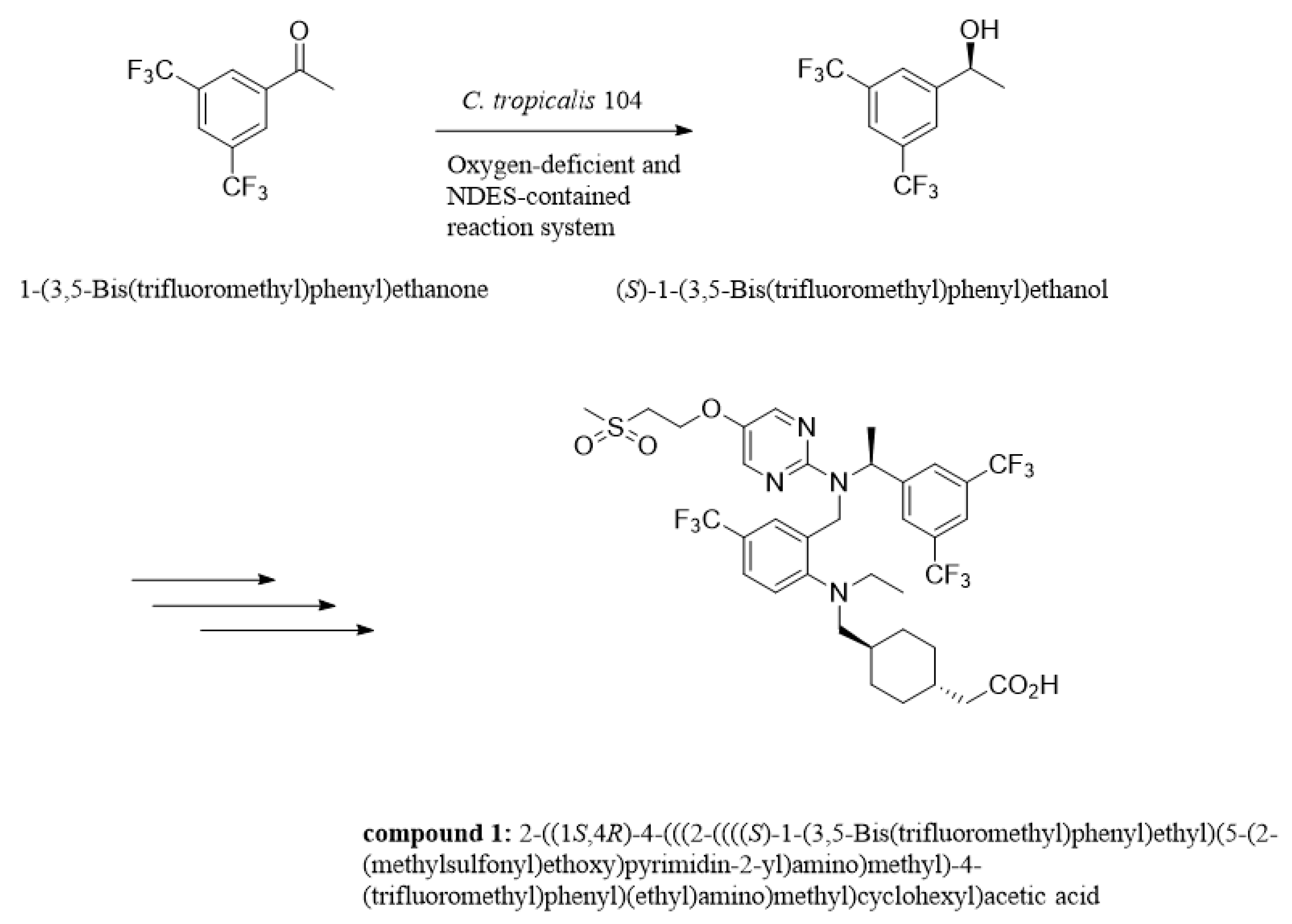
High-Efficient Production of (S)-1-[3,5-Bis(trifluoromethyl)phenyl]ethanol via Whole-Cell Catalyst in Deep-Eutectic Solvent-Containing Micro-Aerobic Medium System


Abstract
1. Introduction
2. Results and Discussion
2.1. Oxygen Control Means Selecting and Effect on (S)-3,5-BTPE Production by C. tropicalis 104
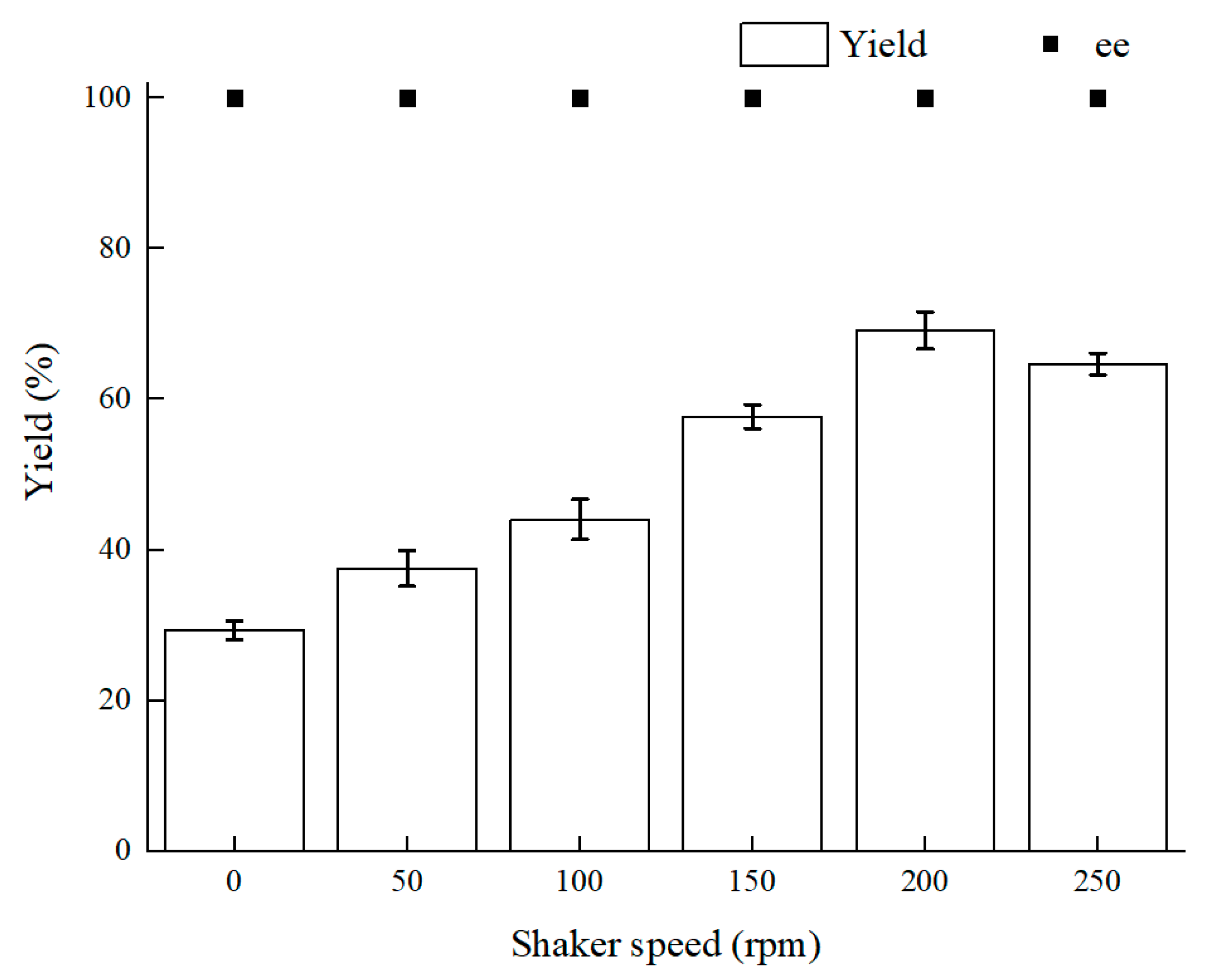
2.1.1. Developing an Oxygen Control Proposal for (S)-3,5-BTPE Production by C. tropicalis 104
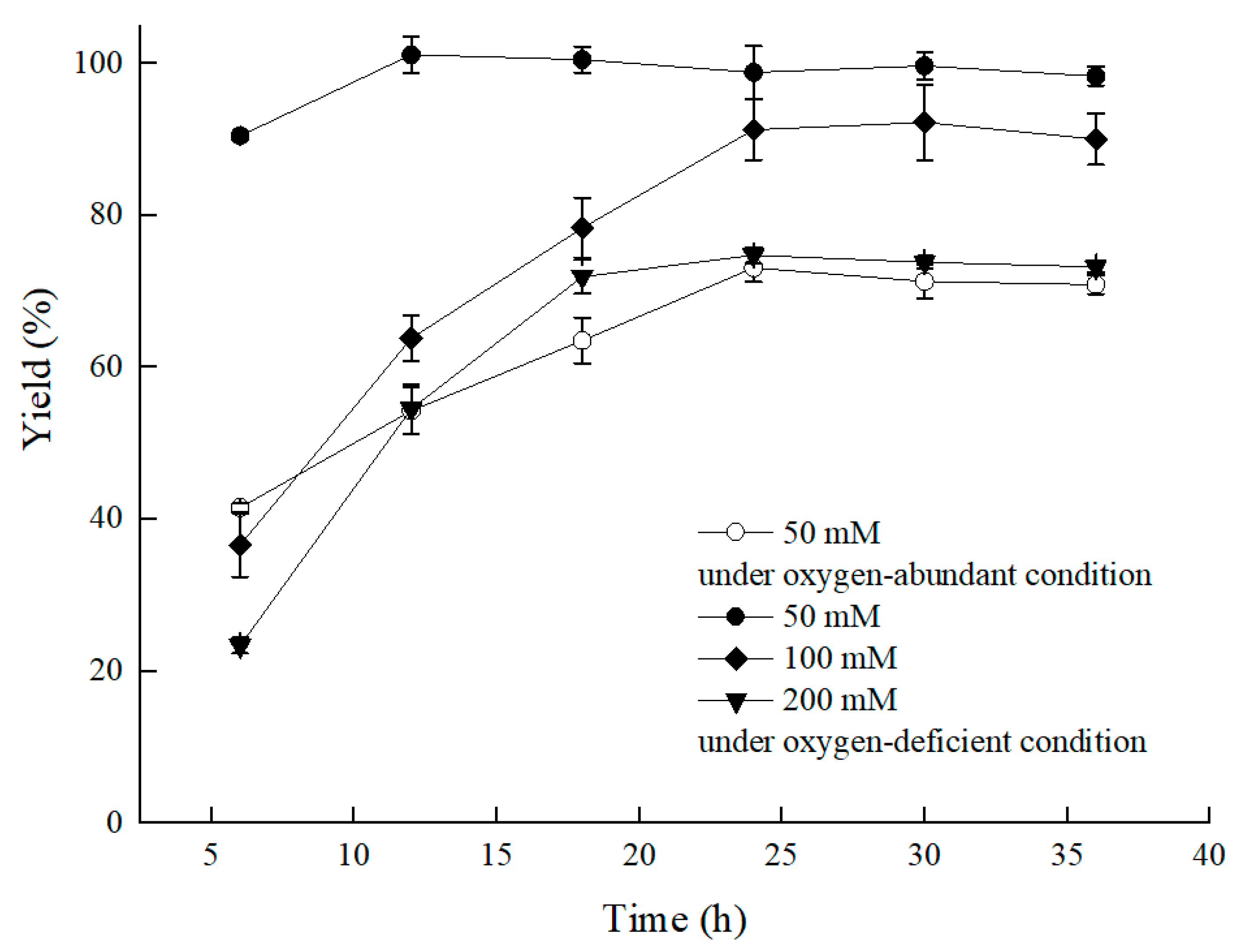
2.1.2. Effect of Limited Oxygen on C. tropicalis 104 in Bioprocess
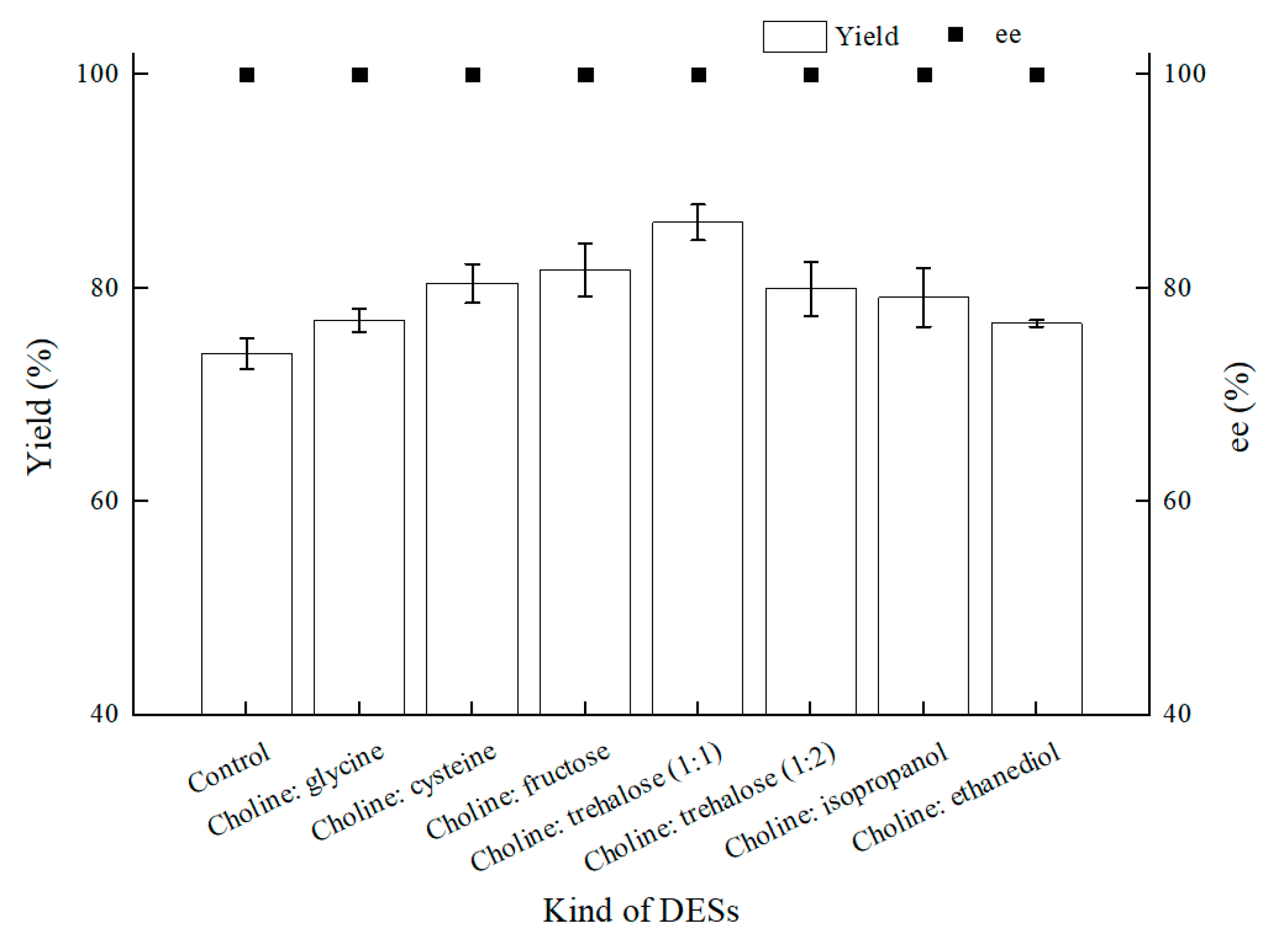
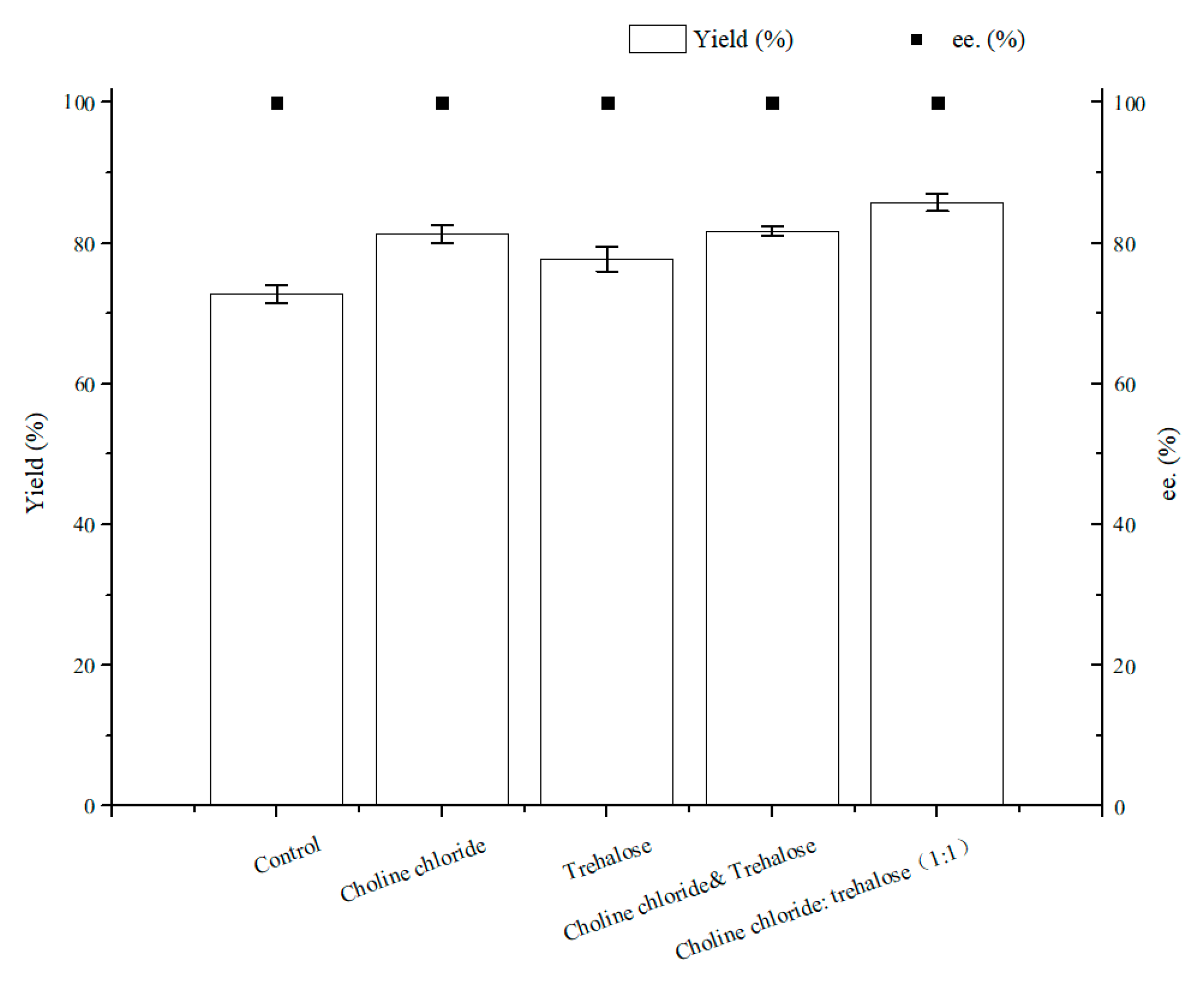
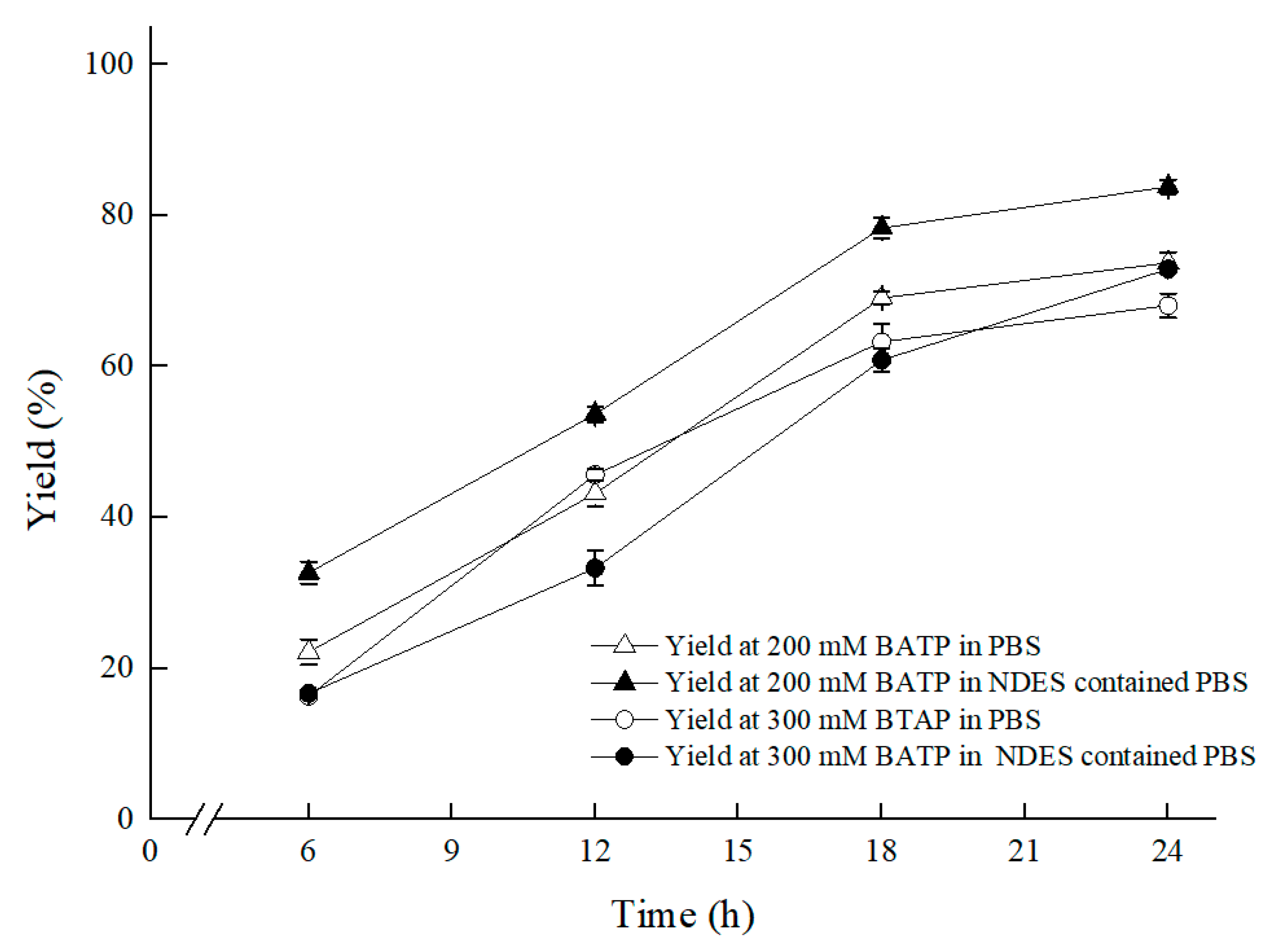
2.2. NADESs Selection and Effect on (S)-3,5-BTPE Production by C. tropicalis 104
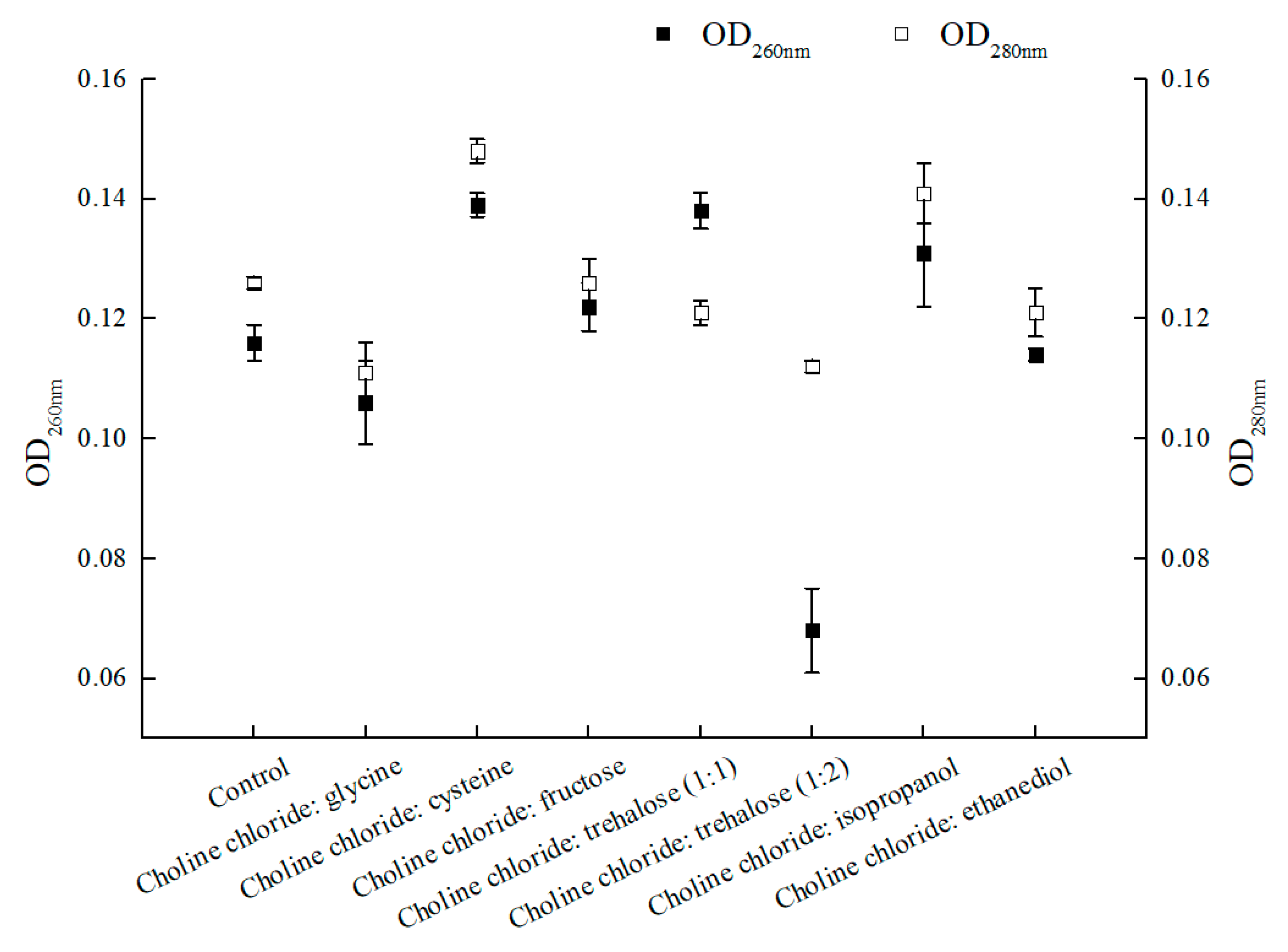
2.2.1. Effect of Various NADESs on Membrane Permeability of C. tropicalis 104
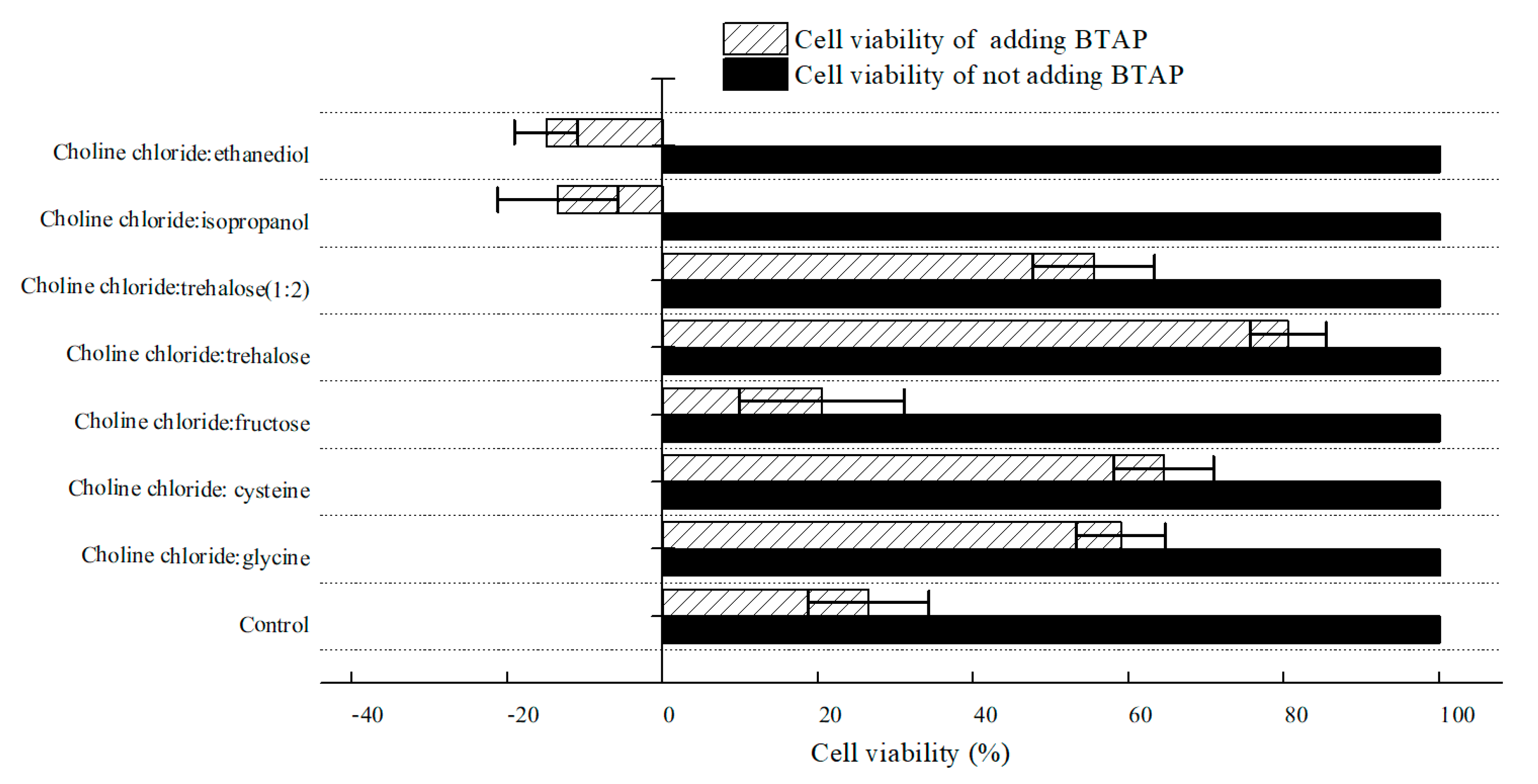
2.2.2. Effect of Various NADESs on Cell Viability of C. tropicalis 104
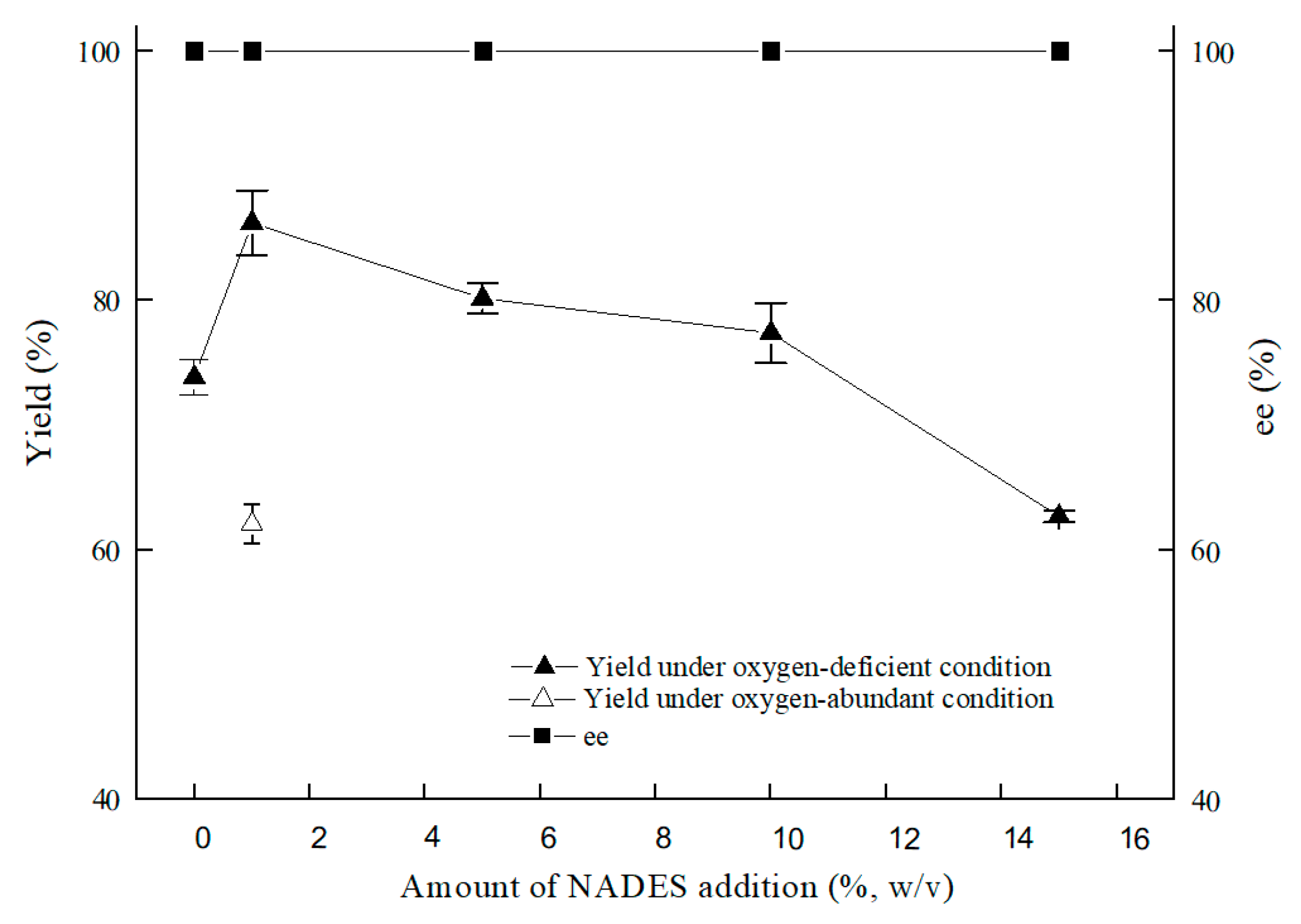
2.2.3. Effect of NADESs on (S)-3,5-BTPE Production with C. tropicalis 104 under Oxygen-Deficient Environment
3. Materials and Methods
3.1. Materials
3.2. Methods
3.2.1. Microorganism and Cultivation
3.2.2. General Procedure for Asymmetric Bio-Reduction of BTAP to (S)-3,5-BTPE
3.2.3. Investigation of Cofactor Specificity
3.2.4. Assays of Intracellular NADH Concentration
3.2.5. Assays of Membrane Permeability
3.2.6. Assays of Cell Viability
3.2.7. GC Analysis for (S)-3,5-BTPE and BTAP
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Fabián, G.P.; Liliana, B.F.; José, H.A.; Diego, G.S. Whole cells as biocatalysts in organic transformations. Molecules 2018, 23, 1265. [Google Scholar] [CrossRef]
