
α-Terpineol: An Aggregation Pheromone in Optatus palmaris (Coleoptera: Curculionidae) (Pascoe, 1889) Enhanced by Its Host-Plant Volatiles

 , ,
, ,  and
and
Abstract
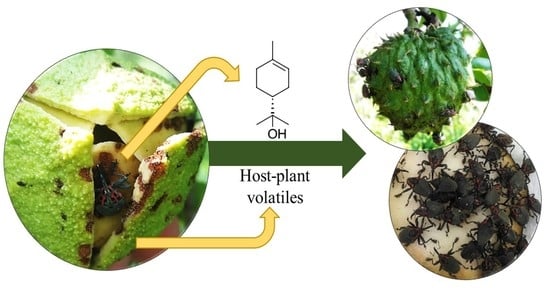
1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Insect Collection and Species Corroboration
4.2. Volatile Capture
4.2.1. Solid-Phase Microextraction
4.2.2. Dynamic Headspace
4.3. Gas Chromatography Coupled to Mass Spectrometry (GC-MS)
4.3.1. Matrix Recovering Effects
4.3.2. Limit of Detection (LOD) and Limit of Quantification (LOQ)
4.4. Bioassays
4.4.1. Reversal Feeding Experiments
4.4.2. Behavioral Response of Males and Female Weevils to Volatile Compounds
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Morrone, J.J. Biodiversity of Curculionoidea (Coleoptera) in Mexico. Rev. Mex. Biodivers. 2014, 85, 312–324. [Google Scholar] [CrossRef]
- Hespenheide, H.A. Beetles. In Encyclopedia of Biodiversity, 1st ed.; Levin, S.A., Ed.; Academic Press: Princeton, NJ, USA, 2001; Volume 1, pp. 351–358. [Google Scholar]
- Servín, V.R.; García, H.J.L.; Tejas, R.A.; Martínez, C.J.L.; Toapanta, M.A. Susceptibility of pepper weevil (Anthonomus eugenii Cano) (Coleoptera: Curculionidae) to seven insecticides in rural areas of Baja California Sur, Mexico. Acta Zool. Mex. 2008, 24, 45–54. [Google Scholar]
- Oehlschlager, A.C.; Chinchilla, C.; Castillo, G.; González, L. Control of red ring disease by mass trapping of Rhynchophorus palmarum (Coleoptera: Curculionidae). FLA Entomol. 2002, 85, 507–513. [Google Scholar] [CrossRef]
- Ocan, D.; Mukasa, H.H.; Rubaihayo, P.R.; Tinzaara, W.; Blomme, G. Effects of banana weevil damage on plant growth and yield of East African Musa genotypes. J. Appl. Biol. Sci. 2008, 9, 407–415. [Google Scholar]
- Hernández, F.L.M.; Castañeda, V.A.; Urías, L.M.A. Weevil borers in tropical fruit crops: Importance, biology and management. In Insect Physiology and Ecology, 2nd ed.; Shields, V.D.C., Ed.; IntechOpen: Rijeka, Croatia, 2017; Volume 3, pp. 154–196. [Google Scholar]
- Showler, A.T. Boll weevil (Coleoptera: Curculionidae) damage to cotton bolls under standard and proactive spraying. J. Econ. Entomol. 2006, 99, 1251–1257. [Google Scholar] [CrossRef] [PubMed]
- Esparza, D.G.; Olguín, A.; Carta, L.K.; Skantar, A.M.; Villanueva, R.T. Detection of Rhynchonphorus palmarum (Coleoptera:Curculionidae) and identification of associated nematodes in South Texas. FLA Entomol. 2013, 96, 1513–1521. [Google Scholar] [CrossRef]
- Azuara, D.A.; Terán, V.A.P.; Soto, S.A.; Aguilar, P.N.; Martínez, B.L. Evaluación del tipo de trampa, atrayente alimenticio y feromona de agregación en el trampeo del picudo del agave Scyphophorus acupunctatus Gyllenhal en Tamaulipas, México. Entomotropica 2014, 29, 1–8. [Google Scholar]
- Bautista, H.C.F.; Cibrián, T.J.; Velázquez, G.J.C.; Rodríguez, G.M.P. Evaluación en campo de atrayentes para la captura de Anthonomus eugenii Cano (Coleoptera: Curculionidae). Rev. Chil. Entomol. 2020, 46, 211–219. [Google Scholar]
- Coelho, L.M.A.; Alves, R.E. Soursop (Annona muricata L.). In Postharvest Biology and Technology of Tropical and Subtropical Fruits. Mangosteen to White Sapote; Yahia, M.E., Ed.; Woodhead Publishing Limited: Cambridge, UK, 2011; Volume 4, pp. 363–391. [Google Scholar]
- Jiménez, Z.J.O.; Balois, M.R.; Alia, T.I.; Juárez, L.P.; Sumaya, M.M.T.; Bello, L.J.E. Characterization of soursop fruit (Annona muricata L.) in Tepic, Nayarit, Mexico. Rev. Mex. Cienc. Agríc. 2016, 7, 1261–1270. [Google Scholar]
