
Changes in Human Erythrocyte Membrane Exposed to Aqueous and Ethanolic Extracts from Uncaria tomentosa


Abstract
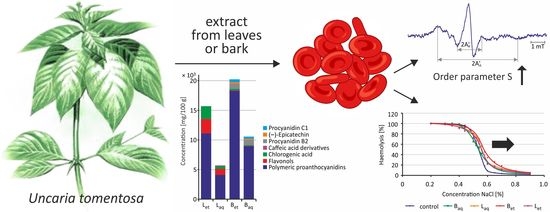
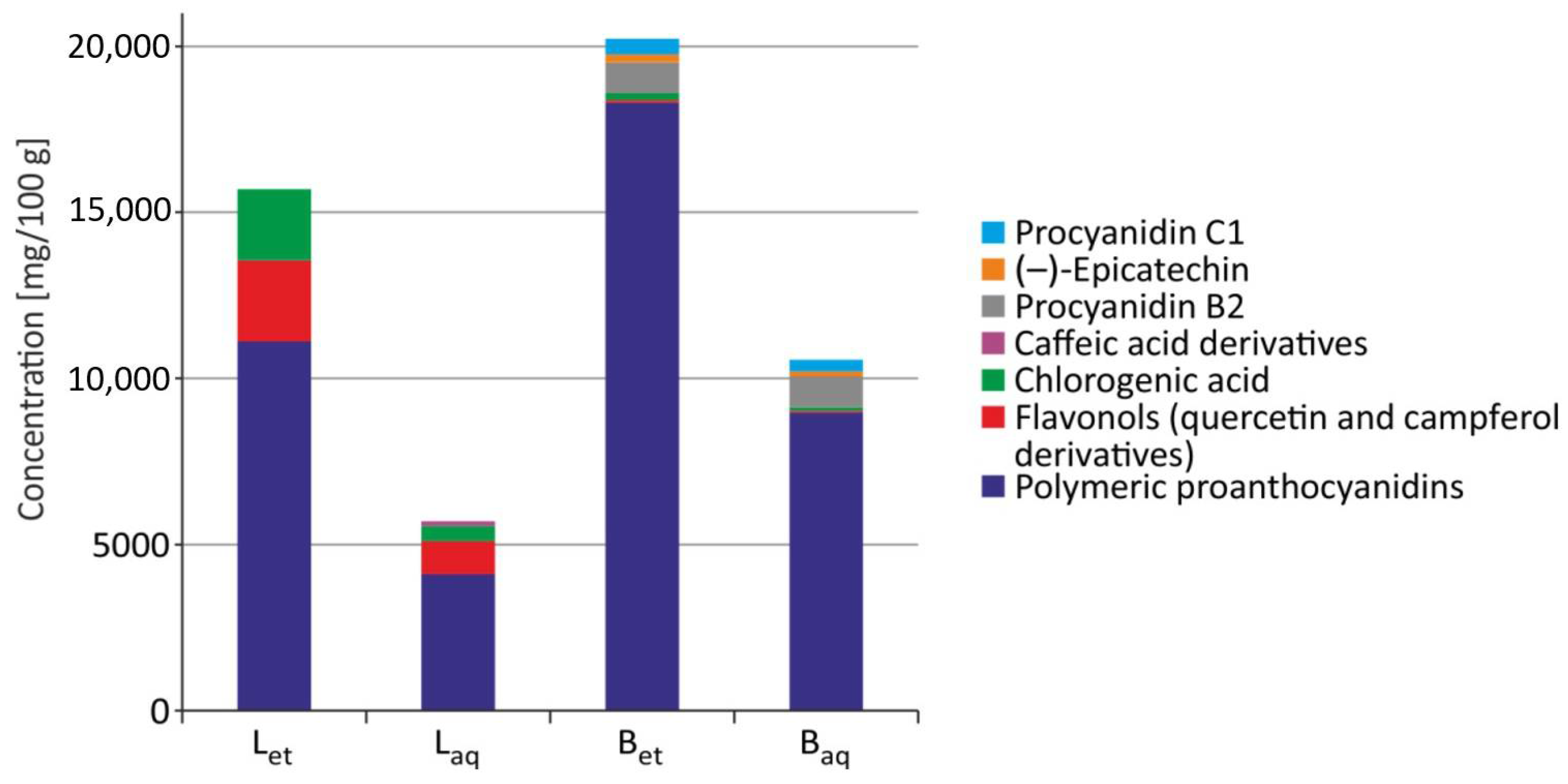
1. Introduction
2. Results
2.1. Membrane Fluidity
2.1.1. Order Parameter S
2.1.2. Correlation Time τB and Time τC
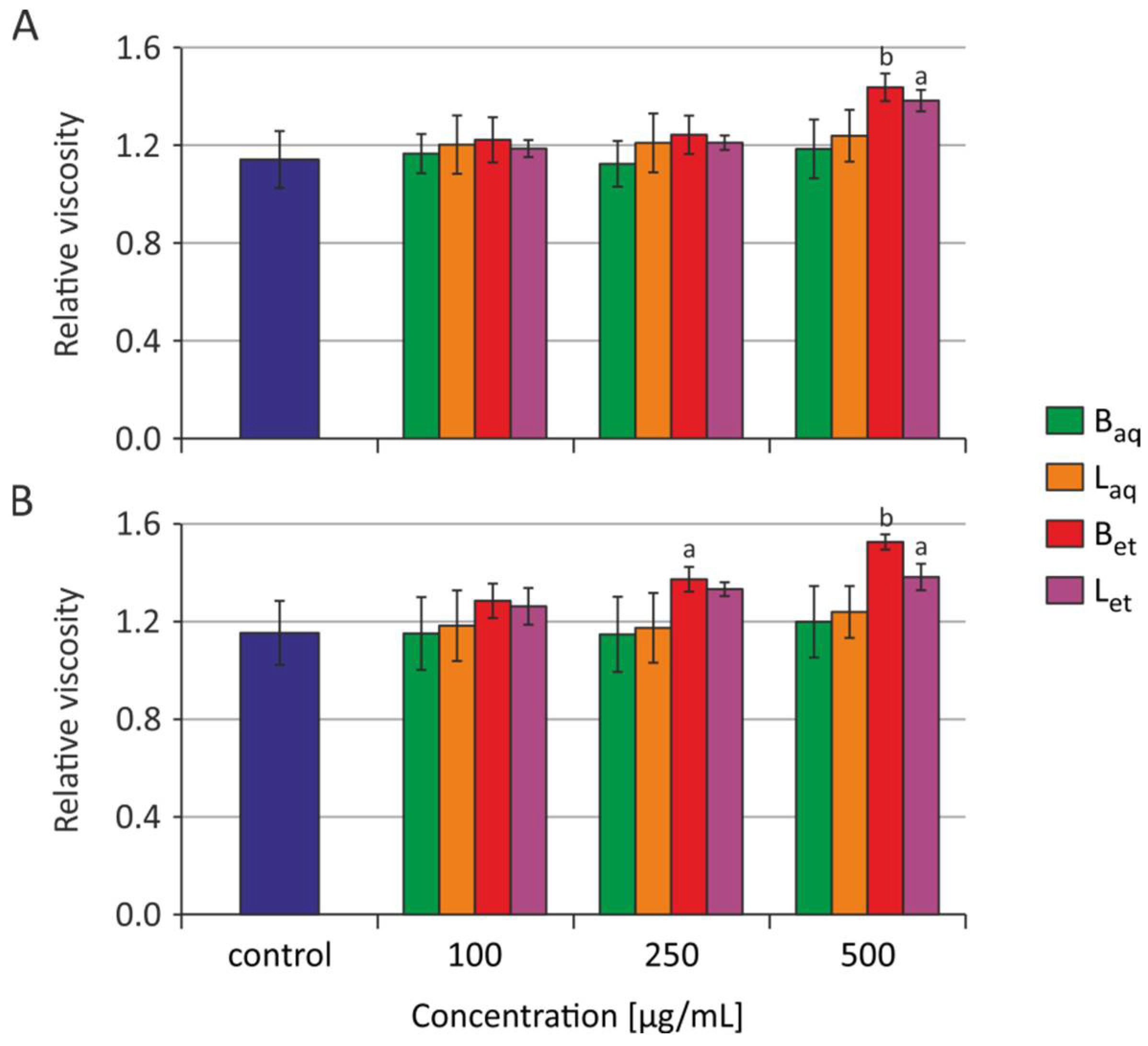
2.2. Internal Viscosity of Red Blood Cells
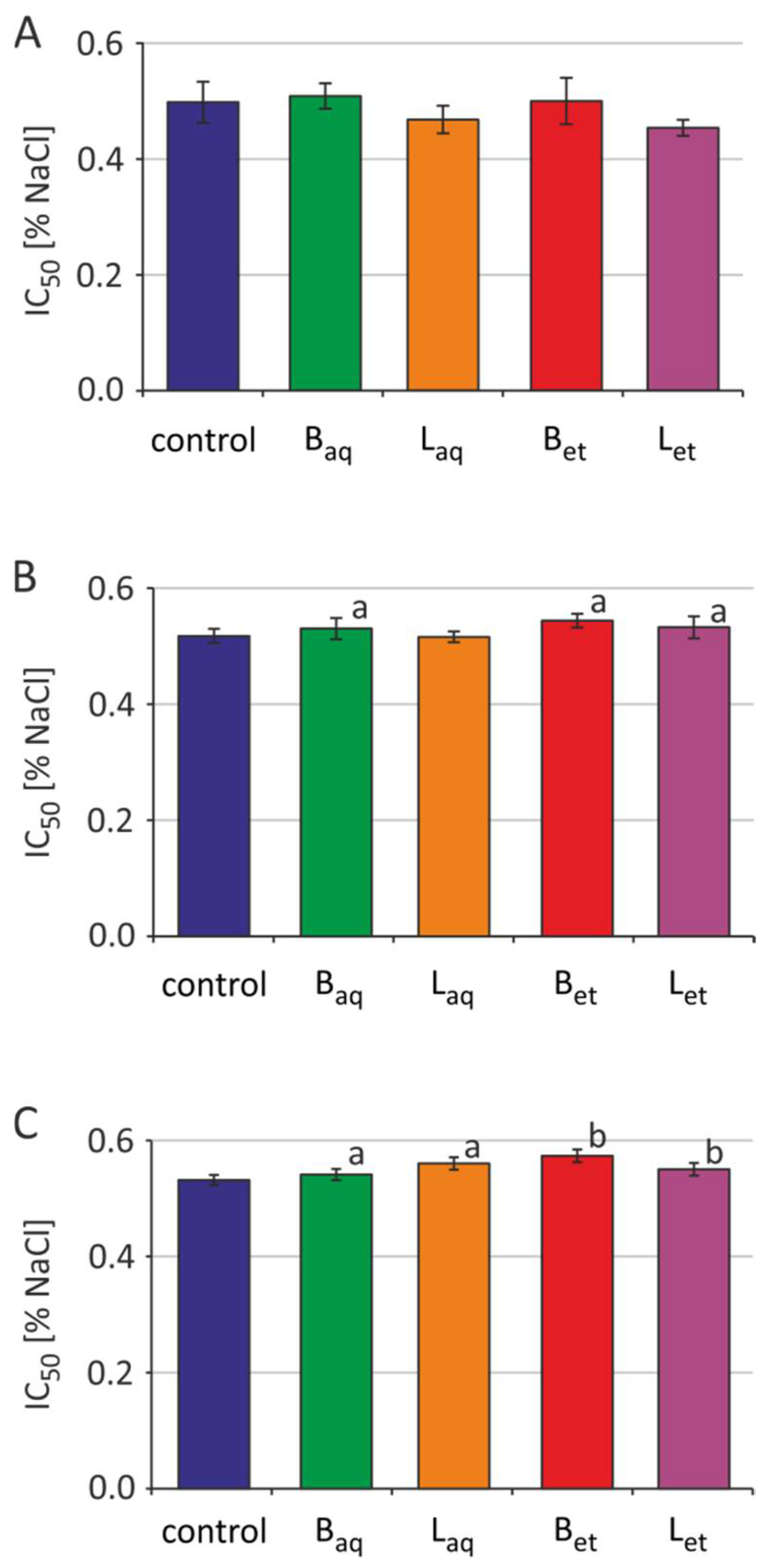
2.3. Osmotic Fragility
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Biological Material
4.2.1. Plant
4.2.2. Isolation of Erythrocytes
4.3. Membrane Fluidity
4.4. Internal Viscosity of Red Blood Cells
4.5. Osmotic Fragility
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- de Azevedo, B.C.; Roxo, M.; Borges, M.C.; Peixoto, H.; Crevelin, E.J.; Bertoni, B.W.; Contini, S.H.; Lopes, A.A.; França, S.C.; Pereira, A.M.; et al. Antioxidant Activity of an Aqueous Leaf Extract from Uncaria tomentosa and Its Major Alkaloids Mitraphylline and Isomitraphylline in Caenorhabditis elegans. Molecules 2019, 24, 3299. [Google Scholar] [CrossRef]
- Navarro, M.; Arnaez, E.; Moreira, I.; Hurtado, A.; Monge, D.; Monagas, M. Polyphenolic Composition and Antioxidant Activity of Uncaria tomentosa Commercial Bark Products. Antioxidants 2019, 8, 339. [Google Scholar] [CrossRef]
- Pilarski, R.; Zieliński, H.; Ciesiołka, D.; Gulewicz, K. Antioxidant activity of ethanolic and aqueous extracts of Uncaria tomentosa (Willd.) DC. J. Ethnopharmacol. 2006, 104, 18–23. [Google Scholar] [CrossRef]
- Santo, G.D.; Grotto, A.; Boligon, A.A.; Da Costa, B.; Rambo, C.L.; Fantini, E.A.; Sauer, E.; Lazzarotto, L.M.V.; Bertoncello, K.T.; Júnior, O.T.; et al. Protective effect of Uncaria tomentosa extract against oxidative stress and genotoxicity induced by glyphosate-Roundup using zebrafish (Danio rerio) as a model. Environ. Sci. Pollut. Res. Int. 2018, 25, 11703–11715. [Google Scholar] [CrossRef]
- Allen-Hall, L.; Arnason, J.T.; Cano, P.; Lafrenie, R.M. Uncaria tomentosa acts as a potent TNF-alpha inhibitor through NF-kappaB. J. Ethnopharmacol. 2010, 127, 685–693. [Google Scholar] [CrossRef]
