
Study on Cecropin B2 Production via Construct Bearing Intein Oligopeptide Cleavage Variants



 and
and 
Abstract
1. Introduction
2. Results
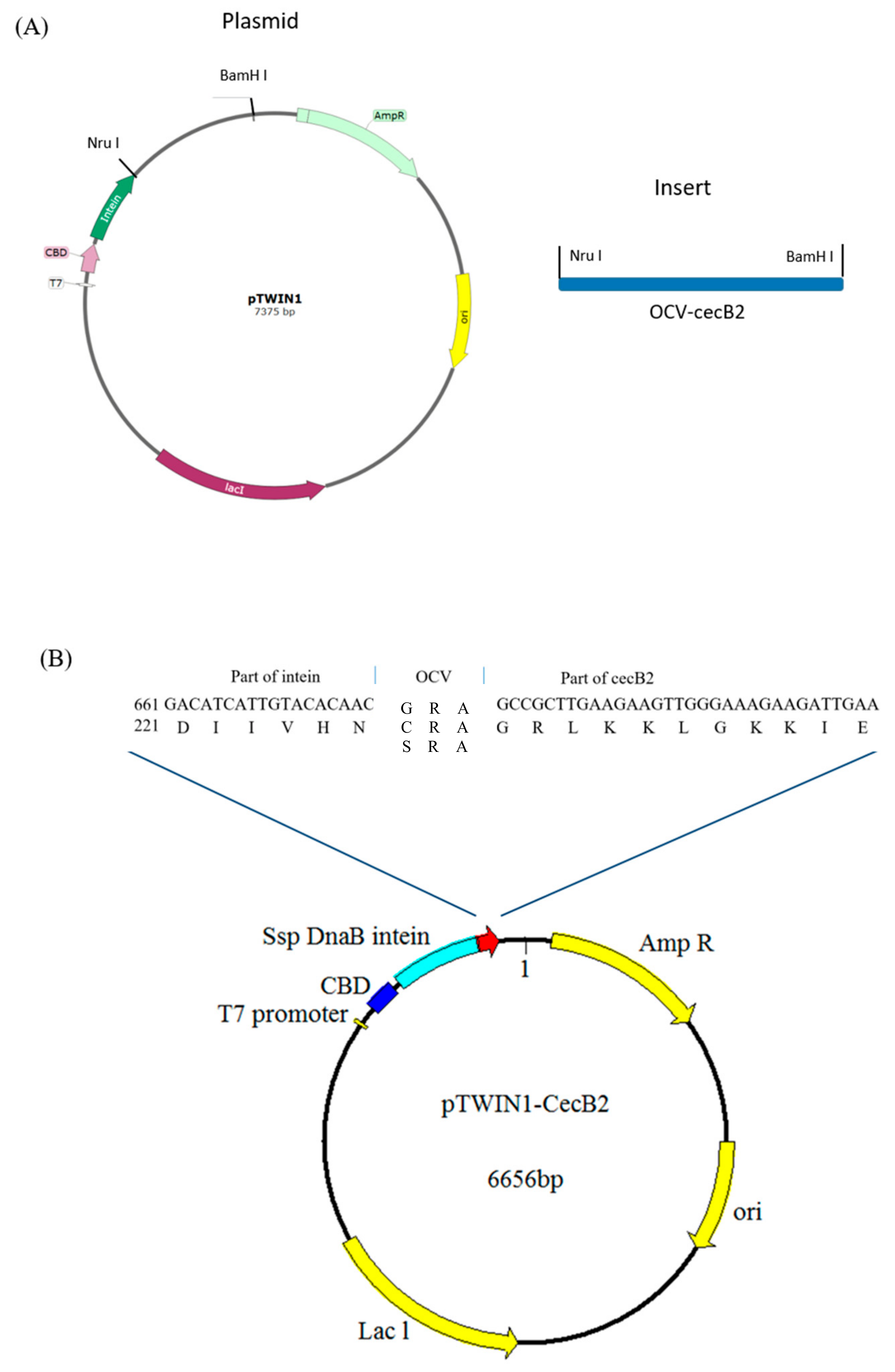
2.1. Gene Construction
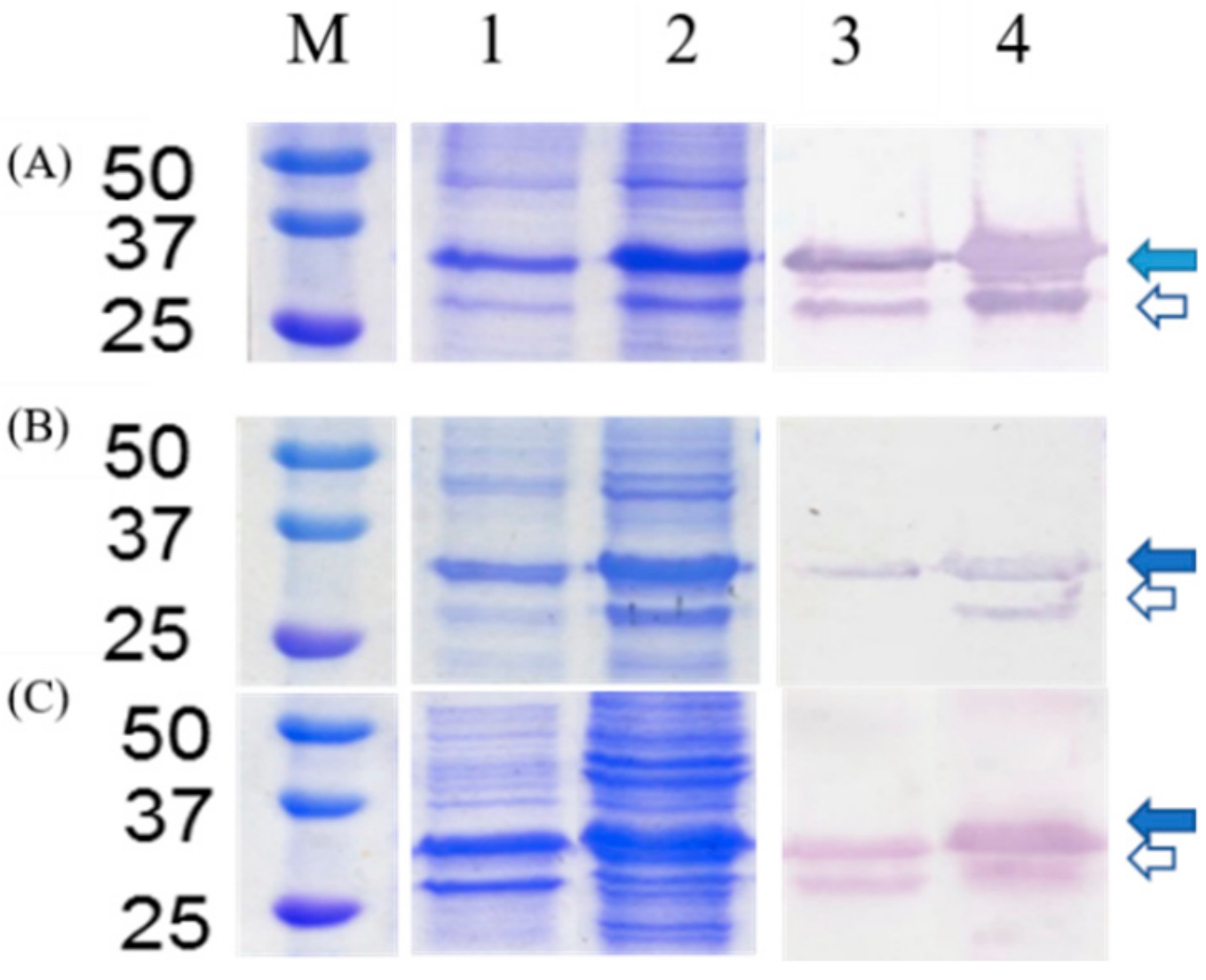
2.2. Fusion Protein Expression
2.3. Determining Cleavage Timescale for Various OCV
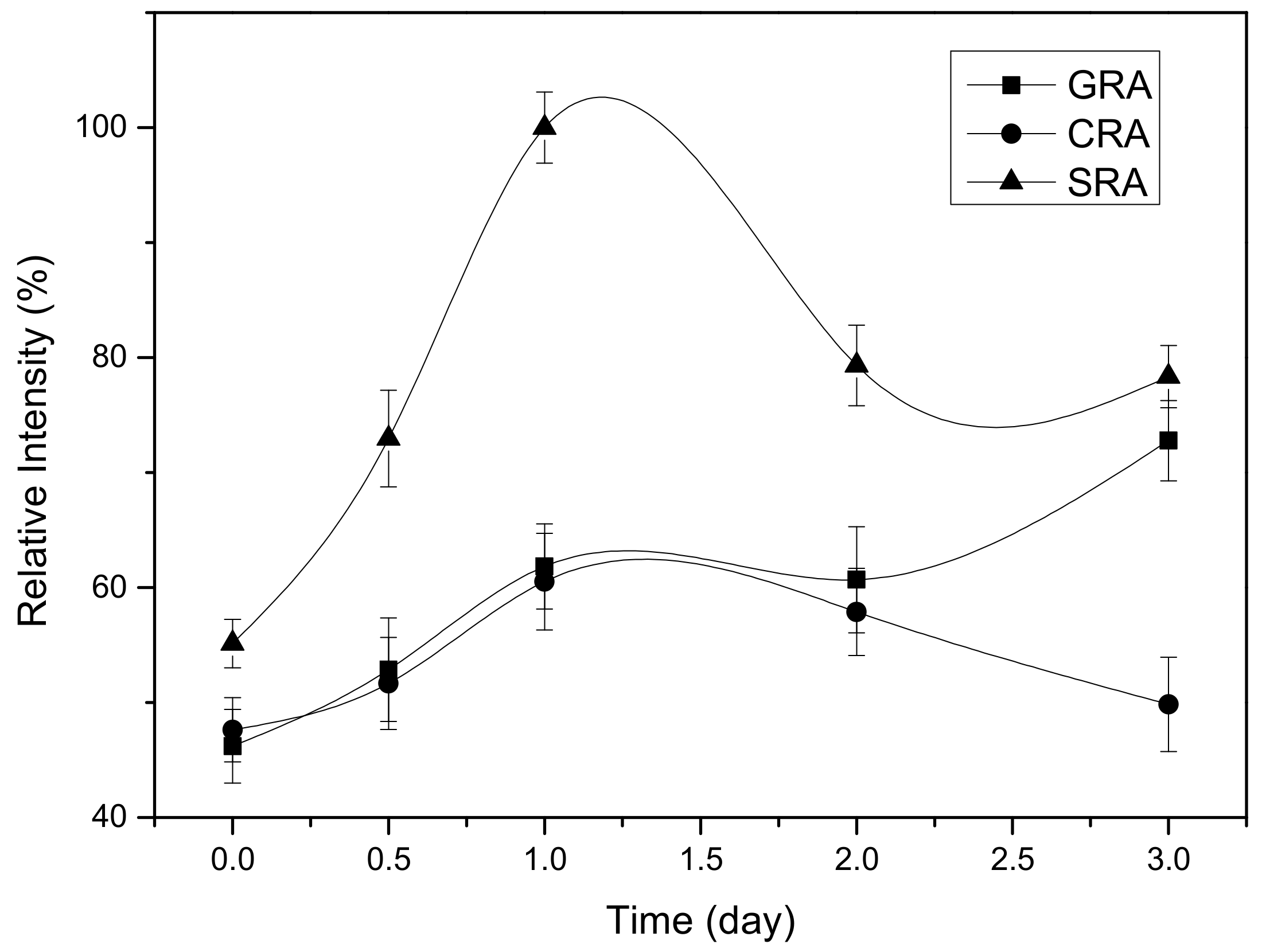
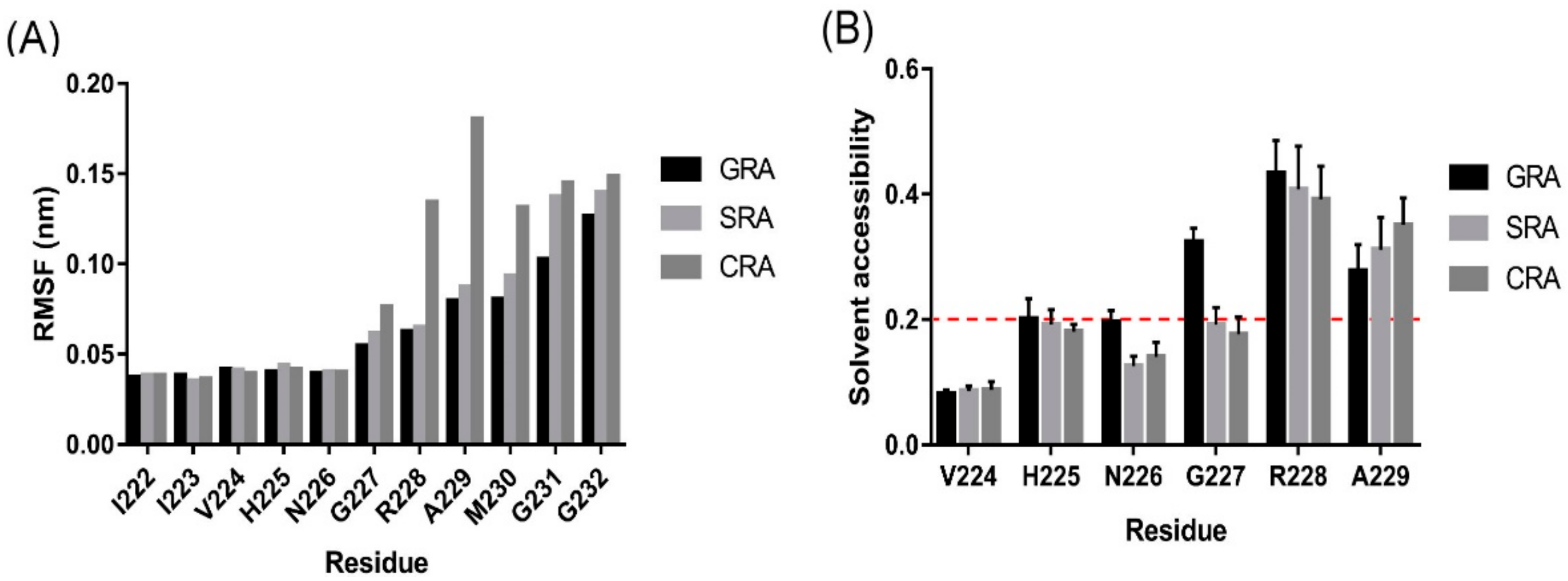
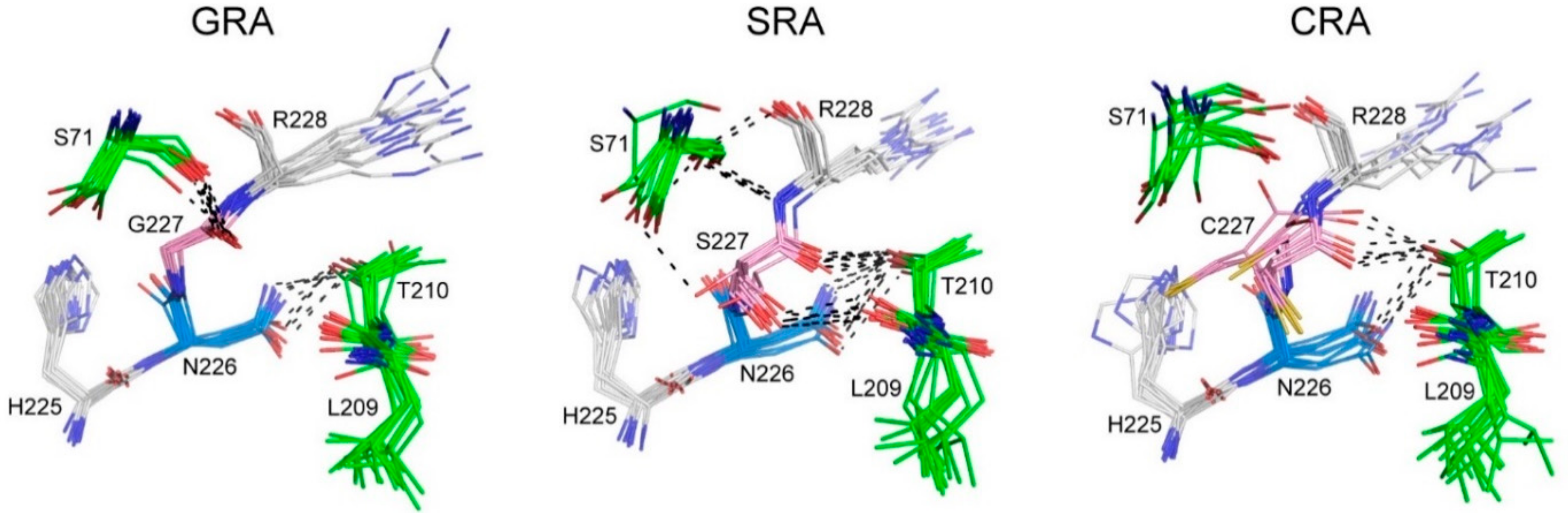
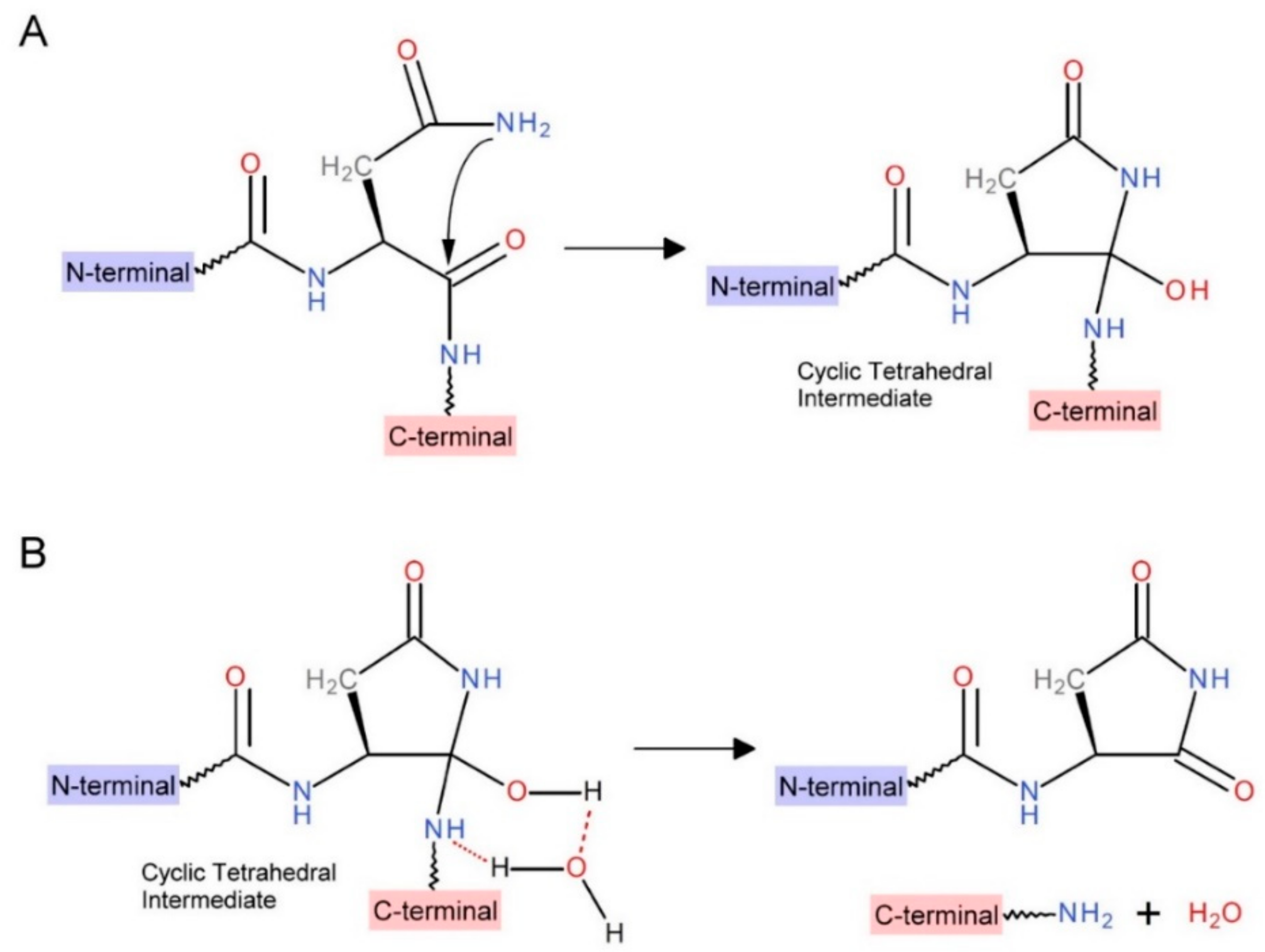
2.4. Kinetic Study of INT Cleavage Reaction
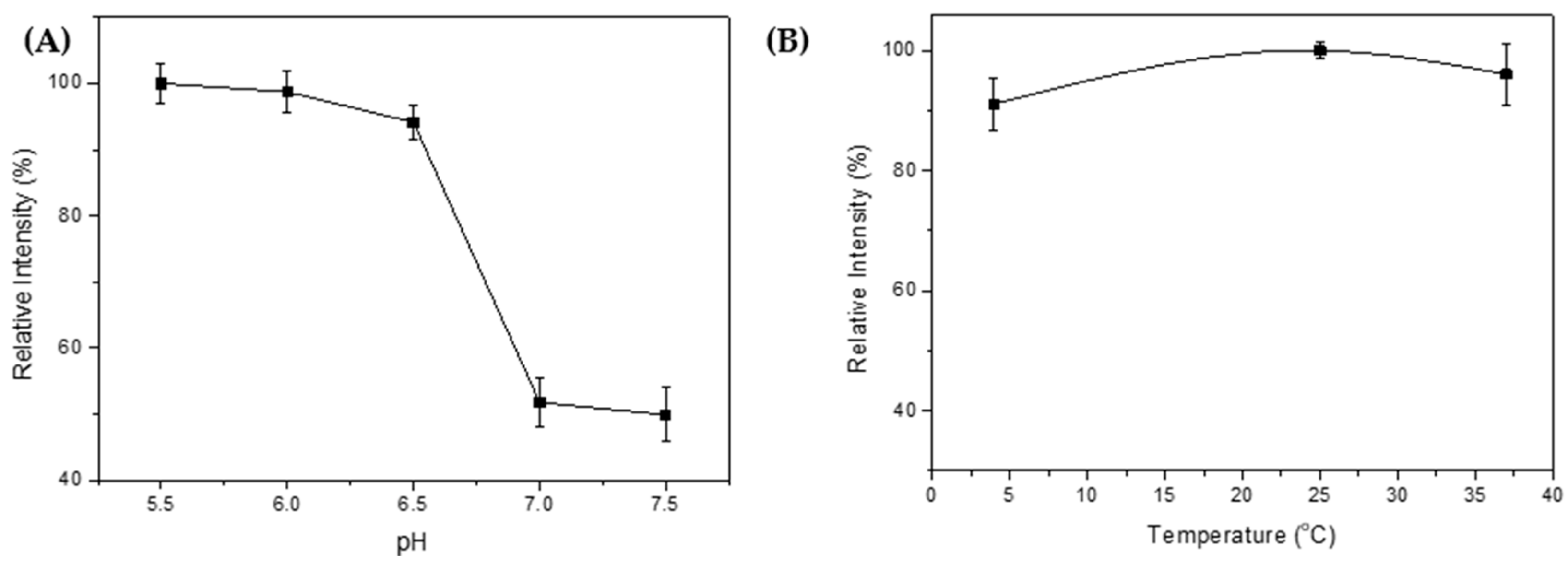
2.5. Intein Cleavage Conditions
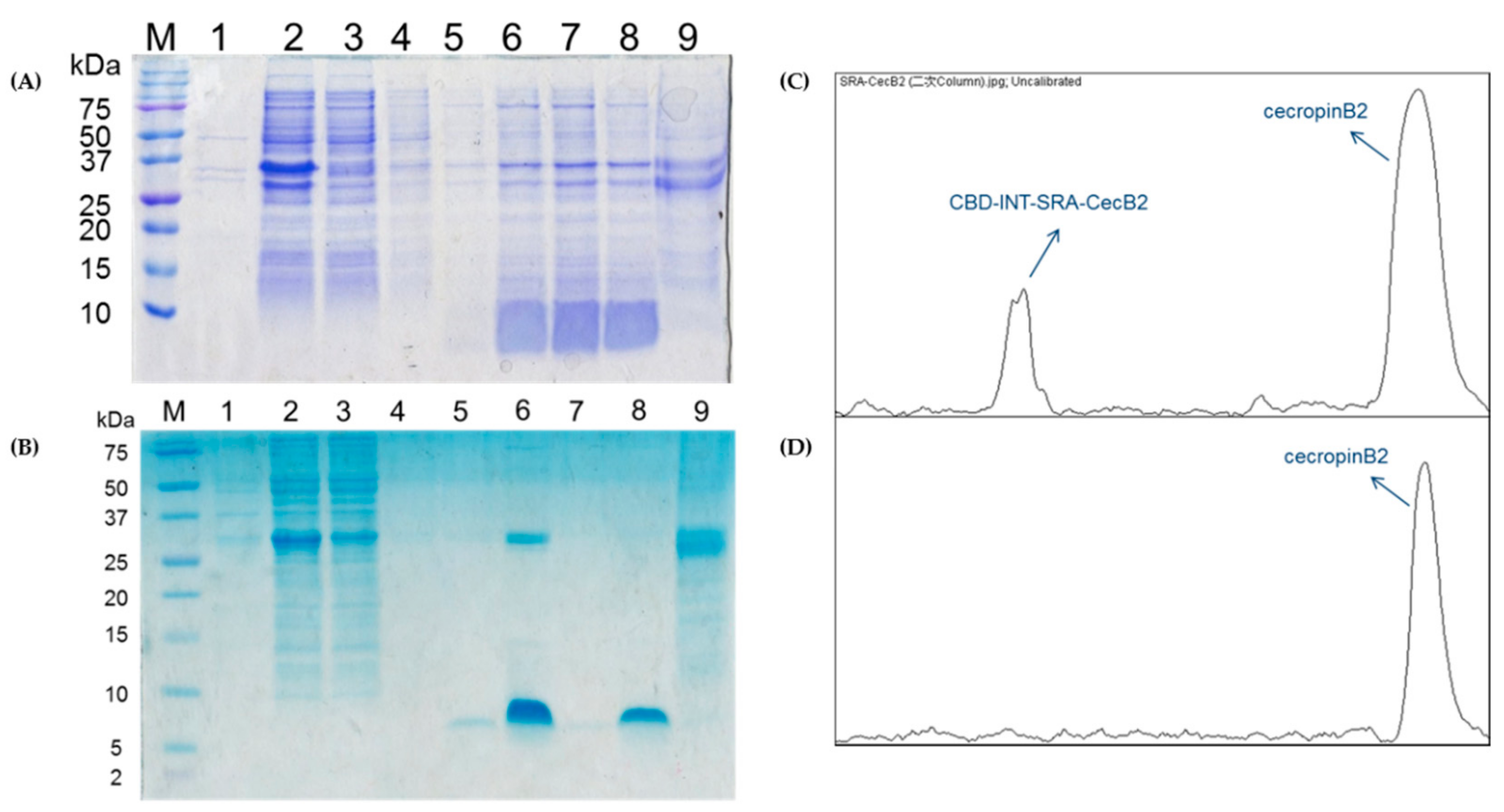
2.6. In-Column Purification for CecB2 Production and Its Activity
3. Discussion
4. Materials and Methods
4.1. Strains, Plasmids, and Cecropin B2
4.2. Construction of Expression Systems
4.3. Cultivation Conditions
4.4. CecB2 Production
4.5. Assays
4.6. Protein Modeling and in Silico MD Simulation
4.7. Kinetics Models
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hancock, R.E.; Scott, M.G. The role of antimicrobial peptides in animal defenses. Proc. Natl. Acad. Sci. USA 2000, 97, 8856–8861. [Google Scholar] [CrossRef] [PubMed]
- Bulet, P.; Hetru, C.; Dimarcq, J.-L.; Hoffmann, D. Antimicrobial peptides in insects; structure and function. Dev. Comp. Immunol. 1999, 23, 329–344. [Google Scholar] [CrossRef]
- Rajanbabu, V.; Chen, J.-Y. Applications of antimicrobial peptides from fish and perspectives for the future. Peptides 2011, 32, 32–415. [Google Scholar] [CrossRef]
- Broekaert, W.F.; Cammue, B.P.A.; DeBolle, M.F.C.; Thevissen, K.; DeSamblanx, G.W.; Osborn, R.W. Antimicrobial peptides from plants. Crit. Rev. Plant Sci. 1997, 16, 297–323. [Google Scholar] [CrossRef]
