
Geranyl Functionalized Materials for Site-Specific Co-Immobilization of Proteins




Abstract
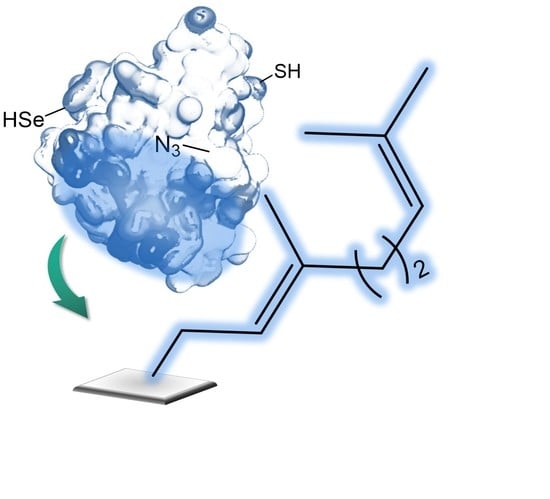
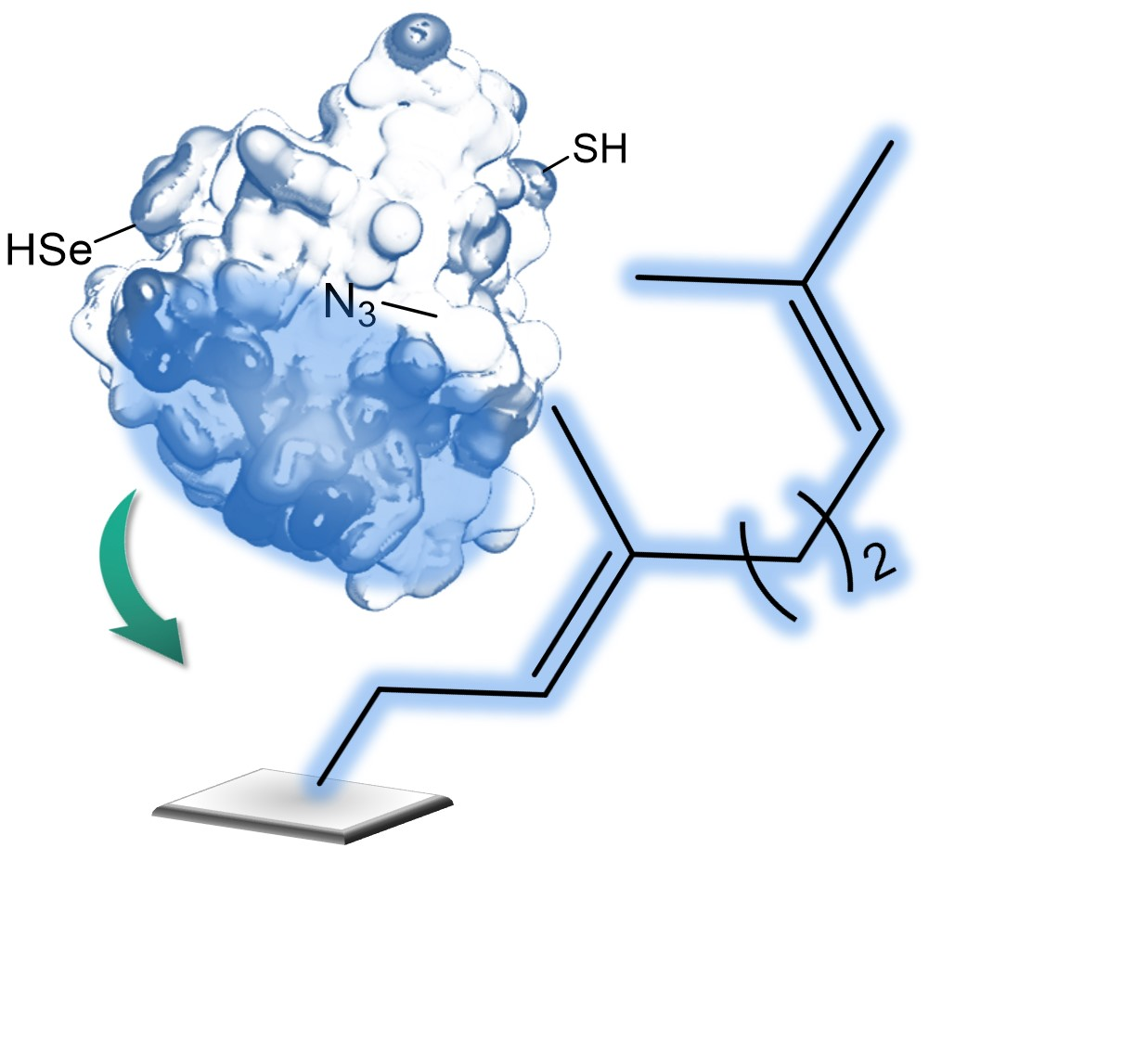
1. Introduction
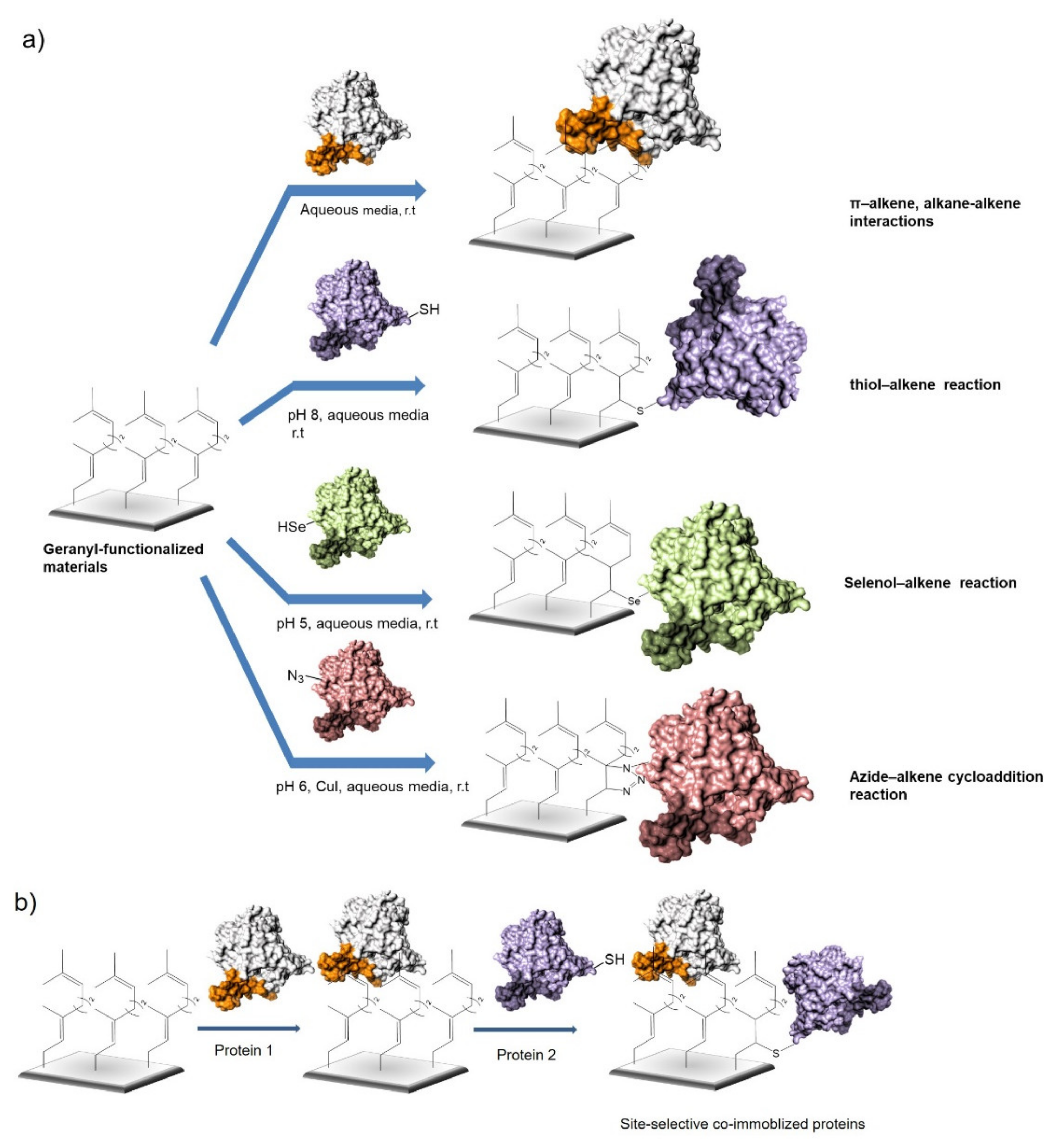
2. Results and Discussion
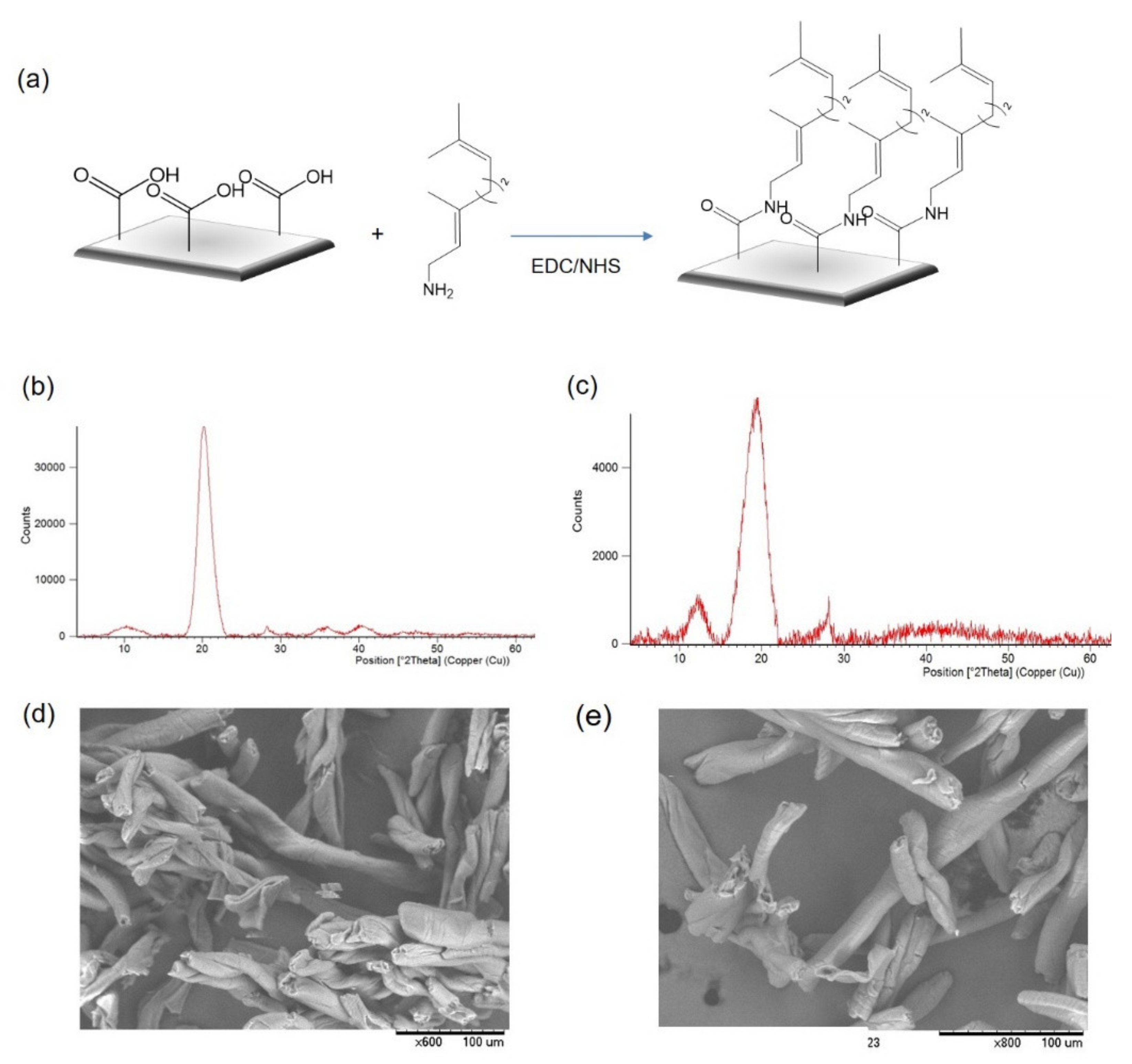
2.1. Synthesis of Geranyl-Functionalized Materials
2.2. Selective Immobilization of Enzymes on Geranyl Support by Hydrophobic Interactions
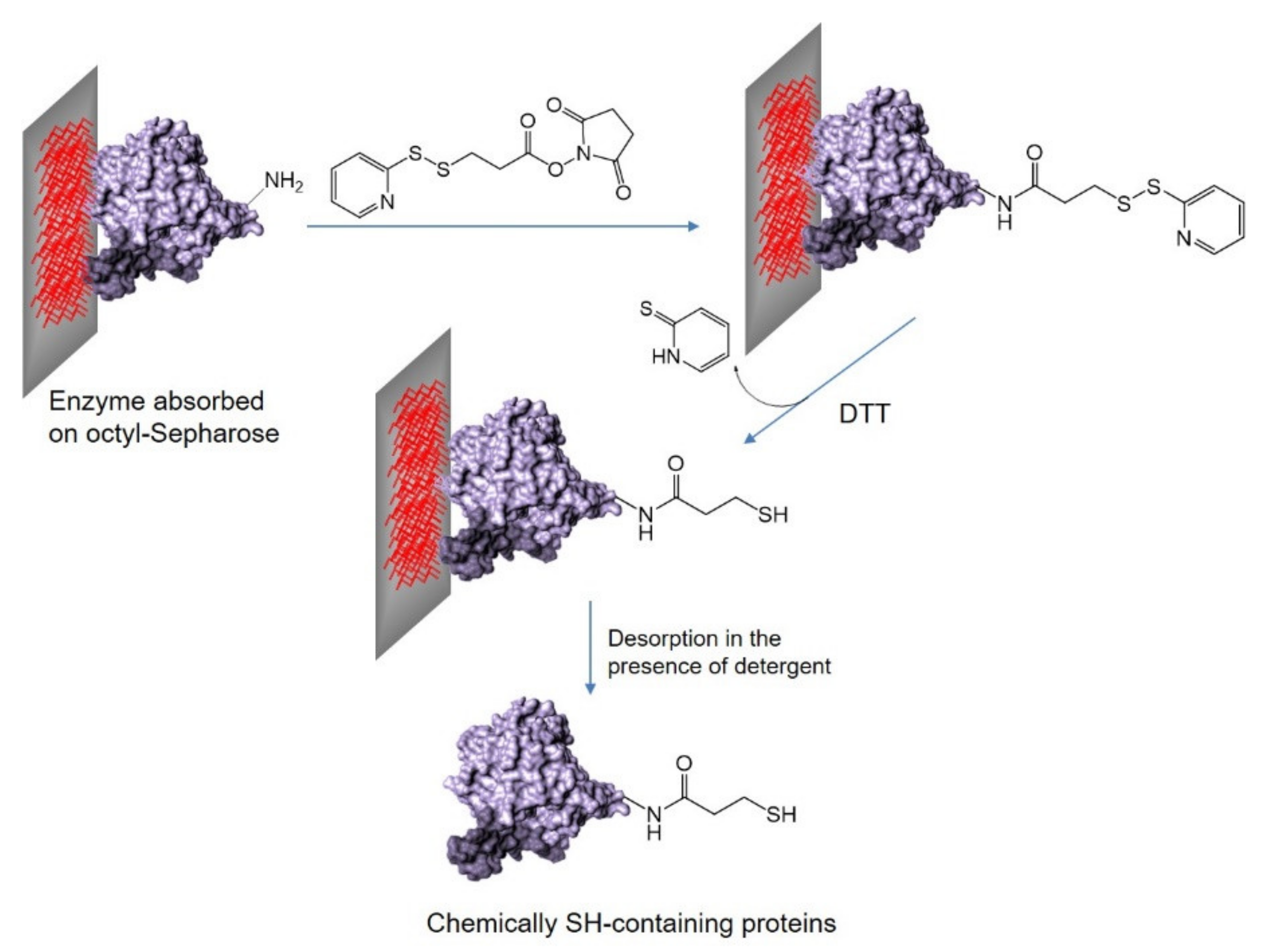
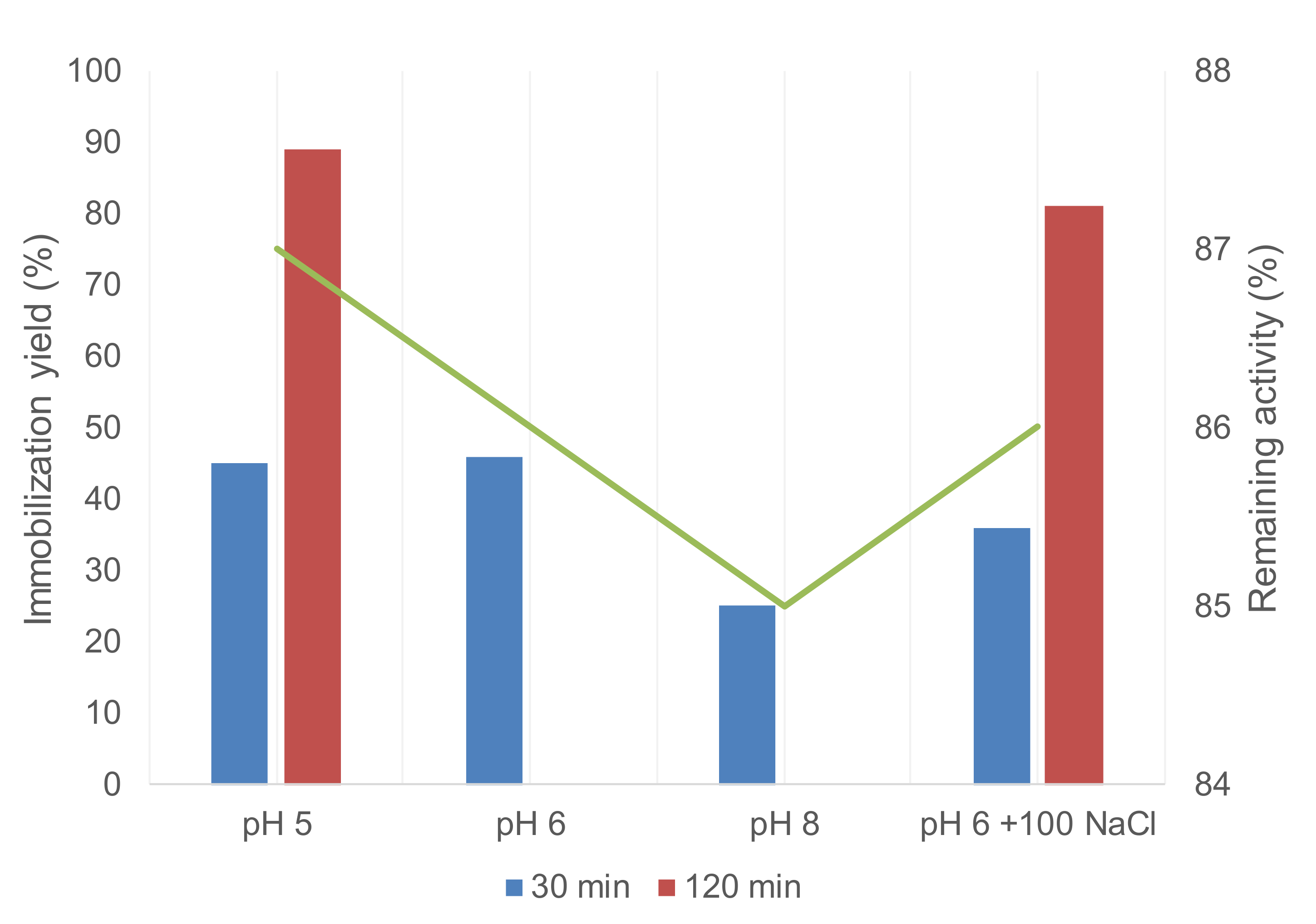
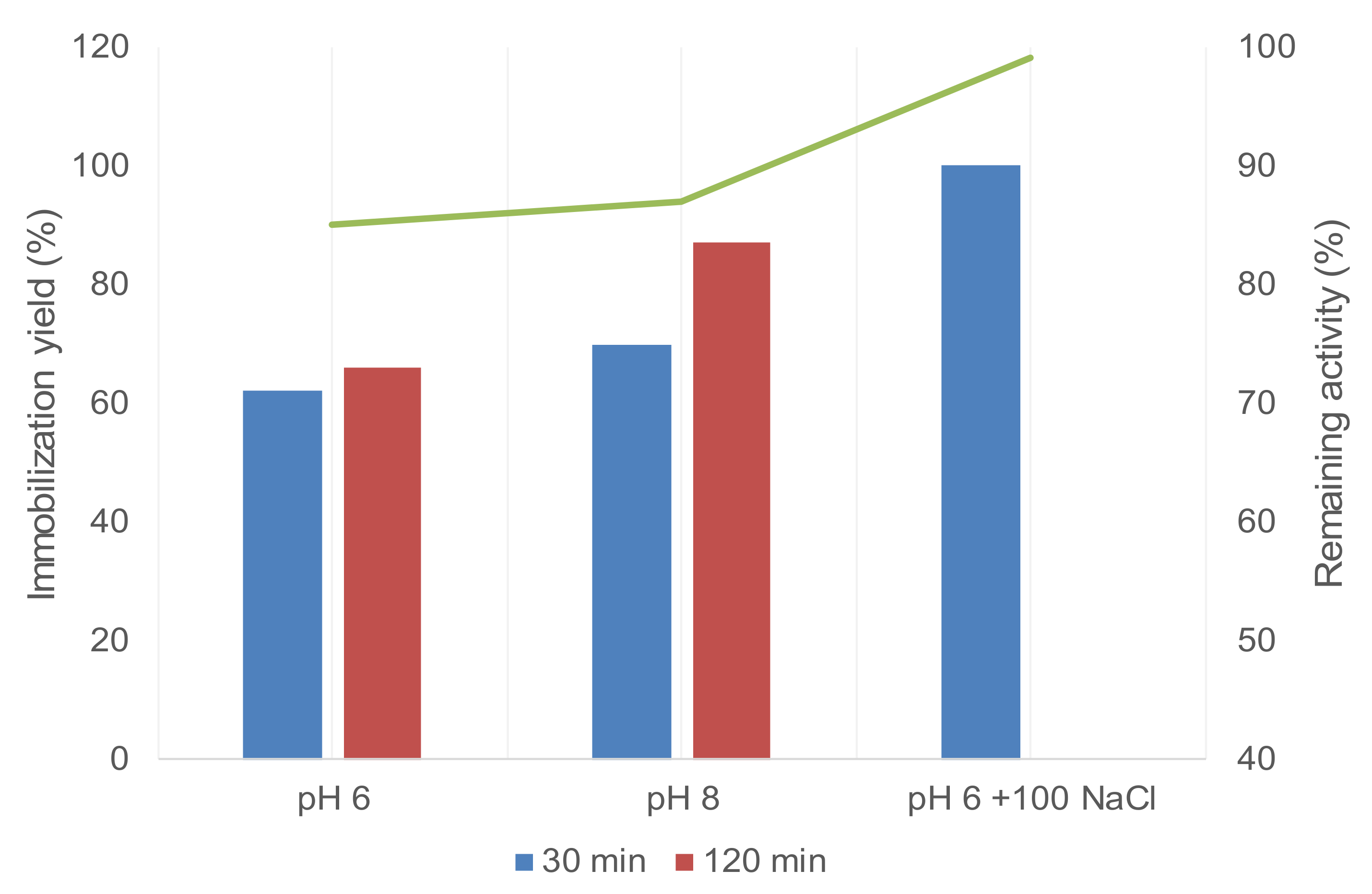
2.3. Selective Covalent Immobilization of Enzymes on Geranyl Support by Thiol-Alkene Reaction
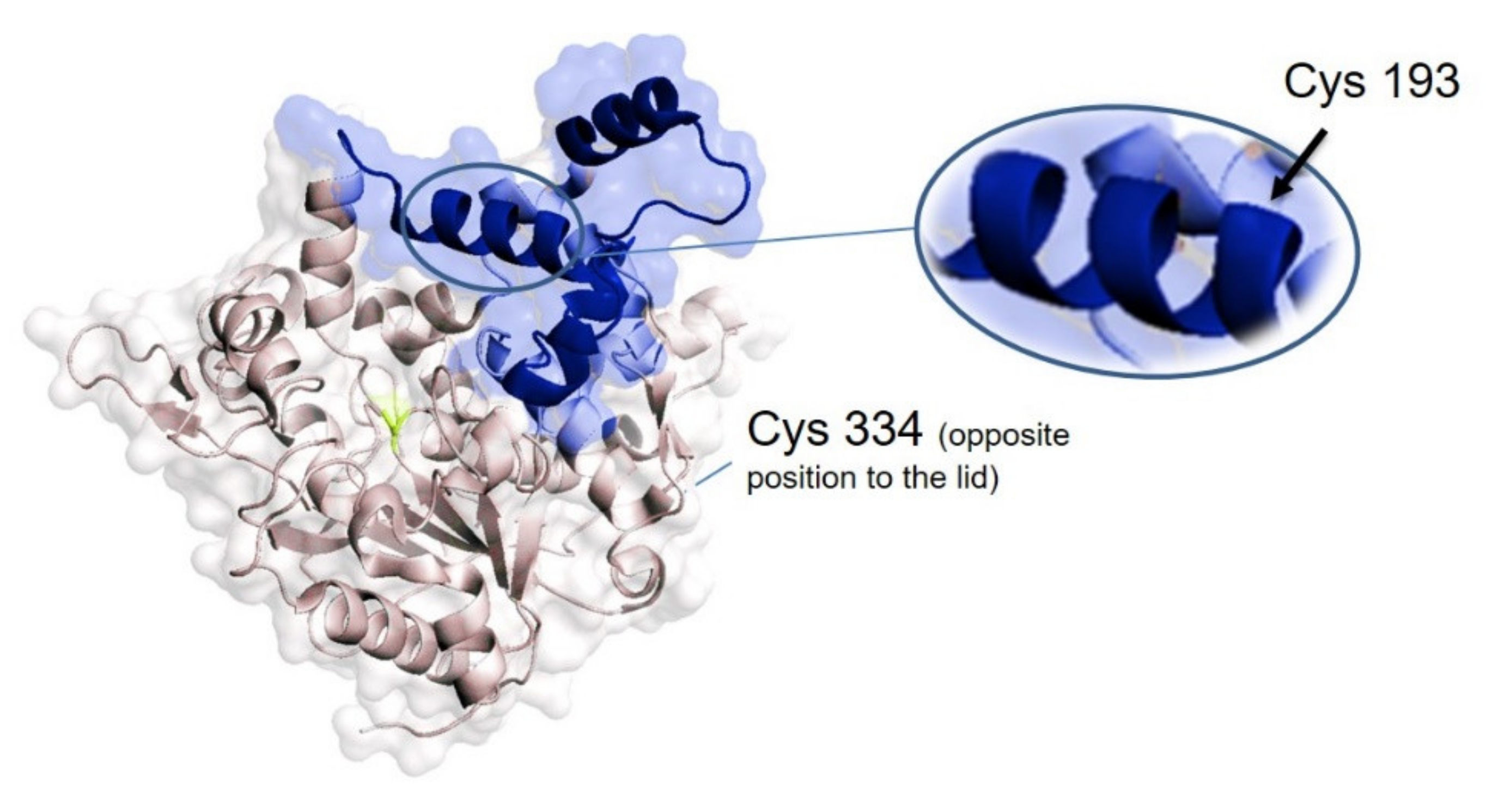
2.4. Selective Covalent Immobilization of Seleno Enzymes on Geranyl Support by Selenol-Alkene Reaction
2.5. Selective Covalent Immobilization of Azido Enzymes on Geranyl Support by Azide-Cycloaddition Reaction
3. Materials and Methods
3.1. General
3.2. Preparation of the Geranyl-Functionalized Support
3.3. Enzymatic Activity Assays
3.3.1. Lipase Activity Assay
3.3.2. Tyrosinase Activity Assay
3.3.3. Glycosidase Activity Assay
3.4. Immobilization of Enzymes on Geranyl-Support by Hydrophobic Interactions
3.5. Covalent Immobilization of SH-, Se, or N3-Containing Enzymes on Geranyl Support
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Barsan, M.M.; Enache, T.A.; Preda, N.; Stan, G.; Apostol, N.G.; Matei, E.; Kuncser, A.; Diculescu, V.C. Direct Immobilization of Biomolecules through Magnetic Forces on Ni Electrodes via Ni Nanoparticles: Applications in Electrochemical Biosensors. ACS Appl. Mater. Interfaces 2019, 11, 19867–19877. [Google Scholar] [CrossRef]
