
Chemical Composition, Antipathogenic and Cytotoxic Activity of the Essential Oil Extracted from Amorpha fruticosa Fruits

 ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Results
2.1. Extraction of A. fruticosa Essential Oil
2.2. GC-MS Analysis of Essential Oil
2.3. Antimicrobial Activity
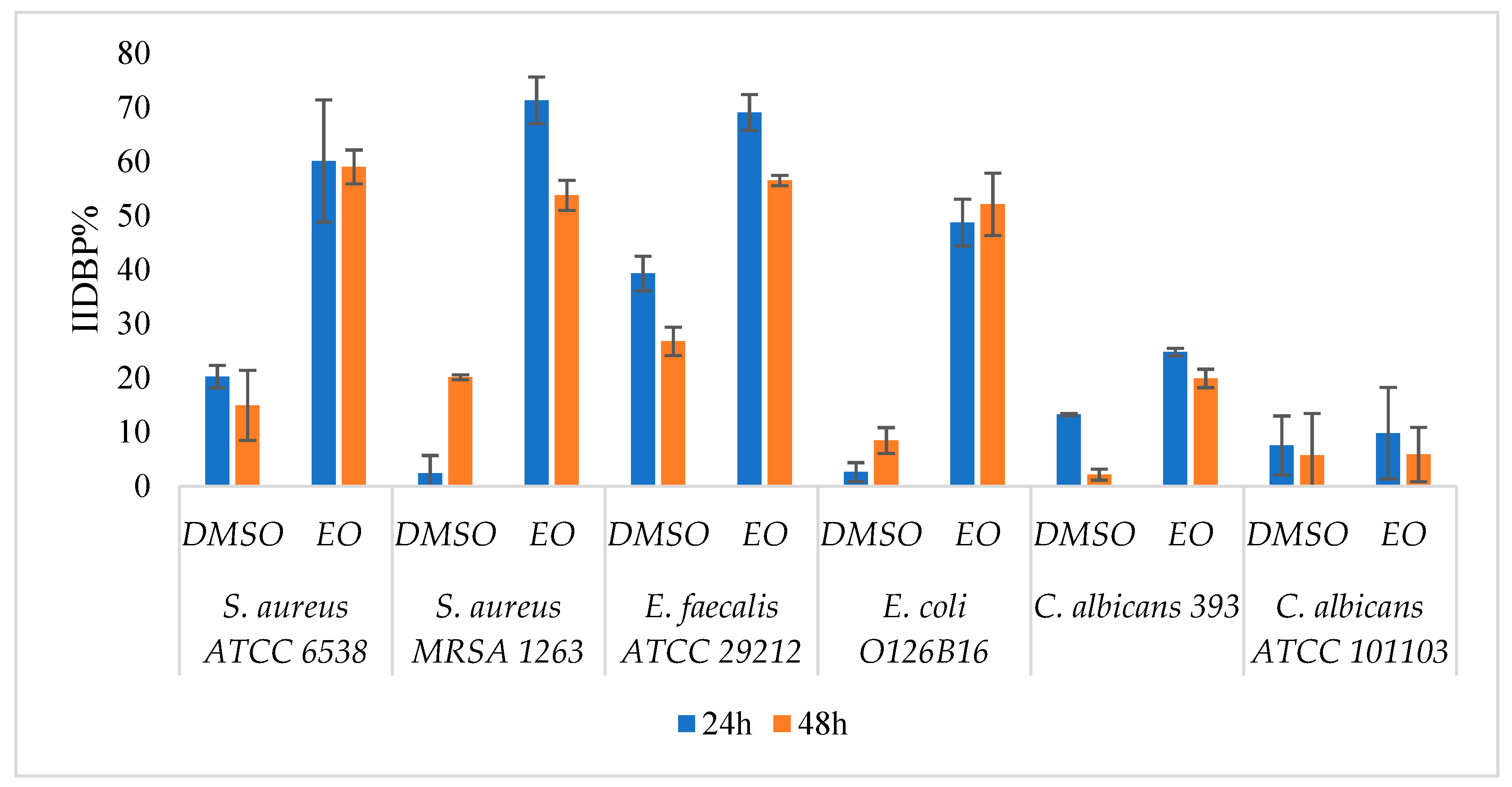
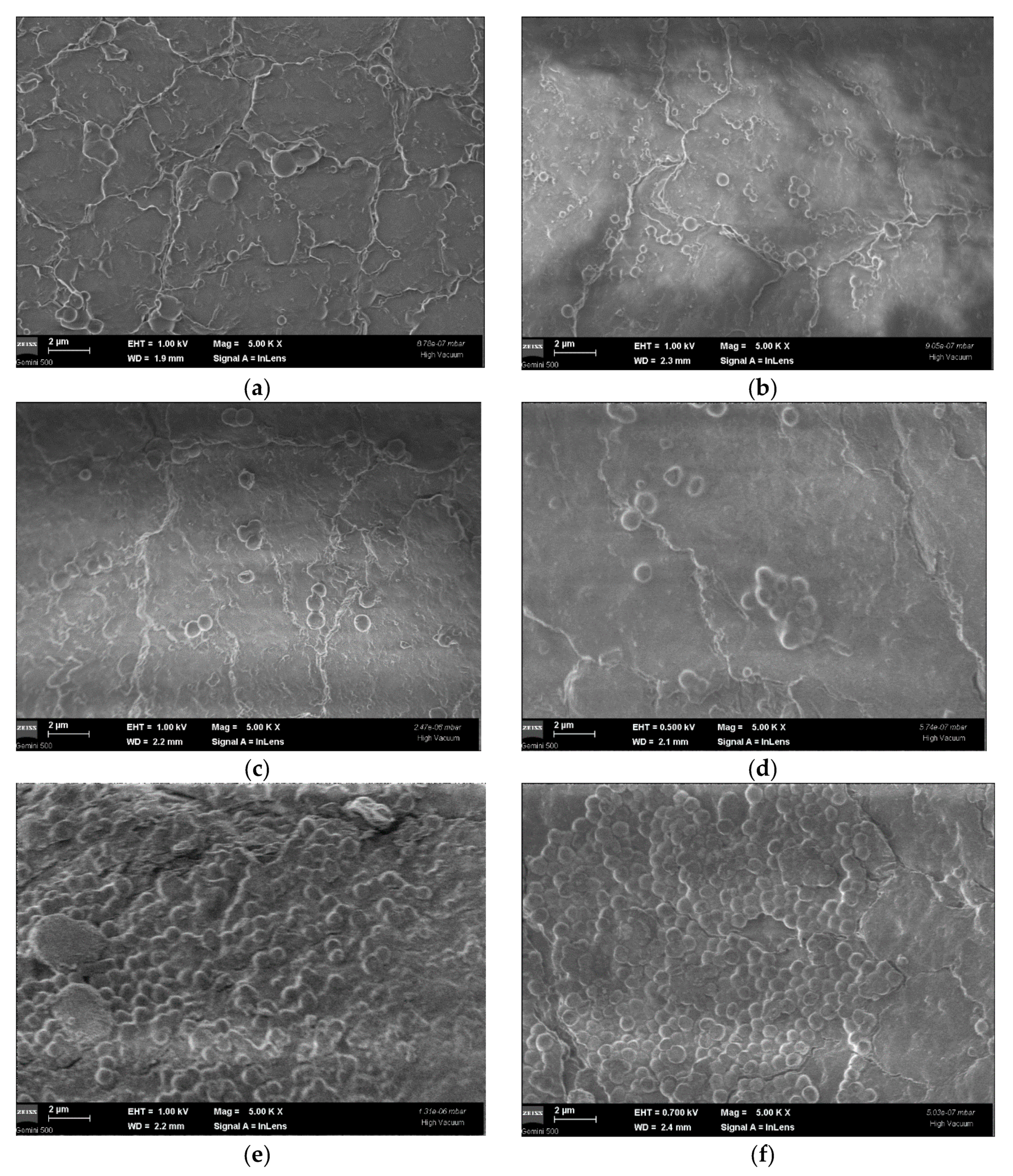
2.3.1. Eradication of Preformed Biofilms on Inert Substrates
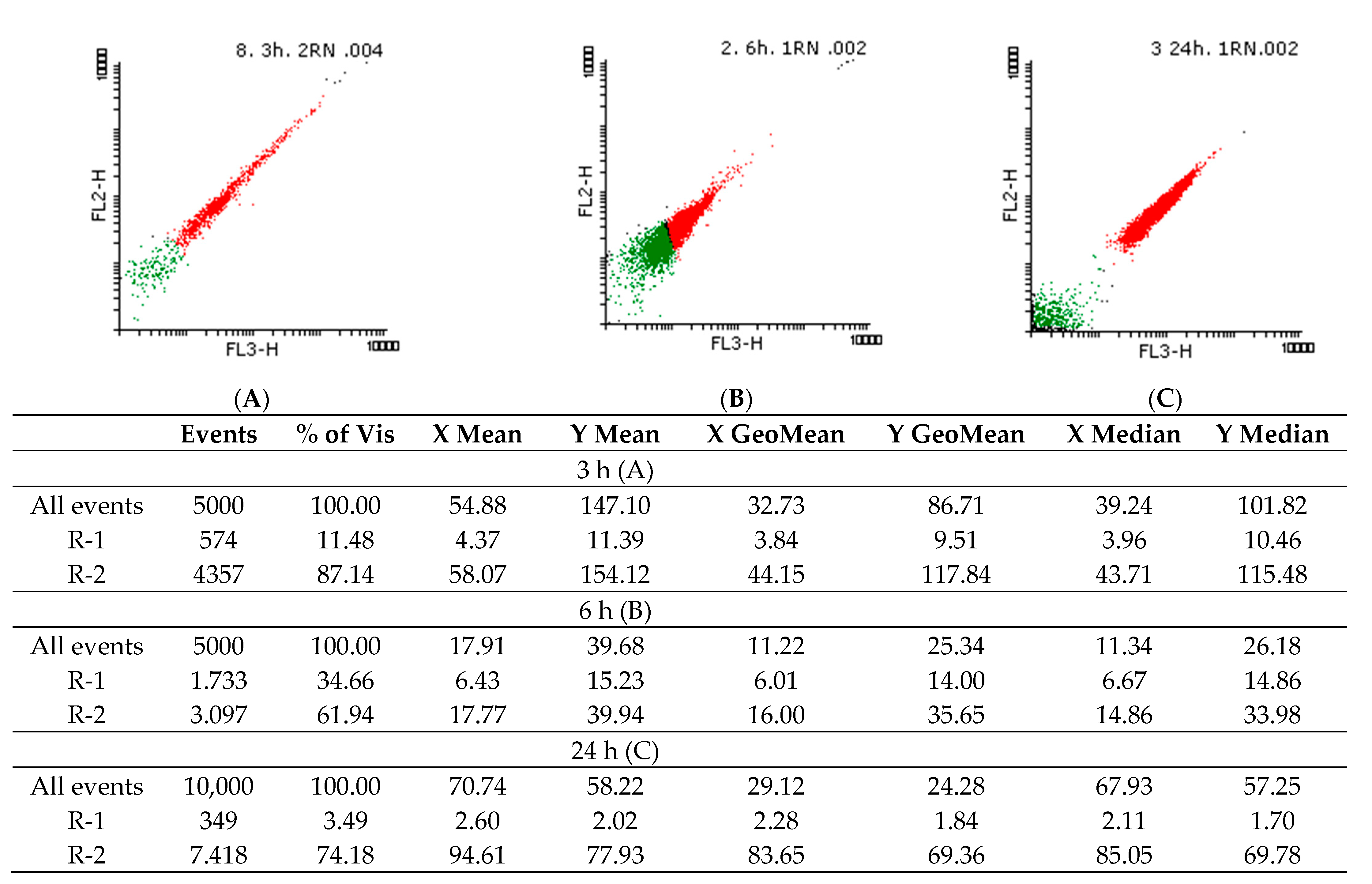
2.3.2. Flow Cytometry Evaluation of the Potential Antifungal Mechanisms
2.4. Assessment of Human Cells Viability and EO Cytotoxicity
2.5. Influence of A. fruticosa Fruits EO on Microbial Ability to Adhere to Mammalian Cells
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Chemicals and Reagents
4.3. Extraction of A. fruticosa Fruits Essential Oil
4.4. GC-MS Analysis of Essential Oil
4.5. Antimicrobial Activity Assay
4.5.1. Microbial Strains
4.5.2. Antimicrobial Activity against Planktonic Cells
4.5.3. The Influence of A. fruticosa Fruits EO on the Microbial Adherence Capacity to the Inert Surface
4.5.4. The Influence of A. fruticosa Fruits EO on Microbial Biofilms Developed on Inert Surface
4.5.5. The Influence of A. fruticosa Fruits EO on Preformed Microbial Biofilms on Foley Catheter Sections
4.5.6. Influence of A. fruticosa Fruits EO on the Hydrophobicity of Sessile and Planktonic Bacterial Cells
4.5.7. Flow Cytometry Evaluation of Viability and Intracellular Lipids Modification Using Propidium Iodide (PI) and Nile Red (NR) Staining
4.6. Assessment of Cell Viability and Cytotoxicity by MTT and LDH Assay
4.6.1. MTT Assay
4.6.2. LDH Assay
4.7. Influence of A. fruticosa Fruits EO on Microbial Ability to Adhere to Mammalian Cells
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Dumitraşcu, M.; Grigorescu, I.; Kucsicsa, G.; Dragotă, C.S.; Năstase, M. Non-Native and Native Invasive Terrestrial Plant Species in Comana Natural Park. Case-Studies: Amorpha Fruticosa and Crataegus Monogyna. Rev. Roum. Géogr./Rom. Journ. Geogr 2011, 55, 81–89. [Google Scholar]
- Anastasiu, P.; Negrean, G.; Başnou, C.; Sîrbu, C.; Oprea, A. A preliminary study on the neophytes of wetlands in Romania. In Biology Invasions—From Ecology to Conservation; Rabitsch, W., Essl, F., Klingenste, F., Eds.; Vienna University Campus: Vienna, Austria, 2008. [Google Scholar]
- Kozuharova, E.; Matkowski, A.; Wozniak, D.; Simeonova, R.; Naychov, Z.; Malainer, C.; Mocan, A.; Nabavi, S.M.; Atanasov, A.G. Amorpha fruticose—A noxious invasive alien plant in Europe or a medicinal plant against metabolic disease? Front. Pharmacol. 2017, 8, 1–17. [Google Scholar] [CrossRef]
