O.; Luengnaruemitchai, A. Comparative Study of Three Chemical Pretreatments and Their Effects on the Structural Changes of Rice Straw and Butanol Production. Waste Biomass Valorization 2019, 11, 2771–2781. [Google Scholar] [CrossRef]
- Scopel, E.; Rezende, C.A. Biorefinery on-demand: Modulating pretreatments to recover lignin, hemicellulose, and extractives as co-products during ethanol production. Ind. Crop. Prod. 2021, 163, 113336. [Google Scholar] [CrossRef]
- Mosier, N.; Hendrickson, R.; Ho, N.; Sedlak, M.; Ladisch, M.R. Optimization of pH controlled liquid hot water pretreatment of corn stover. Bioresour. Technol. 2005, 96, 1986–1993. [Google Scholar] [CrossRef]
- Wu, L.; Feng, S.; Deng, J.; Yu, B.; Wang, Y.; He, B.; Peng, H.; Li, Q.; Hu, R.; Peng, L. Altered carbon assimilation and cellulose accessibility to maximize bioethanol yield under low-cost biomass processing in corn brittle stalk. Green Chem. 2019, 21, 4388–4399. [Google Scholar] [CrossRef]
- Clark, J.H.; Luque, R.; Matharu, A.S. Green Chemistry, Biofuels, and Biorefinery. Annu. Rev. Chem. Biomol. Eng. 2012, 3, 183–207. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, P.; Zhang, G.; Yang, Q.; Lu, J.; Xia, T.; Peng, L.; Wang, Y. Cascading of engineered bioenergy plants and fungi sustainable for low-cost bioethanol and high-value biomaterials under green-like biomass processing. Renew. Sustain. Energy Rev. 2021, 137, 110586. [Google Scholar] [CrossRef]
- Li, Y.; Liu, P.; Huang, J.; Zhang, R.; Hu, Z.; Feng, S.; Wang, Y.; Wang, L.; Xia, T.; Peng, L. Mild chemical pretreatments are sufficient for bioethanol production in transgenic rice straws overproducing glucosidase. Green Chem. 2018, 20, 2047–2056. [Google Scholar] [CrossRef]
- Zhang, G.; Wang, L.; Li, X.; Bai, S.; Xue, Y.; Li, Z.; Tang, S.; Wang, Y.; Wang, Y.; Hu, Z. Distinctively altered lignin biosynthe-sis by site-modification of OsCAD2 for enhanced biomass saccharification in rice. GCB Bioenergy 2020, 13, 305–319. [Google Scholar] [CrossRef]
- Cheng, L.; Wang, L.; Wei, L.; Wu, Y.; Xia, T. Combined mild chemical pretreatments for complete cadmium release and cellulosic ethanol co-production distinctive in wheat mutant straw. Green Chem. 2019, 21, 3693–3700. [Google Scholar] [CrossRef]
- Hu, M.; Yu, H.; Li, Y.; Li, A.; Cai, Q.; Liu, P.; Tu, Y.; Wang, Y.; Hu, R.; Hao, B.; et al. Distinct polymer extraction and cellulose DP reduction for complete cellulose hydrolysis under mild chemical pretreatments in sugarcane. Carbohydr. Polym. 2018, 202, 434–443. [Google Scholar] [CrossRef]
- Valinhas, R.V.; Pantoja, L.A.; Maia, A.; Miguel, M.; Santos, A.S. Xylose fermentation to ethanol by new galactomyces geotrichum and candida akabanensis strains. PeerJ 2018, 6, e4673. [Google Scholar] [CrossRef] [Green Version]
- Nadchanok, R.; Pachara, S.; Mamoru, Y. Highly efficient conversion of xylose to ethanol without glucose repression by newly isolated thermotolerant spathaspora passalidarum CMUWF1–2. BMC Microbiol. 2018, 18, 73. [Google Scholar]
