Detection of the agr System and Resistance to Antimicrobials in Biofilm-Producing S. epidermidis

 ,
,  , and
, and 
Abstract
1. Introduction
2. Results
2.1. Identification of Strains
2.2. Detection of Biofilm Genes
2.3. Detection of mRNA of Biofilm Genes in CoNS Samples
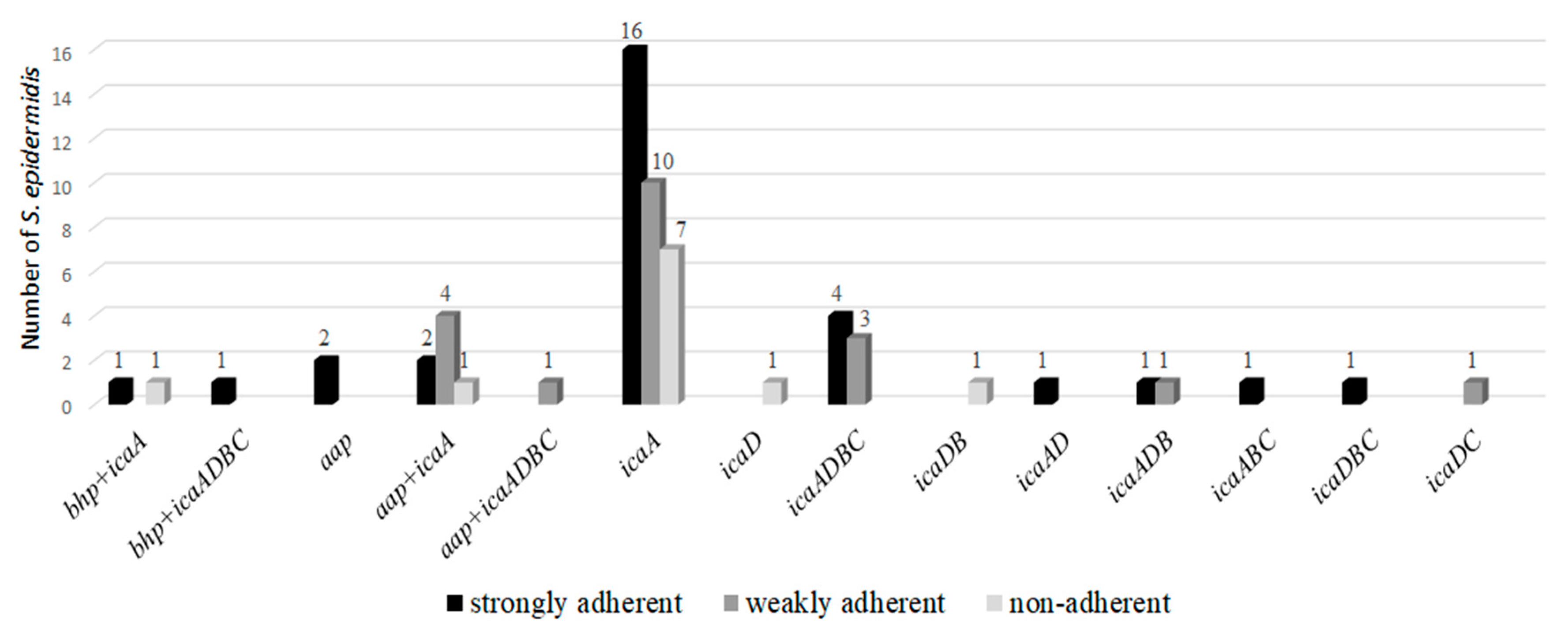
2.4. Investigation of Biofilm Production by Adherence to Polystyrene Plates
2.5. Determination of the agr Locus in Biofilm-Producing S. epidermidis
2.6. Detection of mecA Gene and SCCmec in Biofilm-Producing S. epidermidis
2.7. Determination of Minimum Inhibitory Concentration (MIC) of Antimicrobials in Biofilm-Producing S. epidermidis
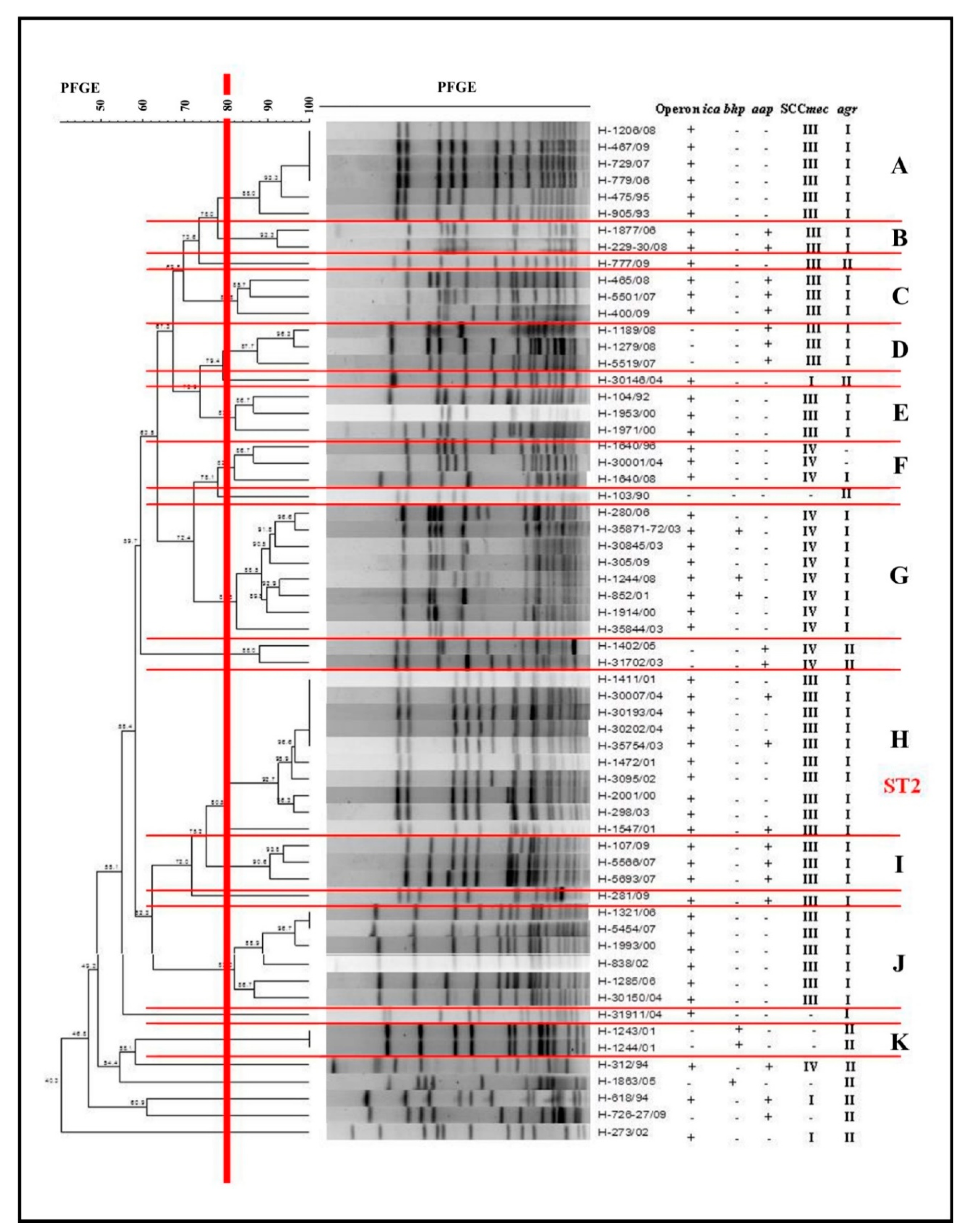
2.8. Determination of the Clonal Profile of the Biofilm-Producer S. epidermidis
3. Discussion
4. Materials and Methods
4.1. Strains
4.2. DNA Extraction
4.3. Detection of the Biofilm and mecA Gene
4.4. RNA Extraction and RT-PCR
4.5. Investigation of Biofilm Production by Adherence to Polystyrene Plates
4.6. Determination of SCCmec
4.7. Determination of Minimum Inhibitory Concentration by the E-Test
4.8. Determination of agr Group
4.9. Pulsed-Field Gel Electrophoresis
4.10. Determination of Multilocus Sequence Typing (MLST)
5. Conclusions
- In some strains, biofilm formation may be additionally or exclusively mediated by proteins, such as Bhp or Aap.
- The agrI was associated with the ica operon and aap gene and strains that were positive for bhp gene expression showed agrII.
- The mecA gene was positive in 90.1% of the S. epidermidis capable of producing biofilm.