- Novak, N.; Novak, M.; Barić, K.; Šćepanović, M.; Ivić, D. Allelopathic potential of segetal and ruderal invasive alien plants. J. Cent. Eur. Agric. 2018, 19, 408–422. [Google Scholar] [CrossRef]
- Zhuo, X.; Liu, C.; Pan, R.; Dong, X.; Li, Y. Nanocellulose Mechanically Isolated from Amorpha fruticosa Linn. ACS Sustain. Chem. Eng. 2017, 5, 4414–4420. [Google Scholar] [CrossRef]
- Kozuharova, E.; Benbassat, N.; Berkov, S.; Ionkova, I. Ailanthus altissima and Amorpha fruticose—Invasive arboreal alien plants as cheap sources of valuable essential oils. Pharmacia 2020, 67, 71–81. [Google Scholar] [CrossRef]
- Ciuvăț, A.L.; Vasile, D.; Dinu, C.; Apostol, E.; Apostol, B.; Petrițan, A.M. Valorisation possibilities of invasive indigobush (Amorpha fruticosa L.) in Romania. Rev. Silvic. Cineg. 2016, 21, 96–99. [Google Scholar]
- Lis, A.; Góra, J. Essential oil of Amorpha fruticosa L. J. Essent. Oil Res. 2001, 13, 340–342. [Google Scholar] [CrossRef]
- Chen, F.; Jia, J.; Zhang, Q.; Gu, H.; Yang, L. A modified approach for isolation of essential oil from fruit of Amorpha fruticosa Linn using microwave-assisted hydrodistillation concatenated liquid-liquid extraction. J. Chromatogr. A 2017, 1524, 254–265. [Google Scholar] [CrossRef]
- Stoyanova, A.; Georgiev, E.; Lis, A.; Majda, T.; Gora, J. Essential oil from stored fruits of Amorpha fruticosa L. J. Essent. Oil-Bearing Plants 2003, 6, 195–197. [Google Scholar] [CrossRef]
- Ivanescu, B.; Lungu, C.; Spac, A.; Tuchilus, C. Essential Oils from Amorpha fruticosa L. Fruits—Chemical Characterization and Antimicrobial Activity. Biol. Veg. 2014, 60, 33–39. [Google Scholar]
- Georgiev, E.; Stoianova, A.; Lis, A.; Gora, J. Seasonal variation of the fruit essential oil of Amorpha fruticosa L. Herba Pol. 2000, 46, 220–234. [Google Scholar]
- Kon, K.V.; Rai, M.K. Combining Essential Oils with Antibiotics and other Antimicrobial Agents to Overcome Multidrug-Resistant Bacteria. In Fighting Multidrug Resistance with Herbal Extracts, Essential Oils and Their Components; Elsevier: Amsterdam, The Netherlands, 2013; pp. 149–164. [Google Scholar] [CrossRef]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef]
- Khatoon, Z.; McTiernan, C.D.; Suuronen, E.J.; Mah, T.-F.; Alarcon, E.I. Bacterial biofilm formation on implantable devices and approaches to its treatment and prevention. Heliyon 2018, 4, e01067. [Google Scholar] [CrossRef]
- Del Sorbo, G.; Schoonbeek, H.; De Waard, M.A. Fungal Transporters Involved in Efflux of Natural Toxic Compounds and Fungicides. Fungal Genet. Biol. 2000, 30, 1–15. [Google Scholar] [CrossRef]