- Chen, J.; Wang, X.; Zhang, B.; Yang, Y.; Song, S.; Zhang, F.; Liu, B.; Zhou, Y.; Yi, Y.; Shan, Y.; et al. Integrating enzymatic hydrolysis into subcritical water pretreatment optimization for bioethanol production from wheat straw. Sci. Total Environ. 2021, 770, 145321. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhao, M.; Xu, Q.; Ren, H.; Yin, J. Enhanced enzymatic hydrolysis of sorghum stalk by supercritical carbon dioxide and ultrasonic pretreatment. Appl. Biochem. Biotech. 2019, 188, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Paz-Cedeno, F.R.; Henares, L.R.; Solorzano-Chavez, E.G.; Scontri, M.; Picheli, F.P.; Roldán, I.U.M.; Monti, R.; Oliveira, S.C.; Masarin, F. Evaluation of the effects of different chemical pretreatments in sugarcane bagasse on the response of enzymatic hydrolysis in batch systems subject to high mass loads. Renew. Energ. 2021, 165, 1–13. [Google Scholar] [CrossRef]
- Bay, M.S.; Karimi, K.; Esfahany, M.N.; Kumar, R. Structural modification of pine and poplar wood by alkali pretreatment to improve ethanol production. Ind. Crop. Prod. 2020, 152, 112506. [Google Scholar] [CrossRef]
- Terrett, O.M.; Dupr Ee, P. Covalent interactions between lignin and hemicelluloses in plant secondary cell walls. Curr. Opin. Biotechnol. 2019, 56, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Yoo, C.G.; Meng, X.; Pu, Y.; Ragauskas, A.J. The critical role of lignin in lignocellulosic biomass conversion and recent pretreatment strategies: A comprehensive review. Bioresour. Technol. 2020, 301, 122784. [Google Scholar] [CrossRef]
- dos Santos, A.C.; Ximenes, E.; Kim, Y.; Ladisch, M.R. Lignin–Enzyme Interactions in the Hydrolysis of Lignocellulosic Biomass. Trends Biotechnol. 2019, 37, 518–531. [Google Scholar] [CrossRef]
- Li, M.; Si, S.; Hao, B.; Zha, Y.; Wan, C.; Hong, S.; Kang, Y.; Jia, J.; Zhang, J.; Li, M.; et al. Mild alkali-pretreatment effectively extracts guaiacyl-rich lignin for high lignocellulose digestibility coupled with largely diminishing yeast fermentation inhibitors in Miscanthus. Bioresour. Technol. 2014, 169, 447–454. [Google Scholar] [CrossRef]
- Pei, Y.; Li, Y.; Zhang, Y.; Yu, C.; Fu, T.; Zou, J.; Tu, Y.; Peng, L.; Chen, P. G-lignin and hemicellulosic monosaccharides dis-tinctively affect biomass digestibility in rapeseed. Bioresour. Technol. 2016, 203, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Zhang, G.; Muhammad, A.; Samad, R.A.; Wang, Y.; Walton, J.D.; He, Y.; Peng, L.; Wang, L. Genetic loci simultaneously controlling lignin monomers and biomass digestibility of rice straw. Sci. Rep. 2018, 8, 3636. [Google Scholar] [CrossRef]
- Vangse, C.T.; Srensen, J.F.; Knudsen, K. Aleurone cells are the primary contributor to arabinoxylan oligosaccharide production from wheat bran after treatment with cell wall-degrading enzymes. Int. J. Food Sci. Tech. 2019, 54, 2847–2853. [Google Scholar] [CrossRef]
- Guo, X.; Liu, Y.; Zhang, R.; Luo, J.; Song, Y.; Li, J.; Wu, K.; Peng, L.; Liu, Y.; Du, Y.; et al. Hemicellulose modification promotes cadmium hyperaccumulation by decreasing its retention on roots in Sedum alfredii. Plant Soil 2019, 447, 241–255. [Google Scholar] [CrossRef]