- Elgawish, R.A.; Abdelrazek, H.M.A.; Ismail, S.A.A.; Loutfy, N.M.; Soliman, M.T.A. Hepatoprotective activity of Uncaria tomentosa extract against sub-chronic exposure to fipronil in male rats. Environ. Sci. Pollut. Res. Int. 2019, 26, 199–207. [Google Scholar] [CrossRef]
- Ciani, F.; Tafuri, S.; Troiano, A.; Cimmino, A.; Fioretto, B.S.; Guarino, A.M.; Pollice, A.; Vivo, M.; Evidente, A.; Carotenuto, D.; et al. Anti-proliferative and pro-apoptotic effects of Uncaria tomentosa aqueous extract in squamous carcinoma cells. J. Ethnopharmacol. 2018, 211, 285–294. [Google Scholar] [CrossRef]
- Zari, A.; Alfarteesh, H.; Buckner, C.; Lafrenie, R.M. Treatment with Uncaria tomentosa Promotes Apoptosis in B16-BL6 Mouse Melanoma Cells and Inhibits the Growth of B16-BL6 Tumours. Molecules 2021, 26, 1066. [Google Scholar] [CrossRef] [PubMed]
- Kośmider, A.; Czepielewska, E.; Kuraś, M.; Gulewicz, K.; Pietrzak, W.; Nowak, R.; Nowicka, G. Uncaria tomentosa Leaves Decoction Modulates Differently ROS Production in Cancer and Normal Cells, and Effects Cisplatin Cytotoxicity. Molecules 2017, 22, 620. [Google Scholar] [CrossRef] [PubMed]
- Almeida, I.V.; Soares, L.C.; Lucio, F.T.; Cantagalli, L.B.; Reusing, A.F.; Vicentini, V.E.P. Chemotherapeutical effects of the herbal medicine Uncaria tomentosa (Willd.) DC. Genet. Mol. Res. 2017, 16. [Google Scholar] [CrossRef] [PubMed]
- Caon, T.; Kaiser, S.; Feltrin, C.; de Carvalho, A.; Sincero, T.C.M.; Ortega, G.G.; Simões, C.M.O. Antimutagenic and antiherpetic activities of different preparations from Uncaria tomentosa (cat’s claw). Food Chem. Toxicol. 2014, 66, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Batiha, G.E.-S.; Magdy Beshbishy, A.; Wasef, L.; Elewa, Y.H.A.; Abd El-Hack, M.E.; Taha, A.E.; Al-Sagheer, A.A.; Devkota, H.P.; Tufarelli, V. Uncaria tomentosa (Willd. ex Schult.) DC.: A Review on Chemical Constituents and Biological Activities. Appl. Sci. 2020, 10, 2668. [Google Scholar] [CrossRef]