- Liu, Z.-Q.; Yin, H.-H.; Zhang, X.-J.; Zhou, R.; Wang, Y.-M.; Zheng, Y.-G. Improvement of carbonyl reductase activity for the bioproduction of tert-butyl (3R,5S)-6-chloro-3,5-dihydroxyhexanoate. Bioorganic Chem. 2018, 80, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Koto, H.; Shinjiro, T.; Tatsuya, T. Asymmetric aerobic oxidation of secondary alcohols catalyzed by poly(N-vinyl-2-pyrrolidone)-stabilized gold clusters modified with cyclodextrin derivatives. Chem. Commun. 2019, 55, 15033. [Google Scholar] [CrossRef]
- Christopher, S.; Nadia, M.J.R. Progress in the development of neurokinin antagonists. Annu. Rep. Med. Chem. 1999, 34, 51–60. [Google Scholar] [CrossRef]
- Böhm, H.-J.; Banner, D.; Bendels, S.; Kansy, M.; Kuhn, B.; Müller, K.; Obst-Sander, U.; Stahl, M. Fluorine in Medicinal Chemistry. ChemBioChem 2004, 5, 637–643. [Google Scholar] [CrossRef]
- Murakami, T.; Murakami, K.; Ogiya, T.; Shibuya, K. Agent for Inhibiting Expression of NFKBIL1, ITGAZ, FCER1A, PHACTR1, and/or IRF5 mRNA, and Agent for Preventing and/or Treating Immune Disease. Patent Number JP 2013112674, 29 May 2013. [Google Scholar]
- Tadaaki, O.; Takaeshi, M.; Katsutoshi, M.; Koichi, Y.; Taichi, K. Optically Active Dibenzylamine Derivative, and Method for Preparing Thereof. U.S. Patent 201,302,258,14A, 29 August 2013. [Google Scholar]
- Szőllősi, G.; Kolcsár, V.J. Highly Enantioselective Transfer Hydrogenation of Prochiral Ketones Using Ru(II)-Chitosan Catalyst in Aqueous Media. ChemCatChem 2018, 11, 820–830. [Google Scholar] [CrossRef]
- Yu, S.; Ma, S. Allenes in Catalytic Asymmetric Synthesis and Natural Product Syntheses. Angew. Chem. Int. Ed. 2012, 51, 3074–3112. [Google Scholar] [CrossRef]
- Diya, A.; Safaa, A.; Francesca, P. Extreme halophilic alcohol dehydrogenase mediated highly efficient syntheses of enantiopure aromatic alcohols. Org. Biomol. Chem. 2017, 15, 9169–9175. [Google Scholar] [CrossRef]