- The SCCmec typing made possible the characterization of SCCmec I, III, and IV in these samples, with SCCmec III being the most frequent.
- The expression of the ica operon, agrI locus, and SCCmec III was found in clusters A, H, and J, and that of SCCmec IV in G.
- The MLST analysis characterized the major cluster (H) as ST2.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kloos, W.E. Staphylococcus and Micrococcus. In Manual of Clinical Microbiology; Murray, P.R., Baron, E.J., Pfaller, M.A., Tenover, F.C., Yolken, R.H., Eds.; American Society Microbiology: Washington, DC, USA, 1999; pp. 264–282. [Google Scholar]
- Rowlinson, M.C.; LeBourgeois, P.; Ward, K.; Song, Y.; Finegold, S.M.; Bruckner, D.A. Isolation of a strictly anaerobic strain of Staphylococcus epidermidis. J. Clin. Microbiol. 2006, 44, 857–860. [Google Scholar] [CrossRef]
- Fey, P.D.; Olson, M.E. Current concepts in biofilm formation of Staphylococcus epidermidis. Future Microbiol. 2010, 5, 917–933. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Virulence factors of the coagulase-negative staphylococci. Front. Biosci. 2004, 9, 841. [Google Scholar] [CrossRef] [PubMed]
- Cucarella, C.; Tormo, M.Á.; Úbeda, C.; Trotonda, M.P.; Monzón, M.; Peris, C.; Amorena, B.; Lasa, Í.; Penadés, J.R. Role of Biofilm-Associated Protein Bap in the Pathogenesis of Bovine Staphylococcus aureus. Infect. Immun. 2004, 72, 2177–2185. [Google Scholar] [CrossRef] [PubMed]
- Novick, R.P.; Geisinger, E. Quorum sensing in staphylococci. Annu. Rev. Genet. 2008, 42, 541–564. [Google Scholar] [CrossRef] [PubMed]
- McCulloch, J.A. Avaliação da Funcionalidade do Locus Acessory Gene Regulator (Agr) em Cepas de Staphylococcus aureus Brasileiras com Suscetibilidade Reduzida aos Glicopeptídeos. Ph.D. Thesis, Universidade de São Paulo, Sao Paulo, Brazil, 2006. [Google Scholar]
- Li, M.; Guan, M.; Jiang, X.F.; Yuan, F.Y.; Xu, M.; Zhang, W.Z.; Lu, Y. Genetic polymorphism of the accessory gene regulator (agr) locus in Staphylococcus epidermidis and its association with pathogenicity. J. Med. Microbiol. 2004, 53, 545–549. [Google Scholar] [CrossRef] [PubMed]
- Huseby, M.; Shi, K.; Kent Brown, C.; Digre, J.; Mengistu, F.; Keun, S.S.; Bohach, G.A.; Schlievert, P.M.; Ohlendorf, D.H.; Earhart, C.A. Structure and biological activities of beta toxin from Staphylococcus aureus. J. Bacteriol. 2007, 189, 8719–8726. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Staphylococcal biofilms. Curr. Top. Microbiol. Immunol. 2008, 322, 207–228. [Google Scholar] [CrossRef]
- Lázaro-Díez, M.; Remuzgo-Martínez, S.; Rodríguez-Mirones, C.; Acosta, F.; Icardo, J.M.; Martínez-Martínez, L.; Ramos-Vivas, J. Effects of Subinhibitory Concentrations of Ceftaroline on Methicillin-Resistant Staphylococcus aureus (MRSA) Biofilms. PLoS ONE 2016, 11, e0147569. [Google Scholar] [CrossRef]
- Bouchami, O.; Achour, W.; Mekni, M.A.; Rolo, J.; Hassen, A. Ben Antibiotic resistance and molecular characterization of clinical isolates of methicillin-resistant coagulase-negative staphylococci isolated from bacteremic patients in oncohematology. Folia Microbiol. (Praha) 2011, 56, 122–130. [Google Scholar] [CrossRef]
- International Working Group on the Staphylococcal Cassette Chromosome Elements SCCmec about IWG-SCC EN. Available online: http://www.sccmec.org/Pages/SCC_aboutIWG-SCCEN.html (accessed on 17 August 2018).