- Santos Araújo, M.d.C.; Farias, I.L.; Gutierres, J.; Dalmora, S.L.; Flores, N.; Farias, J.; de Cruz, I.; Chiesa, J.; Morsch, V.M.; Chitolina Schetinger, M.R. Uncaria tomentosa-Adjuvant Treatment for Breast Cancer: Clinical Trial. Evid. Based Complement. Alternat. Med. 2012, 2012, 676984. [Google Scholar] [CrossRef]
- Navarro Hoyos, M.; Sánchez-Patán, F.; Murillo Masis, R.; Martín-Álvarez, P.J.; Zamora Ramirez, W.; Monagas, M.J.; Bartolomé, B. Phenolic Assesment of Uncaria tomentosa L. (Cat’s Claw): Leaves, Stem, Bark and Wood Extracts. Molecules 2015, 20, 22703–22717. [Google Scholar] [CrossRef]
- Farias, I.; do Carmo Araújo, M.; Zimmermann, E.S.; Dalmora, S.L.; Benedetti, A.L.; Alvarez-Silva, M.; Asbahr, A.C.C.; Bertol, G.; Farias, J.; Schetinger, M.R.C. Uncaria tomentosa stimulates the proliferation of myeloid progenitor cells. J. Ethnopharmacol. 2011, 137, 856–863. [Google Scholar] [CrossRef] [PubMed]
- Yunis-Aguinaga, J.; Claudiano, G.S.; Marcusso, P.F.; Manrique, W.G.; de Moraes, J.R.E.; de Moraes, F.R.; Fernandes, J.B.K. Uncaria tomentosa increases growth and immune activity in Oreochromis niloticus challenged with Streptococcus agalactiae. Fish. Shellfish Immunol. 2015, 47, 630–638. [Google Scholar] [CrossRef] [PubMed]
- Bors, M.; Bukowska, B.; Pilarski, R.; Gulewicz, K.; Oszmiański, J.; Michałowicz, J.; Koter-Michalak, M. Protective activity of the Uncaria tomentosa extracts on human erythrocytes in oxidative stress induced by 2,4-dichlorophenol (2,4-DCP) and catechol. Food Chem. Toxicol. 2011, 49, 2202–2211. [Google Scholar] [CrossRef]
- Bors, M.; Michałowicz, J.; Pilarski, R.; Sicińska, P.; Gulewicz, K.; Bukowska, B. Studies of biological properties of Uncaria tomentosa extracts on human blood mononuclear cells. J. Ethnopharmacol. 2012, 142, 669–678. [Google Scholar] [CrossRef]
- Bors, M.; Sicińska, P.; Michałowicz, J.; Wieteska, P.; Gulewicz, K.; Bukowska, B. Evaluation of the effect of Uncaria tomentosa extracts on the size and shape of human erythrocytes (in vitro). Environ. Toxicol. Pharmacol. 2012, 33, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Y.; Li, L.; Holmgren, K.; Pero, R.W. DNA repair enhancement of aqueous extracts of Uncaria tomentosa in a human volunteer study. Phytomedicine 2001, 8, 275–282. [Google Scholar] [CrossRef]