- Pollard, D.; Truppo, M.; Pollard, J.; Chen, C.-Y.; Moore, J. Effective synthesis of (S)-3,5-bistrifluoromethylphenyl ethanol by asymmetric enzymatic reduction. Tetrahedron Asymmetry 2006, 17, 554–559. [Google Scholar] [CrossRef]
- Wang, P.; Su, H.; Sun, L.; He, J.; Lü, Y. Asymmetric Bioreduction of 3,5-Bis(trifluoromethyl) Acetophenone to Its Corresponding Alcohol by Candida tropicalis. Chin. J. Chem. Eng. 2011, 19, 1028–1032. [Google Scholar] [CrossRef]
- Dai, J.-J.; Cheng, J.-S.; Liang, Y.-Q.; Jiang, T.; Yuan, Y.-J. Regulation of extracellular oxidoreduction potential enhanced (R,R)-2,3-butanediol production by Paenibacillus polymyxa CJX518. Bioresour. Technol. 2014, 167, 433–440. [Google Scholar] [CrossRef]
- Benedikt, H.; Robin, L.; Hannah, T.; Lars, R.; Jochen, B. Shake flask methodology for assessing the influence of the maximum oxygen transfer capacity on 2,3-butanediol production. Microb. Cell Fact. 2019, 18, 78. [Google Scholar] [CrossRef]
- Chen, H.J.; Qian, F.; Lin, H.; Wang, P.; Chen, H.; Chen, W. Using Choline Chloride-Based DESs as Co-Solvent for 3,5-Bis(trifluoromethyl) Acetophenone Bioreduction with Rhodococcus erythropolis XS1012. Catalysts 2019, 10, 30. [Google Scholar] [CrossRef]
- Hui, L.; Bi-Shuang, C.; Fayene, D.S.; Lan, L. A comparative study on asymmetric reduction of ketones using the growing and resting cells of marine-derived fungi. Mar. Drugs 2018, 16, 62. [Google Scholar] [CrossRef]
- Wu, T.-T.; Zhao, X.-J.; Yang, R.-L.; Bilal, M.; Wang, Z.-Y.; Luo, H.-Z.; Xu, N.-N.; Nie, Z.-K. Catalytic Performance of a Robust Whole-Cell Biocatalyst in the Regioselective Synthesis of Helicid Esters Under Optimized Processing Conditions. Catal. Lett. 2020, 1–8. [Google Scholar] [CrossRef]