- Oliveira, A.; Cunha, M.D.L.R. Comparison of methods for the detection of biofilm production in coagulase-negative staphylococci. BMC Res. Notes 2010, 3, 260. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z.; Yang, X.; Yang, L.; Jiang, J.; Ou, Y.; Molin, S.; Qu, D. Formation and properties of in vitro biofilms of ica-negative Staphylococcus epidermidis clinical isolates. J. Med. Microbiol. 2007, 56, 83–93. [Google Scholar] [CrossRef]
- Otto, M. Staphylococcus epidermidis—The “accidental” pathogen. Nat. Rev. Microbiol. 2009, 7, 555–567. [Google Scholar] [CrossRef] [PubMed]
- Bowden, M.G.; Chen, W.; Singvall, J.; Xu, Y.; Peacock, S.J.; Valtulina, V.; Speziale, P. Identification and preliminary characterization of cell-wall-anchored proteins of Staphylococcus epidermidis. Microbiology 2005, 1453–1464. [Google Scholar] [CrossRef] [PubMed]
- Rohde, H.; Frankenberger, S.; Zähringer, U.; Mack, D. Structure, function and contribution of polysaccharide intercellular adhesin (PIA) to Staphylococcus epidermidis biofilm formation and pathogenesis of biomaterial-associated infections. Eur. J. Cell Biol. 2010, 89, 103–111. [Google Scholar] [CrossRef]
- Pinheiro, L.; Brito, C.I.; Oliveira, A.; Pereira, V.C.; Cunha, M.L.R.S. Staphylococcus epidermidis and Staphylococcus haemolyticus: Detection of biofilm genes and biofilm formation in blood culture isolates from patients in a Brazilian teaching hospital. Diagn. Microbiol. Infect. Dis. 2016, 86, 11–14. [Google Scholar] [CrossRef]
- Schoenfelder, S.M.K.; Lange, C.; Eckart, M.; Hennig, S.; Kozytska, S.; Ziebuhr, W. Success through diversit—How Staphylococcus epidermidis establishes as a nosocomial pathogen. Int. J. Med. Microbiol. 2010, 300, 380–386. [Google Scholar] [CrossRef]
- Vuong, C.; Kocianova, S.; Yao, Y.; Carmody, A.B.; Otto, M. Increased Colonization of Indwelling Medical Devices by Quorum-Sensing Mutants of Staphylococcus epidermidis In Vivo. J. Infect. Dis. 2018, 190, 1498–1505. [Google Scholar] [CrossRef]
- Cabrera-Contreras, R.; Morelos-Ramírez, R.; Galicia-Camacho, A.N.; Meléndez-Herrada, E. Antibiotic Resistance and Biofilm production in Staphylococcus epidermidis Strains, Isolated from a Tertiary Care Hospital in Mexico City. Int. Sch. Res. Not. 2013, 2013, 1–5. [Google Scholar] [CrossRef]
- Ito, T.; Katayama, Y.; Hiramatsu, K. Cloning and Nucleotide Sequence Determination of the Entire mec DNA of Pre-Methicillin-Resistant Staphylococcus aureus N315 Cloning and Nucleotide Sequence Determination of the Entire mec DNA of Pre-Methicillin-Resistant Staphylococcus aureus N315. Antimicrob. Agents Chemother. 1999, 43, 1449–1458. [Google Scholar] [CrossRef]
- Andersson, D.I.; Hughes, D. Antibiotic resistance and its cost: Is it possible to reverse resistance? Nat. Rev. Microbiol. 2010, 8, 260–271. [Google Scholar] [CrossRef]
- Szczuka, E.; Jabłońska, L.; Kaznowski, A. Effect of subinhibitory concentrations of tigecycline and ciprofloxacin on the expression of biofilm-associated genes and biofilm structure of Staphylococcus epidermidis. Microbiology 2017, 163, 712–718. [Google Scholar] [CrossRef] [PubMed]