- Jabra-Rizk, M.A.; Falkler, W.A.; Meiller, T.F. Fungal Biofilms and Drug Resistance. Emerg. Infect. Dis. 2004, 10, 14–19. [Google Scholar] [CrossRef]
- Mihai, A.; Balotescu-Chifiriuc, C.; V, L.; Stănescu, R.; Burlibasa, M.; Ispas, D. Biofilme microbiene în medicina dentară cu referire la reabilitarea implanto-protetică, Revista de Chirurgie Oro-Maxilo-Facială și Implantologie. Rev. Chir. Oro-Maxilo-Facială Implantol. 2010, 1, 9–12. [Google Scholar]
- Patrinoiu, G.; Calderón-Moreno, J.M.; Chifiriuc, C.M.; Saviuc, C.; Birjega, R.; Carp, O. Tunable ZnO spheres with high anti-biofilm and antibacterial activity via a simple green hydrothermal route. J. Colloid Interface Sci. 2016, 462, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Mendanha, S.A.; Alonso, A. Effects of terpenes on fluidity and lipid extraction in phospholipid membranes. Biophys. Chem. 2015, 198, 45–54. [Google Scholar] [CrossRef]
- Espinoza, J.; Urzúa, A.; Sanhueza, L.; Walter, M.; Fincheira, P.; Muñoz, P.; Mendoza, L.; Wilkens, M. Essential Oil, Extracts, and Sesquiterpenes Obtained From the Heartwood of Pilgerodendron uviferum Act as Potential Inhibitors of the Staphylococcus aureus NorA Multidrug Efflux Pump. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Scandorieiro, S.; de Camargo, L.C.; Lancheros, C.A.C.; Yamada-Ogatta, S.F.; Nakamura, C.V.; de Oliveira, A.G.; Andrade, C.G.T.J.; Duran, N.; Nakazato, G.; Kobayashi, R.K.T. Synergistic and Additive Effect of Oregano Essential Oil and Biological Silver Nanoparticles against Multidrug-Resistant Bacterial Strains. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Kozuharova, E.; Benbassat, N.; Ionkova, I. The invasive alien species Amorpha fruticosa in Bulgaria and its potential as economically prospective source of valuable essential oil. Pharmacia 2020, 67, 357–362. [Google Scholar] [CrossRef]
- González, A.M.; Tracanna, M.I.; Amani, S.M.; Schuff, C.; Poch, M.J.; Bach, H.; Catalán, C.A.N. Chemical Composition, Antimicrobial and Antioxidant Properties of the Volatile Oil and Methanol Extract of Xenophyllum Poposum. Nat. Prod. Commun. 2012, 7. [Google Scholar] [CrossRef]
- Gómez-Sequeda, N.; Cáceres, M.; Stashenko, E.E.; Hidalgo, W.; Ortiz, C. Antimicrobial and Antibiofilm Activities of Essential Oils against Escherichia coli O157:H7 and Methicillin-Resistant Staphylococcus aureus (MRSA). Antibiotics 2020, 9, 730. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, A.C.; Meireles, L.M.; Lemos, M.F.; Guimarães, M.C.C.; Endringer, D.C.; Fronza, M.; Scherer, R. Antibacterial Activity of Terpenes and Terpenoids Present in Essential Oils. Molecules 2019, 24, 2471. [Google Scholar] [CrossRef] [PubMed]