- Zhao, Q.; Ansorge-Schumacher, M.B.; Haag, R.; Wu, C. Living whole-cell catalysis in compartmentalized emulsion. Bioresour. Technol. 2019, 295, 122221. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.-P.; Wang, H.-B.; Wu, Z.-L. Efficient bioreductive production of (S)-N-Boc-3-hydroxypiperidine using ketoreductase ChKRED03. Process. Biochem. 2016, 51, 881–885. [Google Scholar] [CrossRef]
- Klibanov, A.M. Improving enzymes by using them in organic solvents. Nature 2001, 409, 241–246. [Google Scholar] [CrossRef]
- Xiao, Z.-J.; Du, P.-X.; Lou, W.-Y.; Wu, H.; Zong, M.-H. Using water-miscible ionic liquids to improve the biocatalytic anti-Prelog asymmetric reduction of prochiral ketones with whole cells of Acetobacter sp. CCTCC M209061. Chem. Eng. Sci. 2012, 84, 695–705. [Google Scholar] [CrossRef]
- Luo, W.; Deng, X.-X.; Huo, J.; Ruan, T.; Gong, Z.; Yan, J.-B.; Yang, Z.-H.; Quan, C.; Cui, Z.-F. Strengthening NADPH Regeneration for Improving Photo-biocatalytic Ketones Asymmetric Reduction Reaction by Synechocystis Through Overexpression of FNR. Catal. Lett. 2018, 148, 1714–1722. [Google Scholar] [CrossRef]
- Yang, X.; Jiang, L.; Jia, Y.; Hu, Y.; Xu, Q.; Xu, X.; Huang, H. Counteraction of Trehalose on N, N-Dimethylformamide-Induced Candida rugosa Lipase Denaturation: Spectroscopic Insight and Molecular Dynamic Simulation. PLoS ONE 2016, 11, e0152275. [Google Scholar] [CrossRef] [PubMed]
- Lukito, B.R.; Wu, S.; Saw, H.J.J.; Li, Z. One-Pot Production of Natural 2-Phenylethanol from L -Phenylalanine via Cascade Biotransformations. ChemCatChem 2019, 11, 831–840. [Google Scholar] [CrossRef]
- Hussain, W.; Pollard, D.J.; Lye, G.J. The bioreduction of a beta-tetralone to its corresponding alcohol by the yeast Trichosporon capitatum MY1890 and bacterium Rhodococcus erythropolis MA7213 in a range of ionic liquids. Biocatal. Biotransfor. 2007, 25, 443–452. [Google Scholar] [CrossRef]