- Corrales, M.G.J. Observations on the biology and behavioral food of Optatus sp. (Coleoptera: Curculionidae) in Annona cherimola Mill. In Resúmenes de 2do. Congreso Centroamericano y del Caribe y 3ero. Costarricense de Entomología; Editorial San José ASENCO: San José, Costa Rica, 1995; p. 6. [Google Scholar]
- Hernández-Fuentes, L.M.; Gómez, J.R.; Andrés, A.J. Importance, pest insect and diseases fungus of the cultivation of the soursoup. In Libro Técnico Núm. 1, 1st ed.; Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, Campo Experimental Santiago Ixcuintla: Nayarit, México, 2013; Volume 1, pp. 85–87. [Google Scholar]
- Maldonado, E.; Hernández, L.M.; Luna, G.; Gómez, J.R.; Flores, R.J.; Orozco-Santos, M. Bioecology of Optatus palmaris Pascoe (Coleoptera:Curculionidae) in Annona muricata L. Southw. Entomol. 2014, 39, 773–782. [Google Scholar] [CrossRef]
- Castañeda-Vildózola, A.; Nava-Díaz, C.; Hernández-Fuentes, L.M.; Valdez-Carrasco, J.; Colunga-Treviño, B. New host record and geographical distribution of Optatus palmaris Pascoe (Coleoptera: Curculionidae) in Mexico. Acta Zool. Mex. 2009, 25, 663–666. [Google Scholar]
- Castañeda-Vildózola, A.; Morales-Trujillo, M.C.; Franco-Mora, O.; Valdez-Carrasco, J.; Mejía-Carranza, J. A new record of Optatus palmaris Pascoe (Coleoptera: Curculionidae) associated with Annona L. (Annonaceae) in the State of Mexico, Mexico. Revista Chilena de Entomología 2020, 46, 397–400. [Google Scholar] [CrossRef]
- Hernández, F.L.M.; Nolasco, G.Y.; Orozco, S.M.; Montalvo, G.E. Toxicidad de insecticidas contra (Optatus palmaris Pascoe) en guanábana. Rev. Mex. Cienc. Agríc. 2021, 12, 49–60. [Google Scholar]
- Tewari, S.; Leskey, T.C.; Nielsen, A.L.; Piñero, J.C.; Rodriguez, S.C.R. Integrated pest management: Current concepts and ecological perspective. In Integrated Pest Management: Current Concepts and Ecological Perspective, 1st ed.; Abrol, D.P., Ed.; Academic Press: San Diego, CA, USA, 2014; pp. 141–168. [Google Scholar]
- Mormann, M.; Salpin, J.Y.; Kuck, D. Gas-phase titration of C7H9+ ion mixtures by FT-ICR mass spectrometry: Semiquantitative determination of ion populations generated by CI-induced protonation of C7H8 isomers and by EI-induced fragmentation of some monoterpenes. Int. J. Mass Spect. 2006, 249–250, 340–352. [Google Scholar] [CrossRef]
- Nicolescu, T.O. Interpretation of mass spectra. In Mass Spectrometry; Aliofkhazraei, M., Ed.; Intech Open: London, UK, 2017; pp. 24–78. [Google Scholar]
- Beck, J.J.; Torto, B.; Vannette, R.L. Eavesdropping on plant-insect-microbe chemical communications in agricultural ecology: A virtual issue on semiochemicals. J. Agric. Food Chem. 2017, 65, 5101–5103. [Google Scholar] [CrossRef] [PubMed]
- Beck, J.J.; Vannette, R.L. Harnessing insect-microbe chemical communications to control insect pests of agricultural systems. J. Agric. Food Chem. 2017, 65, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Nandagopal, V.; Prakash, A.; Rao, J. Know the pheromones: Basics and its application. J. Biopestic. 2008, 1, 210–215. [Google Scholar]
- Sharma, A.; Sandhi, R.K.; Reddy, G.V.P. A Review of interactions between insect biological control agents and semiochemicals. Insects 2019, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Haenniger, S.; Goergen, G.; Akinbuluma, M.D.; Kunert, M.; Heckel, D.G.; Unbehend, M. Sexual communication of Spodoptera frugiperda from West Africa: Adaptation of an invasive species and implications for pest management. Sci. Rep. 2020, 10, 2892. [Google Scholar] [CrossRef] [PubMed]
- Bruce, T.J.A.; Wadhams, L.J.; Woodcock, C.M. Insect host location: A volatile situation. Trends Plant Sci. 2005, 10, 269–274. [Google Scholar] [CrossRef]