- Peng, L.; Hocart, C.H.; Redmond, J.W.; Williamson, R.E. Fractionation of carbohydrates in Arabidopsis root cell walls shows that three radial swelling loci are specifically involved in cellulose production. Planta 2000, 211, 406–414. [Google Scholar] [CrossRef]
- Huang, Y.; Wei, X.; Zhou, S.; Liu, M.; Tu, Y.; Li, A.; Chen, P.; Wang, Y.; Zhang, X.; Tai, H. Steam explosion distinctively enhances biomass enzymatic saccharification of cotton stalks by largely reducing cellulose polymerization degree in G. Barbadense and G. Hirsutum. Bioresour. Technol. 2015, 181, 224–230. [Google Scholar] [CrossRef]
- Fry, S.C. The Growing Plant Cell Wall: Chemical and Metabolic Analysis, 2nd ed.; Longman: London, UK, 1988; pp. 95–97. [Google Scholar]
- Dische, Z. Color Reactions of Carbohydrates. Methods in Carbohydrate Chemistry, 1st ed.; Whistler, R.L., Wolfrom, M.L., Eds.; Academic Press: New York, NY, USA, 1962; Volume 1, pp. 477–512. [Google Scholar]
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templaton, D.; Crocker, D. Determination of structural carbohydrates and lignin in biomass. Lab. Anal. Proced. 2008, 1617, 1–16. [Google Scholar]
- Fan, C.; Li, Y.; Hu, Z.; Hu, H.; Wang, G.; Li, A.; Wang, Y.; Tu, Y.; Xia, T.; Peng, L. Ectopic expression of a novel osextensin-like gene consistently enhances plant lodging resistance by regulating cell elongation and cell wall thickening in rice. Plant Biotechnol. J. 2018, 16, 254–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, D.; Yang, Q.; Wang, Y.; Gao, H.; He, M.; Lin, X.; Lu, J.; Wang, Y.; Kang, H.; Alam, A.; et al. Distinct mechanisms of enzymatic saccharification and bioethanol conversion enhancement by three surfactants under steam explosion and mild chemical pretreatments in bioenergy Miscanthus. Ind. Crop. Prod. 2020, 153, 112559. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Soluble Sugar | Starch | Cellulose | Hemicellulose | Lignin | Pectin | |
|---|---|---|---|---|---|---|---|
| Hexoses | Pentoses | ||||||
| PS-1 | 26.9 ± 0.6 | 2.4 ± 0.1 | 5.2 ± 0.2 | 35.5 ± 0.7 | 11.9 ± 0.7 | 10.8 ± 0.7 | 5.9 ± 0.1 |
| PS-2 | 32.3 ± 1.5 | 3.2 ± 0.1 | 13.7 ± 0.5 | 27.5 ± 0.5 | 12.2 ± 0.4 | 7.9 ± 0.6 | 5.7 ± 0.1 |
| PS-3 | 9.2 ± 0.7 | 2.5 ± 0.1 | 3.3 ± 0.3 | 38.7 ± 2.1 | 15.1 ± 0.3 | 12.6 ± 0.3 | 5.1 ± 0.2 |
| PS-4 | 21.3 ± 0.4 | 3.4 ± 0.1 | 6.9 ± 0.5 | 37.7 ± 0.9 | 13.9 ± 0.1 | 12.9 ± 0.4 | 3.4 ± 0.1 |
| PS-5 | 22.8 ± 0.2 | 2.9 ± 0.1 | 6.1 ± 0.5 | 37.2 ± 0.5 | 15.2 ± 0.1 | 10.9 ± 0.1 | 4.3 ± 0.1 |
| PS-6 | 8.5 ± 0.5 | 2.3 ± 0.1 | 7.4 ± 0.4 | 36.7 ± 2.4 | 14.2 ± 0.4 | 12.6 ± 0.3 | 3.9 ± 0.1 |
| PS-7 | 17.0 ± 0.5 | 2.6 ± 0.2 | 7.9 ± 0.4 | 32.6 ± 0.8 | 13.7 ± 0.2 | 14.9 ± 0.5 | 6.5 ± 0.4 |
| PS-8 | 16.4 ± 0.3 | 2.3 ± 0.1 | 10.2 ± 0.1 | 35.1 ± 1.9 | 14.7 ± 0.2 | 15.6 ± 0.4 | 5.1 ± 0.2 |
| PS-9 | 14.6 ± 0.4 | 3.8 ± 0.1 | 6.6 ± 0.4 | 37.7 ± 2.1 | 15.7 ± 0.2 | 13.0 ± 0.1 | 5.5 ± 0.2 |