- Lai, W.-S.; Kan, S.-C.; Lin, C.-C.; Shieh, C.-J.; Liu, Y.-C. Antibacterial Peptide CecropinB2 Production via Various Host and Construct Systems. Molecules 2016, 21, 103. [Google Scholar] [CrossRef] [PubMed]
- Moore, A.J.; A Devine, D.; Bibby, M.C. Preliminary experimental anticancer activity of cecropins. Pept. Res. 1994, 7, 7. [Google Scholar]
- Gazit, E.; Lee, W.-J.; Brey, P.T.; Shai, Y. Mode of Action of the Antibacterial Cecropin B2: A Spectrofluorometric Study. Biochemistry 1994, 33, 10681–10692. [Google Scholar] [CrossRef]
- Fu, H.; Björstad, Åse; Dahlgren, C.; Bylund, J. A bactericidal cecropin-a peptide with a stabilized α-helical structure possess an increased killing capacity but no proinflammatory activity. Inflammation 2004, 28, 337–343. [Google Scholar] [CrossRef]
- Steiner, H.; Andreu, D.; Merrifield, R. Binding and action of cecropin and cecropin analogues: Antibacterial peptides from insects. Biochim. et Biophys. Acta (BBA) - Biomembr. 1988, 939, 260–266. [Google Scholar] [CrossRef]
- Zhang, J.; Movahedi, A.; Wei, Z.; Sang, M.; Wu, X.; Wang, M.; Wei, H.; Pan, H.; Yin, T.; Zhuge, Q. High-level SUMO-mediated fusion expression of ABP-dHC-cecropin A from multiple joined genes in Escherichia coli. Anal. Biochem. 2016, 509, 15–23. [Google Scholar] [CrossRef]
- Xu, X.; Jin, F.; Yu, X.; Ji, S.; Wang, J.; Chen, H.; Wang, C.; Zhang, W. Expression and purification of a recombinant antibacterial peptide, cecropin, from Escherichia coli. Protein Expr. Purif. 2007, 53, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Su, Y.; Li, J.; Sun, J.; Yang, Y. Expression and purification of the antimicrobial peptide cecropin AD by fusion with cationic elastin-like polypeptides. Protein Expr. Purif. 2012, 85, 200–203. [Google Scholar] [CrossRef] [PubMed]