- Piscoya, J.; Rodriguez, Z.; Bustamante, S.A.; Okuhama, N.N.; Miller, M.J.; Sandoval, M. Efficacy and safety of freeze-dried cat’s claw in osteoarthritis of the knee: Mechanisms of action of the species Uncaria guianensis. Inflamm. Res. 2001, 50, 442–448. [Google Scholar] [CrossRef]
- Bukowska, B.; Bors, M.; Gulewicz, K.; Koter-Michalak, M. Uncaria tomentosa extracts protect human erythrocyte catalase against damage induced by 2,4-D-Na and its metabolites. Food Chem. Toxicol. 2012, 50, 2123–2127. [Google Scholar] [CrossRef]
- Tena, N.; Martín, J.; Asuero, A.G. State of the Art of Anthocyanins: Antioxidant Activity, Sources, Bioavailability, and Therapeutic Effect in Human Health. Antioxidants 2020, 9, 451. [Google Scholar] [CrossRef]
- Murkovic, M.; Adam, U.; Pfannhauser, W. Analysis of anthocyane glycosides in human serum. Fresenius. J. Anal. Chem. 2000, 366, 379–381. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, I.; Faria, A.; Calhau, C.; de Freitas, V.; Mateus, N. Bioavailability of anthocyanins and derivatives. J. Funct. Foods 2014, 7, 54–66. [Google Scholar] [CrossRef]
- Harris, R.B.; Foote, J.A.; Hakim, I.A.; Bronson, D.L.; Alberts, D.S. Fatty acid composition of red blood cell membranes and risk of squamous cell carcinoma of the skin. Cancer Epidemiol. Biomark. Prev. 2005, 14, 906–912. [Google Scholar] [CrossRef]
- Jewell, S.A.; Petrov, P.G.; Winlove, C.P. The effect of oxidative stress on the membrane dipole potential of human red blood cells. Biochim. Biophys. Acta 2013, 1828, 1250–1258. [Google Scholar] [CrossRef]
- Li, J.; Lykotrafitis, G.; Dao, M.; Suresh, S. Cytoskeletal dynamics of human erythrocyte. Proc. Natl. Acad. Sci. USA 2007, 104, 4937–4942. [Google Scholar] [CrossRef] [PubMed]
- Jarosiewicz, M.; Michałowicz, J.; Bukowska, B. In vitro assessment of eryptotic potential of tetrabromobisphenol A and other bromophenolic flame retardants. Chemosphere 2019, 215, 404–412. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Tateishi, N.; Cicha, I.; Maeda, N. Aggregation and sedimentation of mixtures of erythrocytes with different properties. Clin. Hemorheol. Microcirc. 2001, 25, 105–117. [Google Scholar] [PubMed]