- Pattnaik, S.; Subramanyam, V.R.; Bapaji, M.; Kole, C.R. Antibacterial and antifungal activity of aromatic constituents of essential oils. Microbios 1997, 89, 39–46. [Google Scholar]
- Delaquis, P. Antimicrobial activity of individual and mixed fractions of dill, cilantro, coriander and eucalyptus essential oils. Int. J. Food Microbiol. 2002, 74, 101–109. [Google Scholar] [CrossRef]
- Zarai, Z.; Kadri, A.; Ben Chobba, I.; Ben Mansour, R.; Bekir, A.; Mejdoub, H.; Gharsallah, N. The in-vitro evaluation of antibacterial, antifungal and cytotoxic properties of Marrubium vulgare L. essential oil grown in Tunisia. Lipids Health Dis. 2011, 10, 161. [Google Scholar] [CrossRef]
- Silva, D.R.; Endo, E.H.; Filho, B.P.D.; Nakamura, C.V.; Svidzinski, T.I.E.; De Souza, A.; Young, M.C.M.; Ueda-Nakamura, T.; Cortez, D.A.G. Chemical Composition and Antimicrobial Properties of Piper ovatum Vahl. Molecules 2009, 14, 1171–1182. [Google Scholar] [CrossRef]
- Pieri, F.A.; Souza, M.C.; de Castro Souza, M.C.; Vermelho, L.L.R.; Vermelho, M.L.R.; Perciano, P.G.; Vargas, F.S.; Borges, A.P.B.; da Veiga-Junior, V.F.; Moreira, M.A.S. Use of β-caryophyllene to combat bacterial dental plaque formation in dogs. BMC Vet. Res. 2016, 12, 216. [Google Scholar] [CrossRef]
- Ulubelen, A.; Topcu, G.; Eri§, C.; Sönmez, U.; Kartal, M.; Kurucu, S.; Bozok-Johansson, C. Terpenoids from Salvia sclarea. Phytochemistry 1994, 36, 971–974. [Google Scholar] [CrossRef]
- Claeson, P.; Rådström, P.; Sköld, O.; Nilsson, Å.; Höglund, S. Bactericidal effect of the sesquiterpene T-cadinol on Staphylococcus aureus. Phyther. Res. 1992, 6, 94–98. [Google Scholar] [CrossRef]
- Durgadevi, R.; Veera Ravi, A.; Alexpandi, R.; Krishnan Swetha, T.; Abirami, G.; Vishnu, S.; Karutha Pandian, S. Virulence targeted inhibitory effect of linalool against the exclusive uropathogen Proteus mirabilis. Biofouling 2019, 35, 508–525. [Google Scholar] [CrossRef] [PubMed]
- Raut, J.S.; Shinde, R.B.; Chauhan, N.M.; Karuppayil, S.M. Terpenoids of plant origin inhibit morphogenesis, adhesion, and biofilm formation by Candida albicans. Biofouling 2013, 29, 87–96. [Google Scholar] [CrossRef]
- Xiong, G.; Ji, W.; Wang, F.; Zhang, F.; Xue, P.; Cheng, M.; Sun, Y.; Wang, X.; Zhang, T. Quercetin Inhibits Inflammatory Response Induced by LPS from Porphyromonas gingivalis in Human Gingival Fibroblasts via Suppressing NF- κ B Signaling Pathway. Biomed Res. Int. 2019, 2019, 6282635. [Google Scholar] [CrossRef] [PubMed]