- Liu, J.; Li, H.; Zhao, G.; Caiyin, Q.; Qiao, J. Redox cofactor engineering in industrial microorganisms: Strategies, recent applications and future directions. J. Ind. Microbiol. Biotechnol. 2018, 45, 313–327. [Google Scholar] [CrossRef]
- Annesha, S.; Avinash, V.S.; Sujata, V.S.; Pramod, P.W. The effect of CO2 in enhancing photosynthetic cofactor recycling for alcohol dehydrogenase mediated chiral synthesis in cyanobacteria. J. Biotechnol. 2019, 289, 1–6. [Google Scholar] [CrossRef]
- Zhang, F.; Zhu, C.-T.; Peng, Q.-M.; Wang, F.-Q.; Sheng, S.; Wu, Q.-Y.; Wang, J. Enhanced permeability of recombinant E. coli cells with deep eutectic solvent for transformation of rutin. J. Chem. Technol. Biotechnol. 2019, 95, 384–393. [Google Scholar] [CrossRef]
- Vicente, G.F.; Caroline, E.P. Deep eutectic solvents for redox biocatalysis. J. Biotechnol. 2019, 293, 24–35. [Google Scholar] [CrossRef]
- Mai, N.L.; Koo, Y.-M. Whole-Cell Biocatalysis in ionic liquids. In Advances in Biochemical Engineering/Biotechnology; Springer: Cham, Switzerland, 2018; Volume 168, pp. 105–132. [Google Scholar] [CrossRef]
- Li, J.; Wang, P.; He, Y.-S.; Zhu, Z.-R.; Huang, J. Toward Designing a Novel Oligopeptide-Based Deep Eutectic Solvent: Applied in Biocatalytic Reduction. ACS Sustain. Chem. Eng. 2018, 7, 1318–1326. [Google Scholar] [CrossRef]
- Priyanka, G.; Ismadi, M.-Z.; Paul, J.V.; Andreas, F.; Sameer, J.; Jayesh, B.; Kerry, H. Optimization of agitation speed in spinner flask for microcarrier structural integrity and expansion of induced pluripotent stem cells. Cytotechnology 2016, 68, 45–59. [Google Scholar] [CrossRef]
- Jennifer, S.S.; Nikolaos, P.; Edmund, M. Carbonyl reductase sniffer from the model organism daphnia: Cloning, substrate determination and inhibitory sensitivity. Chem. Bio. Interact. 2019, 307, 29–36. [Google Scholar] [CrossRef]