- Noor, M.; Dworeck, T.; Schenk, A.; Shinde, P.; Fioroni, M.; Schwaneberg, U. Polymersome surface decoration by an EGFP fusion protein employing Cecropin A as peptide “anchor”. J. Biotechnol. 2012, 157, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Sang, M.; Wei, H.; Zhang, J.; Wei, Z.; Wu, X.; Chen, Y.; Zhuge, Q. Expression and characterization of the antimicrobial peptide ABP-dHC-cecropin A in the methylotrophic yeast Pichia pastoris. Protein Expr. Purif. 2017, 140, 44–51. [Google Scholar] [CrossRef]
- Xia, L.; Liu, Z.; Ma, J.; Sun, S.; Yang, J.; Zhang, F. Expression, purification and characterization of cecropin antibacterial peptide from Bombyx mori in Saccharomyces cerevisiae. Protein Expr. Purif. 2013, 90, 47–54. [Google Scholar] [CrossRef]
- Tellez, G.; Castaño-Osorio, J.C. Expression and purification of an active cecropin-like recombinant protein against multidrug resistance Escherichia coli. Protein Expr. Purif. 2014, 100, 48–53. [Google Scholar] [CrossRef]
- Fang, Y.-T.; Lai, W.-S.; Liu, J.-H.; Liu, Y.-C. Enhanced cecropin B2 production via chitin-binding domain and intein self-cleavage system. Biotechnol. Appl. Biochem. 2018, 66, 209–215. [Google Scholar] [CrossRef]
- Mathys, S.; Evans, T.C.; Chute, I.C.; Wu, H.; Chong, S.; Benner, J.; Liu, X.-Q.; Xu, M.-Q. Characterization of a self-splicing mini-intein and its conversion into autocatalytic N- and C-terminal cleavage elements: Facile production of protein building blocks for protein ligation. Gene 1999, 231, 1–13. [Google Scholar] [CrossRef]