- Koneman, E.W.; Allen, S.D.; Janda, W.M.; Schreckenberger, P.C.; Winn, W.C., Jr. Color Atlas and Textbook of Diagnostic Microbiology, 5th ed.; Lippincott: Philadelphia, PA, USA, 1997. [Google Scholar]
- Cunha, M.D.L.R.S.; Sinzato, Y.K.; Silveira, L.V. Comparison of methods for the identification of coagulase-negative staphylococci. Mem. Inst. Oswaldo Cruz 2004, 99, 855–860. [Google Scholar] [CrossRef] [PubMed]
- Barry, T.; Colleran, G.; Glennon, M.; Dunican, L.K.; Gannon, F. The 16S/23S ribosomal spacer region as a target for DNA probes to identify eubacteria. PCR Methods Appl. 1991, 1, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Couto, I.; Pereira, S.; Miragaia, M.; Santos Sanches, I.; De Lencastre, H. Identification of clinical staphylococcal isolates from humans by internal transcribed spacer PCR. J. Clin. Microbiol. 2001, 39, 3099–3103. [Google Scholar] [CrossRef] [PubMed]
- Pereira, V.C.; Pinheiro, L.; Oliveira, A.; Martins, K.B.; Riboli, D.F.; de Souza, M.D. Expression of superantigens and the agr system in Staphylococcus epidermidis. Microb. Pathogenes. 2018, 115, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Arciola, C.R.; Baldassarri, L.; Montanaro, L. Presence of icaA and icaD Genes and Slime Production in a Collection of Staphylococcal Strains from Catheter-Associated Infections. J. Clin. Microbiol. 2001, 39, 2151–2156. [Google Scholar] [CrossRef] [PubMed]
- Murakami, K.; Minamide, W.; Wada, K.; Nakamura, E.; Teraoka, H.; Watanabe, S. Identification of methicillin-resistant strains of staphylococci by polymerase chain reaction. J. Clin. Microbiol. 1991, 29, 2240–2244. [Google Scholar] [CrossRef]
- Mason, W.J.; Blevins, J.S.; Beenken, K.; Wibowo, N.; Ojha, N.; Smeltzer, M.S. Multiplex PCR Protocol for the Diagnosis of Staphylococcal Infection. J. Clin. Microbiol. 2001, 39, 3332–3338. [Google Scholar] [CrossRef]
- Oliveira, D.C.; de Lencastre, H. Multiplex PCR Strategy for Rapid Identification of Structural Types and Variants of the mec Element in Methicillin-Resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2002, 46, 2155–2161. [Google Scholar] [CrossRef]
- Enright, M.C.; Day, N.P.J.; Davies, C.E.; Peacock, S.J.; Spratt, B.G. Multilocus Sequence Typing for Characterization of Methicillin-Resistant and Methicillin-Susceptible Clones of Staphylococcus aureus Multilocus Sequence Typing for Characterization of Methicillin- Resistant and Methicillin-Susceptible Clones of Staphylococcus. J. Clin. Microbiol. 2000, 38, 1008–1015. [Google Scholar] [PubMed]
- Christensen, G.D.; Simpson, W.A.; Bisno, A.L.; Beachey, E.H. Adherence of slime-producing strains of Staphylococcus epidermidis to smooth surfaces. Infect. Immun. 1982, 37, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Machado, A.B.M.P.; Reiter, K.C.; Paiva, R.M.; Barth, A.L. Distribution of staphylococcal cassette chromosome mec (SCCmec) types I, II, III and IV in coagulase-negative staphylococci from patients attending a tertiary hospital in southern Brazil. J. Med. Microbiol. 2007, 56, 1328–1333. [Google Scholar] [CrossRef] [PubMed]