- Zhang, Y.; Huang, Z.; Du, C.; Li, Y.; Cao, Z. Introduction of an NADH regeneration system into Klebsiella oxytoca leads to an enhanced oxidative and reductive metabolism of glycerol. Metab. Eng. 2009, 11, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Cai, J.-B.; Ouyang, Q.; He, J.-Y.; Su, H.-Z. Asymmetric biocatalytic reduction of 3,5-bis(trifluoromethyl) acetophenone to (1R)-[3,5-bis(trifluoromethyl)phenyl] ethanol using whole cells of newly isolated Leifsonia xyli HS0904. Appl. Microbiol. Biotechnol. 2011, 90, 1897–1904. [Google Scholar] [CrossRef]
- Li, J.; Wang, P.; He, J.-Y.; Huang, J.; Tang, J. Efficient biocatalytic synthesis of (R)-[3,5-bis(trifluoromethyl)phenyl] ethanol by a newly isolated Trichoderma asperellum ZJPH0810 using dual cosubstrate: Ethanol and glycerol. Appl. Microbiol. Biotechnol. 2013, 97, 6685–6692. [Google Scholar] [CrossRef] [PubMed]
- Chou, H.-H.; Marx, C.J.; Sauer, U. Transhydrogenase Promotes the Robustness and Evolvability of E. coli Deficient in NADPH Production. PLoS Genet. 2015, 11, e1005007. [Google Scholar] [CrossRef] [PubMed]
- Vemuri, G.N.; Eiteman, M.A.; McEwen, J.E.; Olsson, L.; Nielsen, J. Increasing NADH oxidation reduces overflow metabolism in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2007, 104, 2402–2407. [Google Scholar] [CrossRef]
- Li, B.; Liu, Y.; Wang, L.; Hong, J.; Chen, Y.; Ying, H. RNA accumulation in Candida tropicalis based on cofactor engineering. FEMS Yeast Res. 2019, 19, 028. [Google Scholar] [CrossRef]
- Christer, L.; Inga-Lill, P.; Ricky, A.; Michel, R.; Lennart, A.; Lena, G. The importance of the glycerol 3-phosphate shuttle during aerobic growth of Saccharomyces cerevisiae. Yeast 1998, 14, 347–357. [Google Scholar] [CrossRef]
- He, Y.-C.; Zhang, D.-P.; Tao, Z.-C.; Lu, Y.; Ding, Y.; Liu, F.; Zhu, Z.-Z.; Rui, H.; Zheng, G.-W.; Zhang, X. Improved biosynthesis of ethyl (S)-4-chloro-3-hydroxybutanoate by adding l-glutamine plus glycine instead of NAD+ in β-cyclodextrin–water system. Bioresour. Technol. 2015, 182, 98–102. [Google Scholar] [CrossRef]