- Smith, S.; Goodge, K.; Delaney, M.; Struzyk, A.; Tansey, N.; Frey, M. A Comprehensive Review of the Covalent Immobilization of Biomolecules onto Electrospun Nanofibers. Nanomaterials 2020, 10, 2142. [Google Scholar] [CrossRef]
- Garifullina, A.; Shen, A.Q. Optimized Immobilization of Biomolecules on Nonspherical Gold Nanostructures for Efficient Localized Surface Plasmon Resonance Biosensing. Anal. Chem. 2019, 91, 15090–15098. [Google Scholar] [CrossRef]
- Basso, A.; Serban, S. Industrial applications of immobilized enzymes—A review. Mol. Catal. 2019, 479, 110607. [Google Scholar] [CrossRef]
- Chapman, J.; Ismail, A.E.; Dinu, C.Z. Industrial Applications of Enzymes: Recent Advances, Techniques, and Outlooks. Catalysts 2018, 8, 238. [Google Scholar] [CrossRef]
- Garcia-Galan, C.; Berenguer-Murcia, A.; Fernandez-Lafuente, R.; Rodrigues, R.C. Potential of different enzyme immobilization strategies to improve enzyme performance. Adv. Synth. Catal. 2011, 353, 2885–2904. [Google Scholar] [CrossRef]
- Guzik, U.; Hupert-Kocurek, K.; Wojcieszynska, D. Immobilization as a strategy for improving enzyme properties- Application to oxidoreductases. Molecules 2014, 19, 8995–9018. [Google Scholar] [CrossRef]