- Yan, S.-S.; Yan, J.; Shi, G.; Xu, Q.; Chen, S.-C.; Tian, Y.-W. Production of native protein by using Synechocystis sp. PCC6803 DnaB mini-intein in Escherichia coli. Protein Expr. Purif. 2005, 40, 340–345. [Google Scholar] [CrossRef]
- Xu, C.-G.; Fan, X.-J.; Fu, Y.; Liang, A.-H. Effect of location of the His-tag on the production of soluble and functional Buthus martensii Karsch insect toxin. Protein Expr. Purif. 2008, 59, 103–109. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, Z.H.; Chen, G.Q. Microbial polyhydroxyalkanote synthesis repression protein PhaR as an affinity tag for recombinant protein purification. Microb. Cell Factories 2010, 9, 28. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Wan, Y.; Liu, J.; Yang, X.; Wang, H.; Tao, K.; Han, J.; Shi, J.; Hu, D. Expression and purification of rhIL-10-RGD from Escherichia coli as a potential wound healing agent. J. Microbiol. Methods 2016, 127, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Chen, J.; Yao, H.; Liu, L.; Wang, J.; Zhang, J.; Liu, J.-N. Use of Ssp dnaB derived mini-intein as a fusion partner for production of recombinant human brain natriuretic peptide in Escherichia coli. Protein Expr. Purif. 2005, 43, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Esipov, R.; Stepanenko, V.N.; Chupova, L.A.; Boyarskikh, U.A.; Filipenko, M.L.; Miroshnikov, A.I. Production of recombinant human epidermal growth factor using Ssp dnaB mini-intein system. Protein Expr. Purif. 2008, 61, 1–6. [Google Scholar] [CrossRef]