- Ou, X.-Y.; Wu, X.-L.; Peng, F.; Xu, P.; Zhang, S.-Y.; Zong, M.-H.; Lou, W.-Y. Highly efficient asymmetric reduction of 2-octanone in biphasic system by immobilized Acetobacter sp. CCTCC M209061 cells. J. Biotechnol. 2019, 299, 37–43. [Google Scholar] [CrossRef]
- Weinberg, J.M.; Bienholz, A.; Venkatachalam, M.A. The role of glycine in regulated cell death. Cell. Mol. Life Sci. 2016, 73, 2285–2308. [Google Scholar] [CrossRef]
- Chen, J.; Liu, X.; Mandel, L.J.; Schnellmann, R.G. Progressive Disruption of the Plasma Membrane during Renal Proximal Tubule Cellular Injury. Toxicol. Appl. Pharmacol. 2001, 171, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Mitsuhiro, H.; Satoshi, A.; Hiroki, I.; Shinichi, T.; Noboru, O.; Noriyuki, I.; Nobutaka, S. Restoration of myoglobin native fold from its initial state of amyloid formation by trehalose. J. Phy. Chem. B 2018, 122, 11962–11968. [Google Scholar] [CrossRef]
- Patist, A.; Zoerb, H. Preservation mechanisms of trehalose in food and biosystems. Colloids Surf. B Biointerfaces 2005, 40, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Xin, R.; Qi, S.; Zeng, C.; Khan, F.I.; Yang, B.; Wang, Y. A functional natural deep eutectic solvent based on trehalose: Structural and physicochemical properties. Food Chem. 2017, 217, 560–567. [Google Scholar] [CrossRef]
- Wang, N.-Q.; Li, J.; Sun, J.; Huang, J.; Wang, P. Bioreduction of 3,5-bis(trifluoromethyl)acetophenone using ionic liquid as a co-solvent catalyzed by recombinant Escherichia coli cells. Biochem. Eng. J. 2015, 101, 119–125. [Google Scholar] [CrossRef]
Sample Availability: Samples of (S)-3,5-BTPE are available from the authors. |