- Mateo, C.; Palomo, J.M.; Fernandez-Lorente, G.; Guisan, J.M.; Fernandez-Lafuente, R. Improvement of enzyme activity, stability and selectivity via immobilization techniques. Enzyme Microb. Technol. 2007, 40, 1451–1463. [Google Scholar] [CrossRef]
- Filice, M.; Guisan, J.M.; Terreni, M.; Palomo, J.M. Regioselective monodeprotection of peracetylated carbohydrates. Nat. Protoc. 2012, 7, 1783–1796. [Google Scholar] [CrossRef]
- Palomo, J.M.; Filice, M. New emerging bio-catalysts design in biotransformations. Biotechnol. Adv. 2015, 33, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Adlercreutz, P. Immobilisation and application of lipases in organic media. Chem. Soc. Rev. 2013, 42, 6406–6436. [Google Scholar] [CrossRef] [PubMed]
- Filice, M.; Molina, M.; Benaiges, M.D.; Abian, O.; Valero, F.; Palomo, J.M. Solid-surface activated recombinant Rhizopous oryzae lipase expressed in Pichia Pastoris and chemically modified variants as efficient catalysts in the synthesis of hydroxy monodeprotected glycals. Catal. Sci. Technol. 2017, 7, 1766–1775. [Google Scholar] [CrossRef]
- Wu, S.; Zhou, Y.; Gerngross, D.; Jeschek, M.; Ward, T.R. Chemo-enzymatic cascades to produce cycloalkenes from bio-based resources. Nat. Commun. 2019, 10, 5060. [Google Scholar] [CrossRef] [PubMed]
- Losada-Garcia, N.; Cabrera, Z.; Urrutia, P.; Garcia-Sanz, C.; Andreu, A.; Palomo, J.M. Recent Advances in Enzymatic and Chemoenzymatic Cascade Processes. Catalysts 2020, 10, 1258. [Google Scholar] [CrossRef]
- Ji, Q.; Wanga, B.; Tanb, J.; Zhua, L.; Li, L. Immobilized multienzymatic systems for catalysis of cascade reactions. Process Biochem. 2016, 51, 1193–1203. [Google Scholar] [CrossRef]