- Setrerrahmane, S.; Zhang, Y.; Dai, G.; Lv, J.; Tan, S. Efficient Production of Native Lunasin with Correct N-terminal Processing by Using the pH-Induced Self-Cleavable Ssp DnaB Mini-intein System in Escherichia coli. Appl. Biochem. Biotechnol. 2014, 174, 612–622. [Google Scholar] [CrossRef]
- Ebbensgaard, A.E.; Mordhorst, H.; Overgaard, M.T.; Nielsen, C.G.; Aarestrup, F.M.; Hansen, E.B. Comparative Evaluation of the Antimicrobial Activity of Different Antimicrobial Peptides against a Range of Pathogenic Bacteria. PLoS ONE 2015, 10, e0144611. [Google Scholar] [CrossRef]
- Deslouches, B.; Di, Y.P. Antimicrobial peptides with selective antitumor mechanisms: prospect for anticancer applications. Oncotarget 2017, 8, 46635–46651. [Google Scholar] [CrossRef]
- Wang, J.; Ma, K.; Ruan, M.; Wang, Y.; Li, Y.; Fu, Y.V.; Song, Y.; Sun, H.; Wang, J. A novel cecropin B-derived peptide with antibacterial and potential anti-inflammatory properties. PeerJ 2018, 6, e5369. [Google Scholar] [CrossRef]
- Catak, S.; Monard, G.; Aviyente, V.; Ruiz-Loópez, M.F. Computational Study on Nonenzymatic Peptide Bond Cleavage at Asparagine and Aspartic Acid. J. Phys. Chem. A 2008, 112, 8752–8761. [Google Scholar] [CrossRef]
- Martínez, L. Automatic Identification of Mobile and Rigid Substructures in Molecular Dynamics Simulations and Fractional Structural Fluctuation Analysis. PLoS ONE 2015, 10, e0119264. [Google Scholar]
- Mujika, J.; Lopez, X.; Mulholland, A.J. Modeling Protein Splicing: Reaction Pathway for C-Terminal Splice and Intein Scission. J. Phys. Chem. B 2009, 113, 5607–5616. [Google Scholar] [CrossRef] [PubMed]