- Wang, H.M.; Bai, P.H.; Zhang, J.; Zhang, X.M.; Hui, Q.; Zheng, H.X.; Zhang, X.H. Attraction of bruchid beetles Callosobruchus chinensis (L.) (Coleoptera: Bruchidae) to host plant volatiles. J. Integr. Agric. 2020, 19, 3035–3044. [Google Scholar] [CrossRef]
- Webster, B.; Cardé, R. Use of habitat odour by host-seeking insects. Biol. Rev. 2016, 92, 1241–1249. [Google Scholar] [CrossRef]
- Erb, M.; Robert, C.A.M. Sequestration of plant secondary metabolites by insect herbivores: Molecular mechanisms and ecological consequences. Curr. Opin. Insect. Sci. 2016, 14, 8–11. [Google Scholar] [CrossRef]
- Beran, F.; Köllner, T.G.; Gershenzon, J.; Tholl, D. Chemical convergence between plants and insects: Biosynthetic origins and functions of common secondary metabolites. New Phytol. 2019, 223, 52–67. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Lu, L.H.; Shi, Q.X.; Chen, J.; He, Y.R. Volatile terpenes and terpenoids from workers and queens of Monomorium chinense (Hymenoptera: Formicidae). Molecules 2018, 23, 2838. [Google Scholar] [CrossRef]
- Eller, F.J.; Bartelt, R.J.; Shasha, B.S.; Schuster, D.J.; Riley, D.J.; Stansly, P.A.; Mueller, T.F.; Shuler, K.D.; Johnson, B.; Davis, J.H.; et al. Aggregation pheromone for the pepper weevil, Anthonomus eugenii Cano (Coleoptera: Curculionidae): Identification and field activity. J. Chem. Ecol. 1994, 20, 1537–1555. [Google Scholar] [CrossRef] [PubMed]
- Cross, J.V.; Hall, D.R.; Innocenzi, P.J.; Hesketh, H.; Jay, C.N.; Burgess, C.M. Exploiting the aggregation pheromone of strawberry blossom weevil Anthonomus rubi (Coleoptera: Curculionidae): Part 2. Pest monitoring and control. Crop Prot. 2006, 25, 155–166. [Google Scholar] [CrossRef]
- Hardee, D.D.; McKibben, G.H.; Rummel, D.R.; Huddleston, P.M.; Coppege, J.R. Response of boll weevils to component ratios and doses of the Grandlure. Environ. Entomol. 1974, 3, 135–138. [Google Scholar] [CrossRef]
- Hedin, P.A.; Dollar, D.A.; Collins, J.K.; Dubois, J.G.; Mulder, P.G.; Hedger, G.H.; Smith, M.W.; Eikenbary, R.D. Identification of male pecan weevil pheromone. J. Chem. Ecol. 1997, 23, 965–977. [Google Scholar] [CrossRef]
- Lacey, E.S.; Moreira, J.A.; Millar, J.G.; Hanks, L.M. A Male-produced aggregation pheromone blend consisting of alkanediols, terpenoids, and an aromatic alcohol from the Cerambycid Beetle Megacyllene caryae. J. Chem. Ecol. 2008, 34, 408–417. [Google Scholar] [CrossRef]
- Mitchell, R.F.; Ray, A.M.; Hanks, L.M.; Millar, J.G. The common natural products (s)-α-terpineol and (e)-2-hexenol are important pheromone components of Megacyllene antennata (Coleoptera: Cerambycidae). Environ. Entomol. 2018, 47, 1547–1552. [Google Scholar] [CrossRef] [PubMed]
- Tinzaara, W.; Dicke, M.; Van, H.A.; Gold, C.S. Use of infochemicals in pest management with special reference to the banana weevil, Cosmopolites sordidus (Germar) (Coleoptera: Curculionidae). Int. J. Trop. Insect Sci. 2002, 22, 241–261. [Google Scholar] [CrossRef]
- Müller, M.; Buchbauer, G. Essential oil components as pheromones. A review. Flavour Fragr. J. 2011, 26, 357–377. [Google Scholar] [CrossRef]
- Stelinski, L.L.; Oakleaf, R.; Rodríguez, S.C. Oviposition deterring pheromone deposite don blueberry fruit by the parasitic wasp, Diachasma alloeum. Behavior 2007, 144, 429–445. [Google Scholar]
- Landolt, P.J.; Phillips, T.W. Host plant influences on sex pheromone behavior of phytophagous insects. Annu. Rev. Entomol. 1997, 42, 371–391. [Google Scholar] [CrossRef]
- Dickens, J.C. Green leaf volatiles enhance aggregation pheromone of boll weevil, Anthonomus grandis. Entomol. Exp. Appl. 1989, 52, 191–203. [Google Scholar] [CrossRef]
- Piñero, J.C.; Prokopy, R.J. Field evaluation of plant odor and pheromonal combinations for attracting plum curculios. J. Chem. Ecol. 2003, 29, 2735–2748. [Google Scholar] [CrossRef]