- Hwang, E.T.; Lee, S. Multienzymatic Cascade Reactions via Enzyme Complex by Immobilization. ACS Catal. 2019, 9, 4402–4425. [Google Scholar] [CrossRef]
- Schoffelen, S.; van Hest, J.C.M. Chemical approaches for the construction of multi-enzyme reaction systems. Curr. Opin. Struc. Biol. 2013, 23, 613–621. [Google Scholar] [CrossRef]
- Palomo, J.M. Artificial enzymes with multiple active sites. Curr. Opin. Green Sustain. Chem. 2021, 29, 100452. [Google Scholar] [CrossRef]
- Palomo, J.M. Nanobiohybrids: A new concept for metal nanoparticles synthesis. Chem. Commun. 2019, 55, 9583–9589. [Google Scholar] [CrossRef]
- McIntosh, J.A.; Owen, A.E. Enzyme engineering for biosynthetic cascades. Curr. Opin. Green Sustain. Chem. 2021, 29, 100448. [Google Scholar] [CrossRef]
- Ren, X.; Fasan, R. Engineered and Artificial Metalloenzymes for Selective C–H Functionalization. Curr. Opin. Green Sustain. Chem. 2021, 29, 100494. [Google Scholar] [CrossRef]
- Ren, S.Z.; Li, C.H.; Jiao, X.B.; Jia, S.R.; Jiang, Y.J.; Bilal, M.; Cui, J. Recent progress in multienzymes co-immobilization and multienzyme system applications. Chem. Eng. J. 2019, 373, 1254–1278. [Google Scholar] [CrossRef]
- Xu, K.; Chen, X.; Zheng, R.; Zheng, Y. Immobilization of Multi-Enzymes on Support Materials for Efficient Biocatalysis. Front. Bioeng. Biotechnol. 2020, 8, 660. [Google Scholar] [CrossRef]