- Mujika, J.; Lopez, X. Unveiling the Catalytic Role of B-Block Histidine in the N–S Acyl Shift Step of Protein Splicing. J. Phys. Chem. B 2017, 121, 7786–7796. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Russel, D. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Schägger, H. Tricine-SDS-PAGE. Nat. Protoc. 2006, 1, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Mistry, I.N.; Tavassoli, A. Reprogramming the Transcriptional Response to Hypoxia with a Chromosomally Encoded Cyclic Peptide HIF-1 Inhibitor. ACS Synth. Boil. 2016, 6, 518–527. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Ekengren, S.; Hultmark, D. Drosophila cecropin as an antifungal agent. Insect Biochem. Mol. Boil. 1999, 29, 965–972. [Google Scholar] [CrossRef]
- Wiegand, I.; Hilpert, K.; Hancock, R.E. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef]
- Kelley, L.A.; E Sternberg, M.J. Protein structure prediction on the Web: a case study using the Phyre server. Nat. Protoc. 2009, 4, 363–371. [Google Scholar] [CrossRef]
- Pierce, B.; Wiehe, K.; Hwang, H.; Kim, B.-H.; Vreven, T.; Weng, Z. ZDOCK server: interactive docking prediction of protein-protein complexes and symmetric multimers. Bioinformatics 2014, 30, 1771–1773. [Google Scholar] [CrossRef]
- Webb, B.; Sali, A. Comparative Protein Structure Modeling Using Modeller. In Current Protocols in Bioinformatics; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016. [Google Scholar]