- Piñero, J.C.; Wright, S.E.; Prokopy, R.J. Response of plum curculio (Coleoptera: Curculionidae) to odor-baited traps near woods. J. Econ. Entomol. 2001, 94, 1386–1397. [Google Scholar] [CrossRef]
- Hallett, R.H.; Oehlschlager, A.C.; Borden, J.H. Pheromone trapping protocols for the Asian palm weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae). Int. J. Pest Manag. 1999, 45, 231–237. [Google Scholar] [CrossRef]
- Cruz, F.J.J.; Figueroa, C.P.; Alcántara, J.J.A.; López, M.V.; Silva, G.F. Vegetal synergists for trapping the adult of Scyphophorus acupunctatus Gyllenhal, in pheromone baited traps, in Agave angustifolia Haw, in Morelos, México. Acta Zool. Mex. 2019, 35, 1–9. [Google Scholar] [CrossRef]
- Borden, J.H.; Pureswaran, D.S.; Lafontaine, J.P. Synergistic blends of monoterpenes for aggregation pheromones of the mountain pine beetle (Coleoptera: Curculionidae). J. Econ. Entomol. 2008, 101, 1266–1275. [Google Scholar] [CrossRef]
- Erasto, P.; Viljoen, A.M. Limonene—A Review: Biosynthetic, Ecological and Pharmacological Relevance. Nat. Prod. Commun. 2008, 3, 1193–1202. [Google Scholar] [CrossRef]
- Hartsel, J.A.; Eades, J.; Hickory, B.; Makriyannis, A. Cannabis sativa and Hemp. In Nutraceuticals: Efficacy, Safety and Toxicity, 1st ed.; Gupta, R., Ed.; Academic Press: Princeton, NJ, USA, 2016; Volume 1, pp. 735–754. [Google Scholar]
- Champion, G.C. Biologia Centrali-Americana. Insecta. Coleoptera, Rhynchophora Part 5; Curculionidae. Curculioninae: Porter, London, 1907; Volume 4, p. 513. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| One Odor Source Trials | |
|---|---|
| Air | Soursop |
| Air | Males |
| Air | Females |
| Air | α-terpineol |
| Air | soursop + α-terpineol |
| Air | soursop + males |
| Air | soursop + females |
| Two Odor Source Trials | |
| Soursop | soursop + males |
| Soursop | soursop + females |
| Soursop | soursop + α-terpineol |
| soursop + females | soursop + males |
| Females | Males |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pineda-Ríos, J.M.; Cibrián-Tovar, J.; Hernández-Fuentes, L.M.; López-Romero, R.M.; Soto-Rojas, L.; Romero-Nápoles, J.; Llanderal-Cázares, C.; Salomé-Abarca, L.F. α-Terpineol: An Aggregation Pheromone in Optatus palmaris (Coleoptera: Curculionidae) (Pascoe, 1889) Enhanced by Its Host-Plant Volatiles. Molecules 2021, 26, 2861. https://doi.org/10.3390/molecules26102861
Pineda-Ríos JM, Cibrián-Tovar J, Hernández-Fuentes LM, López-Romero RM, Soto-Rojas L, Romero-Nápoles J, Llanderal-Cázares C, Salomé-Abarca LF. α-Terpineol: An Aggregation Pheromone in Optatus palmaris (Coleoptera: Curculionidae) (Pascoe, 1889) Enhanced by Its Host-Plant Volatiles. Molecules. 2021; 26(10):2861. https://doi.org/10.3390/molecules26102861
Chicago/Turabian StylePineda-Ríos, José Manuel, Juan Cibrián-Tovar, Luis Martín Hernández-Fuentes, Rosa María López-Romero, Lauro Soto-Rojas, Jesús Romero-Nápoles, Celina Llanderal-Cázares, and Luis F. Salomé-Abarca. 2021. "α-Terpineol: An Aggregation Pheromone in Optatus palmaris (Coleoptera: Curculionidae) (Pascoe, 1889) Enhanced by Its Host-Plant Volatiles" Molecules 26, no. 10: 2861. https://doi.org/10.3390/molecules26102861
APA StylePineda-Ríos, J. M., Cibrián-Tovar, J., Hernández-Fuentes, L. M., López-Romero, R. M., Soto-Rojas, L., Romero-Nápoles, J., Llanderal-Cázares, C., & Salomé-Abarca, L. F. (2021). α-Terpineol: An Aggregation Pheromone in Optatus palmaris (Coleoptera: Curculionidae) (Pascoe, 1889) Enhanced by Its Host-Plant Volatiles. Molecules, 26(10), 2861. https://doi.org/10.3390/molecules26102861