- Zhou, F.; Luo, J.; Song, S.; Wan, Y. Nanostructured Polyphenol-Mediated Coating: A Versatile Platform for Enzyme Immobilization and Micropollutant Removal. Ind. Eng. Chem. Res. 2020, 59, 2708–2717. [Google Scholar] [CrossRef]
- Zhang, H.; Luo, J.; Li, S.; Wei, Y.; Wan, Y. Biocatalytic Membrane Based on Polydopamine Coating: A Platform for Studying Immobilization Mechanisms. Langmuir 2018, 27, 2585–2594. [Google Scholar] [CrossRef] [PubMed]
- Lowe, A.B. Thiol–ene “click” reactions and recent applications in polymer and materials synthesis: A first update. Polym. Chem. 2014, 5, 4820–4870. [Google Scholar] [CrossRef]
- Breder, A.; Ortgies, S. Recent developments in sulfur- and selenium-catalyzed oxidative and isohypsic functionalization reactions of alkenes. Tetrahedron Lett. 2015, 56, 2843–2852. [Google Scholar] [CrossRef]
- Santra, S.; Bean, R.; Heckert, B.; Shaw, Z.; Jain, V.; Shrestha, L.; Narayanam, R.; Austin, Q. Alkene–azide chemistry: A facile, one-step, solvent- and catalyst-free approach for developing new functional monomers and polymers. Polym. Chem. 2020, 11, 3723. [Google Scholar] [CrossRef]
- Xi, W.; Scott, T.F.; Kloxin, C.J.; Bowman, C.N. Click Chemistry in Materials Science. Adv. Funct. Mater. 2014, 24, 2572–2590. [Google Scholar] [CrossRef]
- Smith, L.C.; Faustinella, F.; Chan, L. Lipases: Three-dimensional structure and mechanism of action. Curr. Opin. Struct. Biol. 1992, 2, 490–496. [Google Scholar] [CrossRef]