- Cerca, N.; Pier, G.B.; Vilanova, M.; Oliveira, R.; Azeredo, J. Quantitative analysis of adhesion and biofilm formation on hydrophilic and hydrophobic surfaces of clinical isolates of Staphylococcus epidermidis. Res. Microbiol. 2005, 156, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Lather, P.; Mohanty, A.K.; Jha, P.; Garsa, A.K. Contribution of Cell Surface Hydrophobicity in the Resistance of Staphylococcus aureus against Antimicrobial Agents. Biochem. Res. Int. 2016, 2016, 1091290. [Google Scholar] [CrossRef]
- Rubini, D.; Vedha Hari, B.N.; Nithyanand, P. Chitosan coated catheters alleviates mixed species biofilms of Staphylococcus epidermidis and Candida albicans. Carbohydr. Polym. 2021, 252, 117192. [Google Scholar] [CrossRef]
- Mayrs, E.B.C. British Pharmacopoeia, Appendix 9 A238; Medicines & Healthcare products Regulatory Agency: London, UK, 2017; Volume IV. [Google Scholar]
- Koo, I.; Kim, S.; Zhang, X. Comparative analysis of mass spectral matching-based compound identification in gas chromatography–mass spectrometry. J. Chromatogr. A 2013, 1298, 132–138. [Google Scholar] [CrossRef]
- van Den Dool, H.; Kratz, P.D. A generalization of the retention index system including linear temperature programmed gas—liquid partition chromatography. J. Chromatogr. A 1963, 11, 463–471. [Google Scholar] [CrossRef]
- Özkalp, B.; Sevgi, F.; Özcan, M.; Özcan, M.M. The antibacterial activity of essential oil of oregano (Origanum vulgare L.). J. Food, Agric. Environ. 2010, 8, 272–274. [Google Scholar]
- Marinas, I.; Oprea, E.; Geana, E.-I.; Chifiriuc, C.; Lazar, V. Antimicrobial and antioxidant activity of the vegetative and reproductive organs of Robinia pseudoacacia. J. Serbian Chem. Soc. 2014, 79, 1363–1378. [Google Scholar] [CrossRef]
- Oh, S.Y.; Yun, W.; Lee, J.H.; Lee, C.H.; Kwak, W.K.; Cho, J.H. Effects of essential oil (blended and single essential oils) on anti-biofilm formation of Salmonella and Escherichia coli. J. Anim. Sci. Technol. 2017, 59, 4. [Google Scholar] [CrossRef] [PubMed]
- Millezi, A.F.; Costa, K.A.D.; Oliveira, J.M.; Lopes, S.P.; Pereira, M.O.; Piccoli, R.H. Antibacterial and anti-biofilm activity of cinnamon essential oil and eugenol. Ciência Rural 2019, 49. [Google Scholar] [CrossRef]
- Anghel, I.; Grumezescu, A.; Holban, A.; Ficai, A.; Anghel, A.; Chifiriuc, M. Biohybrid Nanostructured Iron Oxide Nanoparticles and Satureja hortensis to Prevent Fungal Biofilm Development. Int. J. Mol. Sci. 2013, 14, 18110–18123. [Google Scholar] [CrossRef]