- Sandoval, M.; Okuhama, N.N.; Zhang, X.; Condezo, L.A.; Lao, J.; Angeles, F.M.; Musah, R.A.; Bobrowski, P.; Miller, M.J. Anti-inflammatory and antioxidant activities of cat’s claw (Uncaria tomentosa and Uncaria guianensis) are independent of their alkaloid content. Phytomedicine 2002, 9, 325–337. [Google Scholar] [CrossRef] [PubMed]
- Maćczak, A.; Duchnowicz, P.; Sicińska, P.; Koter-Michalak, M.; Bukowska, B.; Michałowicz, J. The in vitro comparative study of the effect of BPA, BPS, BPF and BPAF on human erythrocyte membrane; perturbations in membrane fluidity, alterations in conformational state and damage to proteins, changes in ATP level and Na+/K+ ATPase and AChE activities. Food Chem. Toxicol. 2017, 110, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Duchnowicz, P.; Koter, M. Damage to the erythrocyte membrane caused by chlorophenoxyacetic herbicides. Cell. Mol. Biol. Lett. 2003, 8, 25–30. [Google Scholar] [PubMed]
- Boesze-Battaglia, K.; Schimmel, R.J. Cell membrane lipid composition and distribution: Implications for cell function and lessons learned from photoreceptors and platelets. J. Exp. Biol. 1997, 200, 2927–2936. [Google Scholar] [CrossRef] [PubMed]
- Bukowska, B.; Marczak, A.; Michałowicz, J.; Wiśniewska, K. Effects of Phenol, Catechol, Chloro- and Metylphenol on Human Erythrocyte Membrane (in vitro). Pol. J. Environ. Stud. 2009, 18, 569–577. [Google Scholar]
- Kuhn, V.; Diederich, L.; Keller, T.C.S.; Kramer, C.M.; Lückstädt, W.; Panknin, C.; Suvorava, T.; Isakson, B.E.; Kelm, M.; Cortese-Krott, M.M. Red Blood Cell Function and Dysfunction: Redox Regulation, Nitric Oxide Metabolism, Anemia. Antioxid. Redox Signal. 2017, 26, 718–742. [Google Scholar] [CrossRef]
- Koren, E.; Kohen, R.; Ginsburg, I. Polyphenols enhance total oxidant-scavenging capacities of human blood by binding to red blood cells. Exp. Biol. Med. 2010, 235, 689–699. [Google Scholar] [CrossRef]