- Fernandez-Lorente, G.; Cabrera, Z.; Godoy, C.A.; Fernandez-Lafuente, R.; Palomo, J.M.; Guisan, J.M. Interfacially activated lipases against hydrophobic supports: Effect of the support nature on the biocatalytic properties. Process. Biochem. 2008, 43, 1061–1067. [Google Scholar] [CrossRef]
- Iqbal, K.F.; Dongming, L.; Rabia, D.; Weiqian, H.; Zexin, Z.; Yonghua, W. The Lid Domain in Lipases: Structural and Functional Determinant of Enzymatic Properties. Front. Bioeng. Biotechnol. 2017, 5, 16. [Google Scholar]
- Carrasco-López, C.; Godoy, C.; de las Rivas, B.; Fernandez-Lorente, G.; Palomo, J.M.; Guisán, J.M.; Fernández-Lafuente, R.; Martínez-Ripoll, M.; Hermoso, J.A. Activation of bacterial thermoalkalophilic lipases is spurred by dramatic structural rearrangements. J. Biol. Chem. 2009, 284, 4365–4372. [Google Scholar] [CrossRef] [PubMed]
- Romero, O.; Filice, M.; de las Rivas, B.; Carrasco-Lopez, C.; Klett, J.; Morreale, A.; Hermoso, J.A.; Guisan, J.M.; Abian, O.; Palomo, J.M. Semisynthetic peptide-lipase conjugates for improved biotransformations. Chem. Commun. 2012, 48, 9053–9055. [Google Scholar] [CrossRef]
- Godoy, C.A.; Romero, O.; de las Rivas, B.; Mateo, C.; Fernandez-Lorente, G.; Guisan, J.M.; Palomo, J.M. Changes on enantioselectivity of a genetically modified thermophilic lipase by site-directed oriented immobilization. J. Mol. Catal. B Enzym. 2013, 87, 121–127. [Google Scholar] [CrossRef]
- Palomo, J.M. Click reactions in protein chemistry: From the preparation of semisynthetic enzymes to new click enzymes. Org. Biomol. Chem. 2012, 10, 9309–9318. [Google Scholar] [CrossRef]