- Gogra, A.B.; Yao, J.; Sandy, E.H.; Zheng, S.; Zaray, G.; Koroma, B.M.; Hui, Z. Cell surface hydrophobicity (CSH) of Escherichia coli, Staphylococcus aureus and Aspergillus niger and the biodegradation of Diethyl Phthalate (DEP) via Microcalorimetry. J. Am. Sci. 2010, 6, 78–88. [Google Scholar]
- Qiao, G.; Li, H.; Xu, D.-H.; Il Park, S. Modified a Colony Forming Unit Microbial Adherence to Hydrocarbons Assay and Evaluated Cell Surface Hydrophobicity and Biofilm Production of Vibrio scophthalmi. J. Bacteriol. Parasitol. 2012, 03. [Google Scholar] [CrossRef]
- Ramani, R.; Chaturvedi, V. Flow Cytometry Antifungal Susceptibility Testing of Pathogenic Yeasts other than Candida albicans and Comparison with the NCCLS Broth Microdilution Test. Antimicrob. Agents Chemother. 2000, 44, 2752–2758. [Google Scholar] [CrossRef]
- Guzmán, H.M.; de la Jara Valido, A.; Duarte, L.C.; Presmanes, K.F. Estimate by means of flow cytometry of variation in composition of fatty acids from Tetraselmis suecica in response to culture conditions. Aquac. Int. 2010, 18, 189–199. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Weyermann, J.; Lochmann, D.; Zimmer, A. A practical note on the use of cytotoxicity assays. Int. J. Pharm. 2005, 288, 369–376. [Google Scholar] [CrossRef]
- Holban, A.M.; Chifiriuc, M.C.; Cotar, A.I.; Bleotu, C.; Grumezescu, A.M.; Banu, O.; Lazar, V. Virulence markers in pseudomonas aeruginosa isolates from hospitalacquired infections occurred in patients with underlying cardiovascular disease. Rom. Biotechnol. Lett. 2013, 18, 7243–7254. [Google Scholar]
- Fleiszig, S.M.J.; Zaidi, T.S.; Preston, M.J.; Grout, M.; Evans, D.J.; Pier, G.B. Relationship between cytotoxicity and corneal epithelial cell invasion by clinical isolates of Pseudomonas aeruginosa. Infect. Immun. 1996, 64, 2288–2294. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | RT a | RI b | Compound | RA c |
|---|---|---|---|---|
| 1 | 7.85 | 1044 | trans-β-Ocimene (E) | 1.65 |
| 2 | 8.03 | 1055 | cis-β-Ocimene (Z) | 0.50 |
| 3 | 8.93 | 1108 | Linalool | 2.15 |
| 4 | 10.91 | 1263 | Citronellol | 0.51 |
| 5 | 12.73 | 1365 | α-Cubebene | 0.59 |
| 6 | 13.07 | 1390 | α-Ylangene | 2.35 |
| 7 | 13.12 | 1393 | α-Copaene | 6.46 |
| 8 | 13.60 | 1431 | α-Gurjunene | 0.94 |
| 9 | 13.75 | 1443 | β-Caryophyllene | 6.25 |
| 10 | 13.86 | 1451 | β-Gurjunene (calarene) | 1.24 |
| 11 | 14.01 | 1462 | α-Guaiene | 1.98 |
| 12 | 14.19 | 1477 | Humulene (α-caryophyllene) | 1.22 |
| 13 | 14.43 | 1496 | γ-Muurolene | 12.79 |
| 14 | 14.53 | 1504 | Germacrene D | 0.45 |
| 15 | 14.67 | 1516 | α-Amorphene | 4.77 |