- Wiederstein, M.; Sippl, M.J. ProSA-web: interactive web service for the recognition of errors in three-dimensional structures of proteins. Nucleic Acids Res. 2007, 35, W407–W410. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: a program to check the stereochemical quality of protein structures. J. Appl. Crystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Pronk, S.; Páll, S.; Schulz, R.; Larsson, P.; Bjelkmar, P.; Apostolov, R.; Shirts, M.R.; Smith, J.C.; Kasson, P.M.; Van Der Spoel, D.; et al. GROMACS 4.5: a high-throughput and highly parallel open source molecular simulation toolkit. Bioinformatics 2013, 29, 845–854. [Google Scholar] [CrossRef] [PubMed]
- Cavallo, L. POPS: A fast algorithm for solvent accessible surface areas at atomic and residue level. Nucleic Acids Res. 2003, 31, 3364–3366. [Google Scholar] [CrossRef] [PubMed]
- Schrodinger, L.L.C. The PyMOL Molecular Graphics System; Version 1.8; Schrodinger, L.L.C.: New York, NY, USA, 2015. [Google Scholar]
Sample Availability: Samples of the compounds are available from the authors |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Primer | Sequence (5′-3′) | Restriction Enzyme |
|---|---|---|---|
| CBD-INT-GRA-CecB2 | GRA-F’ | GTC GCG AAT GAC ATC ATT GTA CAC AAC GGC CGG GCG GGC CGC TTG AAG AAG | Nru I |
| R’-GRA | CGC GGA TCC TTA TCT TCC TAG TGC CTG | BamH I | |
| CBD-INT-CRA-CecB2 | CRA-F’ | GTC GCG AAT GAC ATC ATT GTA CAC AAC TGC AGA GCC GGC CGC TTG AAG AAG | Nru I |
| R’-CRA | CGC GGA TCC TTA TCT TCC TAG TGC CTG | BamH I | |
| CBD-INT-SRA-CecB2 | SRA-F’ | GTC GCG AAT GAC ATC ATT GTA CAC AAC AGC AGA GCC GGC CGC TTG AAG AAG | Nru I |
| R’-SRA | CGC GGA TCC TTA TCT TCC TAG TGC CTG | BamH I |
| Item | GRA | CRA | SRA |
|---|---|---|---|
| Rate constant k (d−1) a | 0.292 ± 0.013 | 0.240 ± 0.044 | 0.595 ± 0.020 |
| R-square | 0.996 | 0.934 | 0.998 |