- Breugst, M.; Reissig, H.-U. The Huisgen Reaction: Milestones of the 1,3-Dipolar Cycloaddition. Angew. Chem. Int. Ed. 2020, 59, 12293–12307. [Google Scholar] [CrossRef]
- Jewetta, J.C.; Bertozzi, C.R. Cu-free click cycloaddition reactions in chemical biology. Chem. Soc. Rev. 2010, 39, 1272–1279. [Google Scholar] [CrossRef] [PubMed]
- Jain, Y.; Gupta, R.; Yadav, P.; Kumari, M. Chemical Waltz of Organic Molecules “On Water”: Saline-Assisted Sustainable Regioselective Synthesis of Fluorogenic Heterobioconjugates via Click Reaction. ACS Omega 2019, 4, 3582–3592. [Google Scholar] [CrossRef]
- Lopez-Tejedor, D.; Clavería-Gimeno, R.; Velazquez-Campoy, A.; Abian, O.; Palomo, J.M. Tyrosinase from mushroom Agaricus bisporus as an inhibitor of the Hepatitis C virus. bioRxiv 2020. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Entry | Protein 1 | Support | Mg Protein/g Support | Immobilization Yield (%) 2 | Retained Activity (%) 4 |
|---|---|---|---|---|---|
| 1 | CAL-B | Geranyl-SP | 9.0 | 96 | 149 |
| 2 | TLL | Geranyl-SP | 9.2 | 92 | 91 |
| 3 | RML | Geranyl-SP | 7.2 | 84 | 182 |
| 4 | ANL | Geranyl-SP | 8.8 | 70 | 105 |
| 5 | CRL | Geranyl-SP | 8.2 | 81 | 15 |
| 6 | PFL | Geranyl-SP | 8.4 | 71 | 20 |
| 7 | LECI | Geranyl-SP | 12 | 95 | 51 |
| 8 | Tyr | Geranyl-CM | 3 | 40 | nd |
| 9 | Tyr | Geranyl-SP | 3 | 90 | 50 |
| 10 | β-Gal/ Lac | Geranyl-SP | 2.12/11 | 0 | - |
| 11 | Lp_0440 | Geranyl-CM | 1.62 | >95 3 | nd |
| 12 | Lp_3525 | Geranyl-CM | 2.7 | 66 3 | nd |