- Stasiuk, M.; Kijanka, G.M.; Kozubek, A. Transformations of erythrocytes shape and its regulation. Postepy Biochem. 2009, 55, 425–433. [Google Scholar]
- Sheetz, M.P.; Singer, S.J. Biological membranes as bilayer couples. A molecular mechanism of drug-erythrocyte interactions. Proc. Natl. Acad. Sci. USA 1974, 71, 4457–4461. [Google Scholar] [CrossRef]
- Duchnowicz, P.; Broncel, M.; Podsędek, A.; Koter-Michalak, M. Hypolipidemic and antioxidant effects of hydroxycinnamic acids, quercetin, and cyanidin 3-glucoside in hypercholesterolemic erythrocytes (in vitro study). Eur. J. Nutr. 2012, 51, 435–443. [Google Scholar] [CrossRef]
- Ruskovska, T.; Maksimova, V.; Milenkovic, D. Polyphenols in human nutrition: From the in vitro antioxidant capacity to the beneficial effects on cardiometabolic health and related inter-individual variability—An overview and perspective. Br. J. Nutr. 2020, 123, 241–254. [Google Scholar] [CrossRef]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The Role of Polyphenols in Human Health and Food Systems: A Mini-Review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef]
- Heitzman, M.E.; Neto, C.C.; Winiarz, E.; Vaisberg, A.J.; Hammond, G.B. Ethnobotany, phytochemistry and pharmacology of Uncaria (Rubiaceae). Phytochemistry 2005, 66, 5–29. [Google Scholar] [CrossRef] [PubMed]
- Tarahovsky, Y.S.; Kim, Y.A.; Yagolnik, E.A.; Muzafarov, E.N. Flavonoid-membrane interactions: Involvement of flavonoid-metal complexes in raft signaling. Biochim. Biophys. Acta 2014, 1838, 1235–1246. [Google Scholar] [CrossRef]
- Słoczyńska, K.; Kózka, M.; Marona, H. Red blood cell deformability and aggregation in chronic venous disease patients with varicose veins. Postepy Hig. Med. Dosw. 2013, 67, 690–694. [Google Scholar] [CrossRef]
- Naparlo, K.; Bartosz, G.; Stefaniuk, I.; Cieniek, B.; Soszynski, M.; Sadowska-Bartosz, I. Interaction of Catechins with Human Erythrocytes. Molecules 2020, 25, 1456. [Google Scholar] [CrossRef]
- Shahidi, F.; Yeo, J. Bioactivities of Phenolics by Focusing on Suppression of Chronic Diseases: A Review. Int. J. Mol. Sci. 2018, 19, 1573. [Google Scholar] [CrossRef]