- McDougal, L.K.; Steward, C.D.; Killgore, G.E.; Chaitram, J.M.; McAllister, S.K.; Tenover, F.C. Pulsed-field gel electrophoresis typing of oxacillin-resistant Staphylococcus aureus isolates from the United States: Establishing a national database. J. Clin. Microbiol. 2003, 41, 5113–5120. [Google Scholar] [CrossRef]
Sample Availability: The samples of compounds are not available from authors. |



{kind=link}
{kind=link}
| Species | icaA | icaB | icaC | icaD | Operon icaADBC | bhp | aap | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | % | N | % | N | % | N | % | N | % | N | % | N | % | |
| S. epidermidis | 119 | 96.8 | 144 | 87.3 | 181 | 81.5 | 179 | 75.8 | 105 | 98.1 | 34 | 91.9 | 103 | 99.0 |
| S. haemolyticus | 0 | 0.0 | 10 | 6.1 | 13 | 5.9 | 23 | 9.4 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 |
| S. hominis | 3 | 2.4 | 7 | 4.2 | 17 | 7.7 | 17 | 7.2 | 1 | 0.9 | 1 | 2.7 | 1 | 1.0 |
| S. warneri | 1 | 0.8 | 3 | 1.8 | 7 | 3.2 | 6 | 2.5 | 1 | 0.9 | 0 | 0.0 | 0 | 0.0 |
| S. lugdunensis | 0 | 0.0 | 0 | 0.0 | 2 | 0.9 | 7 | 3.0 | 0 | 0.0 | 1 | 2.7 | 0 | 0.0 |
| S. capitis | 0 | 0.0 | 1 | 0.6 | 2 | 0.9 | 4 | 1.7 | 0 | 0.0 | 1 | 2.7 | 0 | 0.0 |
| Locus agr | ||||
|---|---|---|---|---|
| N | I | II | Not Detected | |
| Operon icaADBC | 34 | 28 (45.9%) | 4 (6.5%) | 2 (3.3%) |
| bhp | 4 | 0 (0.0%) | 4 (6.5%) | 0 (0.0%) |
| aap | 6 | 4 (6.6%) | 2 (3.3%) | 0 (0.0%) |
| Operon ica + bhp | 3 | 3 (4.9%) | 0 (0.0%) | 0 (0.0%) |
| Operon ica + aap | 14 | 12 (19.7%) | 2 (3.3%) | 0 (0.0%) |
| mecA | SCCmec | ||||
|---|---|---|---|---|---|
| I | III | IV | Untyped | ||
| Operon icaADCB (n = 34) | 33 (54%) | 3 (5.5%) | 22 (40%) | 8 (14.5%) | 0 (0.0%) |
| bhp (n = 4) | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) |
| aap (n = 6) | 6 (9.8%) | 1 (1.8%) | 3 (5.5%) | 2 (3.6%) | 1 (1.8%) |
| Operon ica + bhp (n = 3) | 3 (4.9%) | 0 (0.0%) | 0 (0.0%) | 3 (5.5%) | 0 (0.0%) |
| Operon ica + aap (n = 14) | 13 (21.3%) | 1 (1.8%) | 12 (21.8%) | 1 (1.8%) | 0 (0.0%) |
| Antimicrobial | Breakpoint (Resistant) | MIC50 | MIC90 | Variation | Resistance |
|---|---|---|---|---|---|
| Oxacillin | ≥0.5 | >256 | >256 | 0.047 to >256 | 83.6% |
| Vancomycin | ≥32 | 1.5 | 2 | 0.125 to 2 | 0 |
| Linezolid | ≥8 | 0.25 | 0.38 | 0.094 to 1 | 0 |
| Daptomycin | >1 | 0.125 | 0.19 | 0.019 to 0.5 | 0 |
| Quinupristin/dalfopristin | ≥4 | 0.19 | 0.5 | 0.094 to 2 | 0 |
| Tigecycline | >5 | 0.125 | 0.25 | 0.016 to 0.75 | 4.9% |
| Primer | Nucleotide Sequences (5′ to 3′) | Pb | Products | References |
|---|---|---|---|---|
| agrA1 | GCTGCAACCAAGAAACAACC | 1022 | agrI, II, III | [8] |
| agrA2 | CGTGTATTCATAATATGCTTCGATT | |||
| agrB1 | TATGCAAGCCAAGCACTTGT | 453 | agrIII | |
| agrB2 | GTGCGAAAGCCGATAACAAT | |||
| agrC1 | CCTTGGCTAGTACTACACCTTC | 615 | agrII | |
| agrC2 | GTGCTTGGCTTGCATAAACA | |||
| L1 | GAAGTCGTAACAAGG | - | 16S | [28,29] |
| G1 | CAAGGCATCCACCGT | 23S | ||
| RNAr16S 1 | CCTATAAGACTGGGATAACTTCGGG | 791 | RNAr 16S | [33] |
| RNAr16S 2 | CTTTGAGTTTCAACCTTGCGGTCG | |||
| icaA1 | TCTCTTGCAGGAGCAATCAA | 187 | IcaA | [31] |
| icaA2 | TCAGGCACTAACATCCAGCA | |||
| icaB1 | CTGATCAAGAATTTAAATCACAAA | 302 | IcaB | |