| 16 | 14.72 | 1520 | α-Muurolene | 12.54 |
| 17 | 14.92 | 1537 | γ-Cadinene | 7.86 |
| 18 | 15.01 | 1544 | δ-Cadinene | 20.09 |
| 19 | 15.19 | 1560 | Cadinadiene-1,4 | 1.87 |
| 20 | 15.28 | 1567 | α-Calacorene | 1.34 |
| 21 | 15.84 | 1615 | Caryophyllene oxide | 0.63 |
| 22 | 16.35 | 1660 | γ-Eudesmol | 1.35 |
| 23 | 16.45 | 1669 | τ-Cadinol | 0.93 |
| 24 | 16.62 | 1684 | α-Eudesmol | 4.46 |
| Monoterpene hydrocarbons | 2.15 | |||
| Oxygenated monoterpenes | 2.66 | |||
| Sesquiterpene hydrocarbons | 82.74 | |||
| Oxygenated sesquiterpenes | 7.37 | |||
| Total | 94.92 |
| Strains | DI a (mm) | MIC b (mg/mL) | MBEC c (mg/mL) | |||||
|---|---|---|---|---|---|---|---|---|
| EO d | ATB e | EO d | L f | DMSO g | EO d | L f | DMSO g | |
| S. aureus ATCC 6538 | 13.00 ± 1.41 | 9.00 ± 0.00 h | 3.69 | 2.48 | >29.50 | 1.84 | 1.24 | >29.50 |
| S. aureus MRSA 1263 | 7.50 ± 0.71 | 28.00 ± 0.00 i | 1.84 | 9.91 | >29.50 | 0.92 | 4.96 | >29.50 |
| B. subtilis 12488 | 8.50 ± 0.71 | 17.33 ± 0.58 j | 7.38 | 4.96 | 7.38 | 0.92 | 0.31 | 1.84 |
| B. subtilis ATCC 6683 | 11.50 ± 0.71 | 20.50 ± 2.12 j | 7.38 | - | 7.38 | 0.92 | - | 1.84 |
| E. faecalis ATCC 29212 | 9.50 ± 0.71 | 27.50 ± 2.10 i | 7.38 | 9.91 | >29.50 | 3.69 | 2.48 | 29.50 |
| P. aeruginosa ATCC 27853 | 7.00 ± 0.00 | 13.25 ± 1.26 k | 14.75 | 4.96 | 29.50 | 14.75 | 2.48 | 14.75 |
| P. aeruginosa 134202 | - | 18.33 ± 1.53 k | 29.50 | ˃9.91 | >29.50 | 29.50 | - | 29.50 |
| K. pneumoniae 11 | 7.00 ± 0.00 | 13.67 ± 1.15 l | >29.50 | 2.48 | >29.50 | >29.50 | - | >29.50 |
| K. pneumoniae ATCC 134202 | 7.50 ± 0.71 | 12.50 ± 0.71 l | 29.50 | 4.96 | 29.50 | 14.75 | 2.48 | 14.75 |
| E. coli ATCC 13202 | - | 24.50 ± 6.36 m | >29.50 | 2.48 | >29.50 | >29.50 | 0.62 | >29.50 |
| E. coli O126B16 | 7.50 ± 0.71 | 9.00 ± 0.00 n | 14.75 | 9.91 | >29.50 | 14.75 | 4.96 | 29.50 |
| A. baumannii 77 sc | 7.00 ± 0.00 | 19.00 ± 0.00 p | 29.50 | 0.62 | 29.50 | 29.50 | - | 29.50 |
| C. famata 945 | - | - | 14.75 | ˃9.91 | 14.75 | 14.75 | - | 14.75 |
| C. albicans 393 | 8.00 ± 0.00 | - | 7.38 | 0.62 | 14.75 | 7.38 | 0.31 | 14.75 |
| C. utilis 15 | - | - | >29.50 | 0.62 | >29.50 | >29.50 | 0.31 | >29.50 |
| C. famata CMGBy.14 | - | - | >29.50 | 1.24 | >29.50 | >29.50 | 0.62 | >29.50 |
| C. albicans ATCC 101103 | ± r | - | 7.38 | 1.24 | 14.75 | 0.92 | - | 7.38 |
| IIBG% a | MICx3 b (mg/mL) | MICx5 c (mg/mL) | |||
|---|---|---|---|---|---|
| EO d | DMSO e | EO d | DMSO e | ||
| S. aureus ATCC 6538 * | 24 h | 71.05 ± 0.84 | 48.88 ± 5.35 | 80.12 ± 1.9 | 51.32 ± 4.56 |
| 48 h | 73.35 ± 0.98 | 17.94 ± 3.68 | 77.57 ± 1.51 | 37.21 ± 1.86 | |