| Protein 1 | Additive | pH | Mg Protein/g Support | Immobilization Yield (%) 4 | Retained Activity (%) 5 |
|---|---|---|---|---|---|
| GTLC193 | Triton X-100 2 | 5 | 10 | <10 | 90 |
| GTLC193 | Triton X-100 2 | 6 | 10 | <10 | 90 |
| GTLC193 | Triton X-100 2 | 8 | 10 | 42 | 84 |
| GTLC334 | Triton X-100 2 | 8 | 10 | 64 | 48 |
| RML_SH | Lauryl Sucrose 3 | 8 | 14 | 64 | 99 |
| B-Gal_SH | Triton X-100 2 | 8 | 0.8 | 79 | 99 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brabcova, J.; Andreu, A.; Aguilera, D.; Cabrera, Z.; de las Rivas, B.; Muñoz, R.; Palomo, J.M. Geranyl Functionalized Materials for Site-Specific Co-Immobilization of Proteins. Molecules 2021, 26, 3028. https://doi.org/10.3390/molecules26103028
Brabcova J, Andreu A, Aguilera D, Cabrera Z, de las Rivas B, Muñoz R, Palomo JM. Geranyl Functionalized Materials for Site-Specific Co-Immobilization of Proteins. Molecules. 2021; 26(10):3028. https://doi.org/10.3390/molecules26103028
Chicago/Turabian StyleBrabcova, Jana, Alicia Andreu, David Aguilera, Zaida Cabrera, Blanca de las Rivas, Rosario Muñoz, and Jose M. Palomo. 2021. "Geranyl Functionalized Materials for Site-Specific Co-Immobilization of Proteins" Molecules 26, no. 10: 3028. https://doi.org/10.3390/molecules26103028
APA StyleBrabcova, J., Andreu, A., Aguilera, D., Cabrera, Z., de las Rivas, B., Muñoz, R., & Palomo, J. M. (2021). Geranyl Functionalized Materials for Site-Specific Co-Immobilization of Proteins. Molecules, 26(10), 3028. https://doi.org/10.3390/molecules26103028