- Sekiya, N.; Shimada, Y.; Shibahara, N.; Takagi, S.; Yokoyama, K.; Kasahara, Y.; Sakakibara, I.; Terasawa, K. Inhibitory effects of Choto-san (Diao-teng-san), and hooks and stems of Uncaria sinensis on free radical-induced lysis of rat red blood cells. Phytomedicine 2002, 9, 636–640. [Google Scholar] [CrossRef]
- Kim, S.; Elgegren, M.; Donayre, A.; Galaretta, B.; Nakamatsu, J. Bioactive Membranes of Polymeric Micro and Nanocomposites Prepared with the Natural Anionic Marine Polysaccharide (Alginate) Functionalized with Extracts of Cat’s Claw (Uncaria tomentosa) and Aloe vera. MSF 2020, 1007, 131–136. [Google Scholar] [CrossRef]
- Valenzuela, R.; Echeverria, F.; Ortiz, M.; Rincón-Cervera, M.Á.; Espinosa, A.; Hernandez-Rodas, M.C.; Illesca, P.; Valenzuela, A.; Videla, L.A. Hydroxytyrosol prevents reduction in liver activity of Δ-5 and Δ-6 desaturases, oxidative stress, and depletion in long chain polyunsaturated fatty acid content in different tissues of high-fat diet fed mice. Lipids Health Dis. 2017, 16, 64. [Google Scholar] [CrossRef] [PubMed]
- Rincón-Cervera, M.A.; Valenzuela, R.; Hernandez-Rodas, M.C.; Marambio, M.; Espinosa, A.; Mayer, S.; Romero, N.; Barrera, M.; Sc, C.; Valenzuela, A.; et al. Supplementation with antioxidant-rich extra virgin olive oil prevents hepatic oxidative stress and reduction of desaturation capacity in mice fed a high-fat diet: Effects on fatty acid composition in liver and extrahepatic tissues. Nutrition 2016, 32, 1254–1267. [Google Scholar] [CrossRef] [PubMed]
- Koter, M.; Franiak, I.; Strychalska, K.; Broncel, M.; Chojnowska-Jezierska, J. Damage to the structure of erythrocyte plasma membranes in patients with type-2 hypercholesterolemia. Int. J. Biochem. Cell Biol. 2004, 36, 205–215. [Google Scholar] [CrossRef]
- Morse, P.D. Use of the spin label tempamine for measuring the internal viscosity of red blood cells. Biochem. Biophys. Res. Commun. 1977, 77, 1486–1491. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration | τB (×10−10 s) | τC (×10−10 s) | |||
|---|---|---|---|---|---|
| µmol/L | 2 h | 24 h | 2 h | 24 h | |