| RC-2 | 42.3 ± 0.2 | 3.6 ± 0.2 | 1.9 ± 0.2 | 24.1 ± 1.7 | 10.4 ± 0.1 | 9.3 ± 0.9 | 3.8 ± 0.1 |
| RC-3 | 3.3 ± 0.3 | 2.7 ± 0.2 | 1.8 ± 0.1 | 21.2 ± 2.2 | 12.8 ± 0.4 | 15.5 ± 0.2 | 3.9 ± 0.2 |
| Material | Pretreatment | Soluble Sugars & Starch | Enzymatic Hydrolysis | Estimated Total Ethanol Yield | Ref. | ||
|---|---|---|---|---|---|---|---|
| Hexoses | Pentoses | Hexoses | Pentoses | ||||
| (% DM) | (% DM) | (% DM) | (% DM) | (% DM) | |||
| Banana (Refen1) Pseudostem (PS-2) | 1% NaOH | 42.4 | 5.3 | 22.8 | 9.2 | 38.4 | This study |
| LHW *, 200 °C, 16 min | 40.7 | 10.2 | 21.7 | 2.5 | 36.4 | ||
| Banana (Refen1)Rachis (RC-2) | 1% NaOH | 36.1 | 4.6 | 21.8 | 8.9 | 34.3 | |
| LHW, 200 °C, 16 min | 33.6 | 8.0 | 23.1 | 11.8 | 30.9 | ||
| Wheat straw | Subcritical water, 220 °C, 22 min | ND | 18.4 | 25.9 | 1.8 | 29.6 | [33] |
| Corn straw | LHW, 200 °C, 20 min | 23.7 | 3.6 | 16.5 | 5.1 | 19.3 | [24] |
| Sweet sorghum stalk | Supercritical carbon dioxide | ND | ND | 43.6 | ND | 22.3 | [34] |
| Sugarcane bagasse | Sulfite-NaOH | 10.2 | 33.3 | 40 | 16.5 | 26.2 | [35] |
| Miscanthus straw | Green liquor, 32%, 150 °C, 32 min | 0.1 | 15.1 | 34.2 | 15.1 | 17.1 | [20] |
| Poplar stem | 8% NaOH | 5.7 | 3.2 | 31.8 | ND | 10.1 | [36] |
| Samples | Cellulose | Hemicellulose | Lignin | Pectin | ||
|---|---|---|---|---|---|---|
| PS-2 | 51.5 ± 0.3 | 22.9 ± 0.3 | 14.9 ± 0.4 | 10.8 ± 0.1 | ||
| RC-2 | 50.7 ± 0.7 | 21.8 ± 0.1 | 19.6 ± 0.2 * | +31% # | 7.9 ± 0.1 * | −27% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Liu, F.; Yu, H.; Li, Y.; Zhou, S.; Ai, Y.; Zhou, X.; Wang, Y.; Wang, L.; Peng, L.; et al. Diverse Banana Pseudostems and Rachis Are Distinctive for Edible Carbohydrates and Lignocellulose Saccharification towards High Bioethanol Production under Chemical and Liquid Hot Water Pretreatments. Molecules 2021, 26, 3870. https://doi.org/10.3390/molecules26133870
Li J, Liu F, Yu H, Li Y, Zhou S, Ai Y, Zhou X, Wang Y, Wang L, Peng L, et al. Diverse Banana Pseudostems and Rachis Are Distinctive for Edible Carbohydrates and Lignocellulose Saccharification towards High Bioethanol Production under Chemical and Liquid Hot Water Pretreatments. Molecules. 2021; 26(13):3870. https://doi.org/10.3390/molecules26133870
Chicago/Turabian StyleLi, Jingyang, Fei Liu, Hua Yu, Yuqi Li, Shiguang Zhou, Yuanhang Ai, Xinyu Zhou, Youmei Wang, Lingqiang Wang, Liangcai Peng, and et al. 2021. "Diverse Banana Pseudostems and Rachis Are Distinctive for Edible Carbohydrates and Lignocellulose Saccharification towards High Bioethanol Production under Chemical and Liquid Hot Water Pretreatments" Molecules 26, no. 13: 3870. https://doi.org/10.3390/molecules26133870
APA StyleLi, J., Liu, F., Yu, H., Li, Y., Zhou, S., Ai, Y., Zhou, X., Wang, Y., Wang, L., Peng, L., & Wang, Y. (2021). Diverse Banana Pseudostems and Rachis Are Distinctive for Edible Carbohydrates and Lignocellulose Saccharification towards High Bioethanol Production under Chemical and Liquid Hot Water Pretreatments. Molecules, 26(13), 3870. https://doi.org/10.3390/molecules26133870