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | NADH | NADPH | Specific Activity (U/g) |
|---|---|---|---|
| + | + | − | 0.37 ± 0.04 |
| + | − | + | none |
| Group | NADH Content μmol/g Cell (DCW) b | Yield (%) a | ee (%) a |
|---|---|---|---|
| Control | 26.9 ± 0.01 | 41.54 ± 0.58 | >99.9 |
| Oxygen-deficient environment | 40.3 ± 0.33 | 90.47 ± 0.54 | >99.9 |
| Control | Oxygen-Deficient Environment | |||
|---|---|---|---|---|
| Catalyst | Yield (%) | ee (%) | Yield (%) | ee (%) |
| C. tropicalis 104 a | 68.8 ± 3.54 | >99.9 | 91.3 ± 1.84 | >99.9 |
| G. candidum ZJPH1704 b | 2.5 ± 1.23 | 66.9 ± 0.84 | 60.4 ± 2.91 | >99.0 |
| R. erythropolis XS1012 c | 78.7 ± 1.69 | >99.9 | 74.4 ± 3.55 | >99.9 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Z.; Bi, S.; Ye, N.; Wang, P. High-Efficient Production of (S)-1-[3,5-Bis(trifluoromethyl)phenyl]ethanol via Whole-Cell Catalyst in Deep-Eutectic Solvent-Containing Micro-Aerobic Medium System. Molecules 2020, 25, 1855. https://doi.org/10.3390/molecules25081855
Zhu Z, Bi S, Ye N, Wang P. High-Efficient Production of (S)-1-[3,5-Bis(trifluoromethyl)phenyl]ethanol via Whole-Cell Catalyst in Deep-Eutectic Solvent-Containing Micro-Aerobic Medium System. Molecules. 2020; 25(8):1855. https://doi.org/10.3390/molecules25081855
Chicago/Turabian StyleZhu, Zhiren, Shunde Bi, Ning Ye, and Pu Wang. 2020. "High-Efficient Production of (S)-1-[3,5-Bis(trifluoromethyl)phenyl]ethanol via Whole-Cell Catalyst in Deep-Eutectic Solvent-Containing Micro-Aerobic Medium System" Molecules 25, no. 8: 1855. https://doi.org/10.3390/molecules25081855
APA StyleZhu, Z., Bi, S., Ye, N., & Wang, P. (2020). High-Efficient Production of (S)-1-[3,5-Bis(trifluoromethyl)phenyl]ethanol via Whole-Cell Catalyst in Deep-Eutectic Solvent-Containing Micro-Aerobic Medium System. Molecules, 25(8), 1855. https://doi.org/10.3390/molecules25081855