| control | 17.636 ± 0.314 | 17.557 ± 0.307 | 23.867 ± 0.468 | 23.757 ± 0.677 | |
| 100 | 17.903 ± 0.308 | 17.787 ± 0.432 | 24.151 ± 0.415 | 23.829 ± 0.602 | |
| Baq | 250 | 17.476 ± 0.398 | 17.912 ± 0.690 | 24.428 ± 0.430 | 24.123 ± 0.589 |
| 500 | 18.229 ± 0.445 | 18.377 ± 0.848 | 24.552 ± 0.583 | 24.216 ± 0.669 | |
| 100 | 17.677 ± 0.410 | 17.611 ± 0.736 | 23.438 ± 0.527 | 23.716 ± 0.756 | |
| Laq | 250 | 17.735 ± 0.537 | 17.677 ± 0.696 | 24.204 ± 0.450 | 23.757 ± 0.659 |
| 500 | 18.290 ± 0.544 | 17.873 ± 0.638 | 24.336 ± 0.391 | 24.123 ± 0.662 | |
| 100 | 17.480 ± 0.371 | 17.631 ± 0.691 | 24.282 ± 0.452 | 23.910 ± 0.485 | |
| Bet | 250 | 17.972 ± 0.537 | 17.918 ± 0.718 | 24.457 ± 0.677 | 24.171 ± 0.746 |
| 500 | 18.650 ± 0.468 a | 18.803 ± 0.786 | 25.102 ± 0.521 a | 24.620 ± 0.625 | |
| 100 | 17.538 ± 0.430 | 17.439 ± 0.352 | 23.673 ± 0.525 | 23.738 ± 0.507 | |
| Let | 250 | 17.426 ± 0.309 | 18.200 ± 0.811 | 23.962 ± 0.525 | 23.943 ± 0.642 |
| 500 | 18.321 ± 0.578 | 18.605 ± 0.542 | 24.287 ± 0.533 | 24.022 ± 0.588 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duchnowicz, P.; Pilarski, R.; Michałowicz, J.; Bukowska, B. Changes in Human Erythrocyte Membrane Exposed to Aqueous and Ethanolic Extracts from Uncaria tomentosa. Molecules 2021, 26, 3189. https://doi.org/10.3390/molecules26113189
Duchnowicz P, Pilarski R, Michałowicz J, Bukowska B. Changes in Human Erythrocyte Membrane Exposed to Aqueous and Ethanolic Extracts from Uncaria tomentosa. Molecules. 2021; 26(11):3189. https://doi.org/10.3390/molecules26113189
Chicago/Turabian StyleDuchnowicz, Piotr, Radosław Pilarski, Jaromir Michałowicz, and Bożena Bukowska. 2021. "Changes in Human Erythrocyte Membrane Exposed to Aqueous and Ethanolic Extracts from Uncaria tomentosa" Molecules 26, no. 11: 3189. https://doi.org/10.3390/molecules26113189
APA StyleDuchnowicz, P., Pilarski, R., Michałowicz, J., & Bukowska, B. (2021). Changes in Human Erythrocyte Membrane Exposed to Aqueous and Ethanolic Extracts from Uncaria tomentosa. Molecules, 26(11), 3189. https://doi.org/10.3390/molecules26113189