| S. aureus MRSA 1263 * | 24 h | 76.49 ± 1.98 | 43.15 ± 3.25 | 77.11 ± 7.05 | 46.25 ± 5.62 |
| 48 h | 79.39 ± 1.17 | 38.65 ± 2.06 | 86.38 ± 3.05 | 48.24 ± 14.11 | |
| E. faecalis ATCC 29212 * | 24 h | 84.09 ± 5.42 | 42.09 ± 4.43 | 94.97 ± 6.8 | 62.61 ± 10.67 |
| 48 h | 48.21 ± 4.30 | 8.31 ± 5.79 | 92.17 ± 9.79 | 47.51 ± 3.70 | |
| E. coli O126B16 * | 24 h | 83.46 ± 4.67 | 52.13 ± 2.79 | 82.52 ± 10.32 | 60.11 ± 0.38 |
| 48 h | 76.67 ± 2.57 | 59.64 ± 0.59 | 77.96 ± 2.93 | 64.92 ± 3.68 | |
| C. albicans 393 * | 24 h | 69.00 ± 9.51 | 41.27 ± 5.21 | 77.39 ± 2.84 | 50.58 ± 2.60 |
| 48 h | 76.93 ± 1.95 | 57.40 ± 3.05 | 75.44 ± 6.1 | 65.37 ± 2.62 | |
| C. albicans ATCC 101103 | 24 h | 61.95 ± 0.49 | 62.94 ± 3.32 | 80.12 ± 1.9 | 66.99 ± 3.42 |
| 48 h | 55.13 ± 6.94 | 56.37 ± 2.59 | 77.57 ± 1.51 | 61.55 ± 2.04 | |
| Strains | S. aureus ATCC 6538 | MRSA 1263 | E. faecalis ATCC 29212 | E. coli O126B16 |
|---|---|---|---|---|
| AICS a (%) | ||||
| EO b | 70.08 | 79.73 | 61.96 | 57.30 |
| DMSO c | 100 | 100 | 96.25 | 98.70 |
| Positive control d | 99.54 | 100 | 98.70 | 100 |
| AP e | ||||
| EO | 1 | 3 | 2 | 1 |
| DMSO | 3 | 3 | 1 | 2 |
| Positive control d | 3 | 3 | 1 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marinas, I.C.; Oprea, E.; Buleandra, M.; Badea, I.A.; Tihauan, B.M.; Marutescu, L.; Angheloiu, M.; Matei, E.; Chifiriuc, M.C. Chemical Composition, Antipathogenic and Cytotoxic Activity of the Essential Oil Extracted from Amorpha fruticosa Fruits. Molecules 2021, 26, 3146. https://doi.org/10.3390/molecules26113146
Marinas IC, Oprea E, Buleandra M, Badea IA, Tihauan BM, Marutescu L, Angheloiu M, Matei E, Chifiriuc MC. Chemical Composition, Antipathogenic and Cytotoxic Activity of the Essential Oil Extracted from Amorpha fruticosa Fruits. Molecules. 2021; 26(11):3146. https://doi.org/10.3390/molecules26113146
Chicago/Turabian StyleMarinas, Ioana Cristina, Eliza Oprea, Mihaela Buleandra, Irinel Adriana Badea, Bianca Maria Tihauan, Luminita Marutescu, Marin Angheloiu, Elena Matei, and Mariana Carmen Chifiriuc. 2021. "Chemical Composition, Antipathogenic and Cytotoxic Activity of the Essential Oil Extracted from Amorpha fruticosa Fruits" Molecules 26, no. 11: 3146. https://doi.org/10.3390/molecules26113146
APA StyleMarinas, I. C., Oprea, E., Buleandra, M., Badea, I. A., Tihauan, B. M., Marutescu, L., Angheloiu, M., Matei, E., & Chifiriuc, M. C. (2021). Chemical Composition, Antipathogenic and Cytotoxic Activity of the Essential Oil Extracted from Amorpha fruticosa Fruits. Molecules, 26(11), 3146. https://doi.org/10.3390/molecules26113146