| icaB2 | AAAGTCCCATAAGCCTGTTT | |||
| icaC1 | TAACTTTAGGCGCATATGTTT | 400 | IcaC | |
| icaC2 | TTCCAGTTAGGCTGGTATTG | |||
| icaD1 | ATGGTCAAGCCCAGACAGAG | 198 | IcaD | |
| icaD2 | CGTGTTTTCAACATTTAATGCAA | |||
| bhp1 | ATGAAAAATAAACAAGGATTTC | 1300 | Bhp | |
| bhp2 | GCCTAAGCTAGATAATGTTTG | |||
| aap1 | ATGGGCAAACGTAGACAAG | 1100 | Aap | |
| aap2 | ACCGTAAAAATCGTAATTATCTC | |||
| mecA1 | AAAATCGATGGTAAAGGTTGG | 533 | PBP2a | [32] |
| mecA2 | AGTTCTGCAGTACCGGATTTG | |||
| DCS F2 | CATCCTATGATAGCTTGGTC | 342 | SCCmec I, II e IV | [34] |
| DCS R1 | CTAAATCATAGCCATGACCG | |||
| CIF2 F2 | TTCGAGTTGCTGGATGAAGAAGG | 495 | SCCmec I | |
| CIF2 R2 | ATTTACCACAAGGACTACCAGC | |||
| KDP F1 | AATCATCTGCCATTGGTGATGC | 284 | SCCmec II | |
| KDP R1 | CGAATGAAGTGAAAGAAAGTGG | |||
| RIF4 F3 | GTGATTGTTCGAGATATGTGG | 243 | SCCmec III | |
| RIF4 R9 | CGCTTTATCTGTATCTATCGC | |||
| arcC-Up | TTGATTCACCAGCGCGTATTGTC | 456 | Carbamate kinase | [35] |
| arcC-Dn | AGGTATCTGCTTCAATCAGCG | |||
| aroE-Up | ATCGGAAATCCTATTTCACATTC | 456 | Chiquimate dehydrogenase | |
| aroE-Dn | GGTGTTGTATTAATAACGATATC | |||
| aroE-Up | CTAGGAACTGCAATCTTAATCC | 465 | Glycerol kinase | |
| aroE-Dn | TGGTAAAATCGCATGTCCAATTC | |||
| gmk-Up | ATCGTTTTATCGGGACCATC | 429 | Guanylate kinase | |
| gmk-Dn | TCATTAACTACAACGTAATCGTA | |||
| pta-Up | GTTAAAATCGTATTACCTGAAGG | 474 | Phosphate acetyltransferase | |
| pta-Dn | GACCCTTTTGTTGAAAAGCTTAA | |||
| tpi-Up | TCGTTCATTCTGAACGTCGTGAA | 402 | Triose-phosphate isomerase | |
| tpi-Dn | TTTGCACCTTCTAACAATTGTAC | |||
| yqiL-Up | CAGCATACAGGACACCTATTGGC | 516 | Acetylcoenzyme A acetyl transferase | |
| yqiL-Dn | CGTTGAGGAATCGATACTGGAAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cataneli Pereira, V.; Pinheiro-Hubinger, L.; de Oliveira, A.; Moraes Riboli, D.F.; Benini Martins, K.; Calixto Romero, L.; Ribeiro de Souza da Cunha, M.d.L. Detection of the agr System and Resistance to Antimicrobials in Biofilm-Producing S. epidermidis. Molecules 2020, 25, 5715. https://doi.org/10.3390/molecules25235715
Cataneli Pereira V, Pinheiro-Hubinger L, de Oliveira A, Moraes Riboli DF, Benini Martins K, Calixto Romero L, Ribeiro de Souza da Cunha MdL. Detection of the agr System and Resistance to Antimicrobials in Biofilm-Producing S. epidermidis. Molecules. 2020; 25(23):5715. https://doi.org/10.3390/molecules25235715
Chicago/Turabian StyleCataneli Pereira, Valéria, Luiza Pinheiro-Hubinger, Adilson de Oliveira, Danilo Flávio Moraes Riboli, Katheryne Benini Martins, Letícia Calixto Romero, and Maria de Lourdes Ribeiro de Souza da Cunha. 2020. "Detection of the agr System and Resistance to Antimicrobials in Biofilm-Producing S. epidermidis" Molecules 25, no. 23: 5715. https://doi.org/10.3390/molecules25235715
APA StyleCataneli Pereira, V., Pinheiro-Hubinger, L., de Oliveira, A., Moraes Riboli, D. F., Benini Martins, K., Calixto Romero, L., & Ribeiro de Souza da Cunha, M. d. L. (2020). Detection of the agr System and Resistance to Antimicrobials in Biofilm-Producing S. epidermidis. Molecules, 25(23), 5715. https://doi.org/10.3390/molecules25235715