| Strain | Serial Dilution Number a |
|---|---|
| E. coli JM109 | 5.0 |
| E. coli DH5α | 4.0 |
| E. coli BL21 | 4.3 |
| Strain or Plasmid | Genotype and Relevant Characteristics | Source |
|---|---|---|
| DH5α | F– endA1 glnV44 thi-1 recA1 relA1 gyrA96 deoR nupG Φ80dlacZ △M15 △ (lacZYA-argF)U169, hsdR17(rK− mK+) λ– | Novagen, USA |
| ER 2566 | F-λ- fhuA2 [lon] ompT lacZ::T7 gene 1 gal sulA 11 △(mcrC-mrr)114::IS10R (mcr-73::miniTn10-TetS)2 R(zgb-210::Tn10)(TetS)endA1 [dcm] | New England Biolabs, USA |
| T-vector | pBluescript IISK(−) with modified MCS | Yeastern biotech |
| pTWIN1 | Cloning vector. Amp | New England Biolabs, USA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, Y.-T.; Li, S.-Y.; Hu, N.-J.; Yang, J.; Liu, J.-H.; Liu, Y.-C. Study on Cecropin B2 Production via Construct Bearing Intein Oligopeptide Cleavage Variants. Molecules 2020, 25, 1005. https://doi.org/10.3390/molecules25041005
Fang Y-T, Li S-Y, Hu N-J, Yang J, Liu J-H, Liu Y-C. Study on Cecropin B2 Production via Construct Bearing Intein Oligopeptide Cleavage Variants. Molecules. 2020; 25(4):1005. https://doi.org/10.3390/molecules25041005
Chicago/Turabian StyleFang, Yi-Ting, Si-Yu Li, Nien-Jen Hu, Jie Yang, Jyung-Hurng Liu, and Yung-Chuan Liu. 2020. "Study on Cecropin B2 Production via Construct Bearing Intein Oligopeptide Cleavage Variants" Molecules 25, no. 4: 1005. https://doi.org/10.3390/molecules25041005
APA StyleFang, Y.-T., Li, S.-Y., Hu, N.-J., Yang, J., Liu, J.-H., & Liu, Y.-C. (2020). Study on Cecropin B2 Production via Construct Bearing Intein Oligopeptide Cleavage Variants. Molecules, 25(4), 1005. https://doi.org/10.3390/molecules25041005