
Saffron: Chemical Composition and Neuroprotective Activity


Abstract
1. Introduction
2. Results
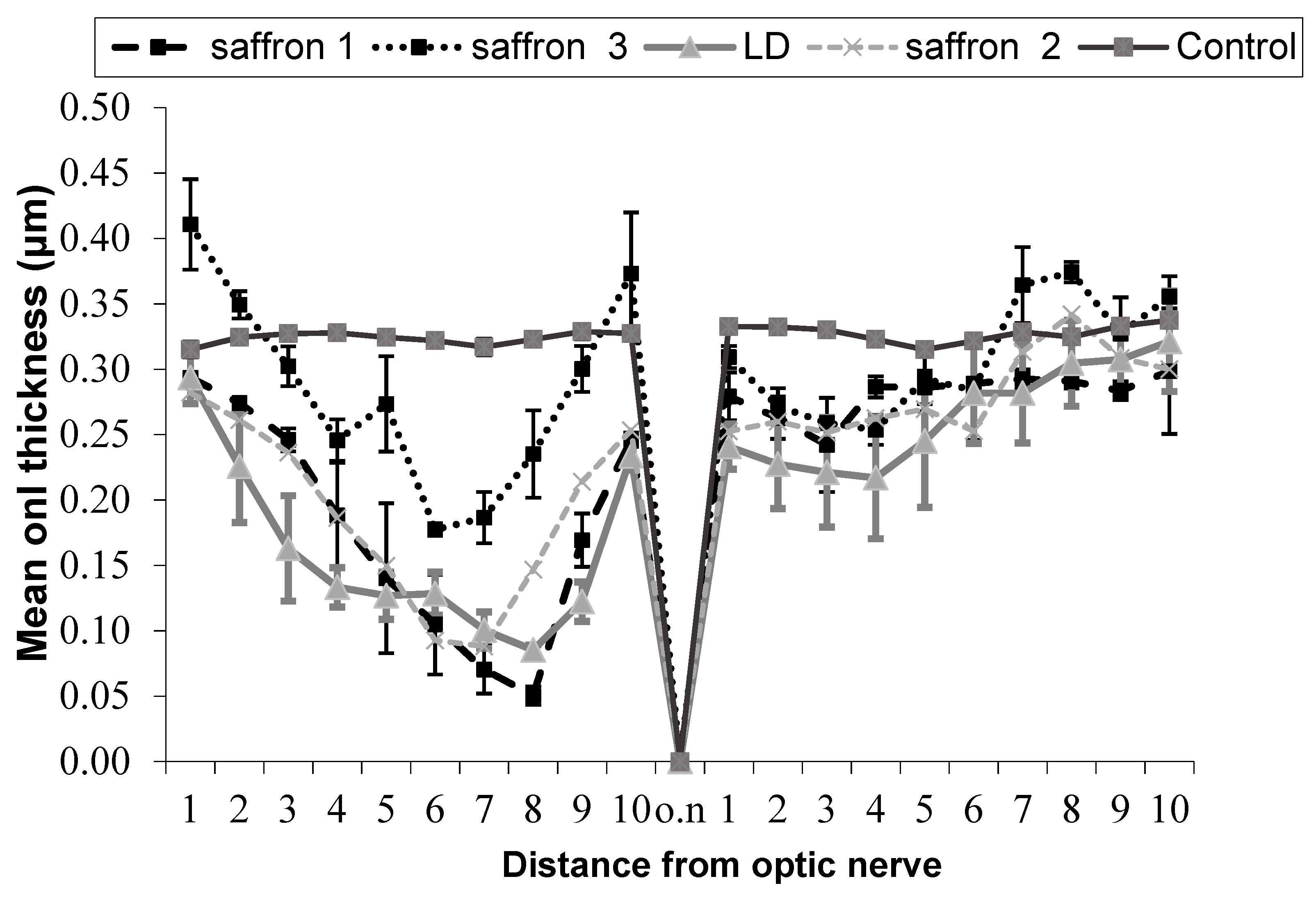
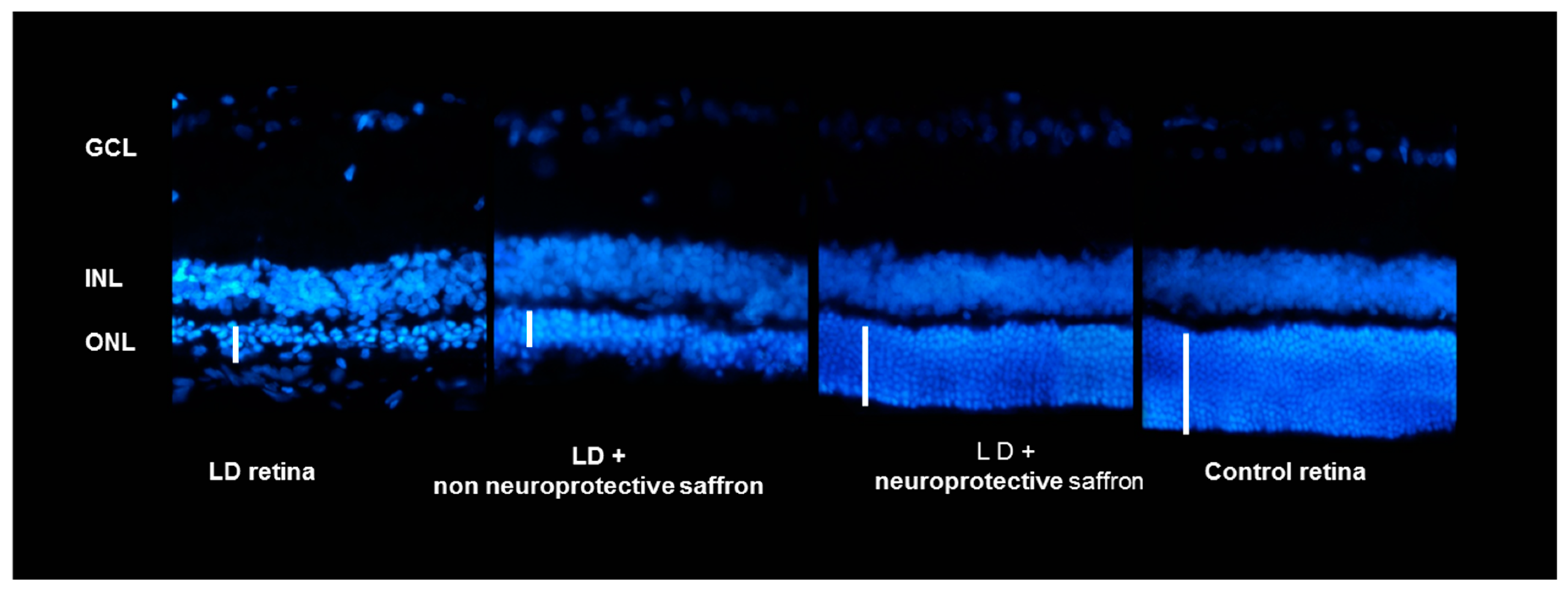
2.1. Correlation between Saffron Chemical Composition and Neuroprotective Activity
- Group saffron 1: Rats treated with saffron having a content of T1 equal to 13% (mg/g) and T2 equal to 5% (mg/g).
- Group saffron 2: Rats treated with saffron having a content of T1 equal to 14% and T2 equal to 5%.
- Group saffron 3: Rats treated with saffron having a content of T1 equal to 17% and T2 equal to 8%.
- Group LD: Rats untreated but subjected to light damage for 24 h (diseased retina).
- Group control: Healthy animals (healthy retina).
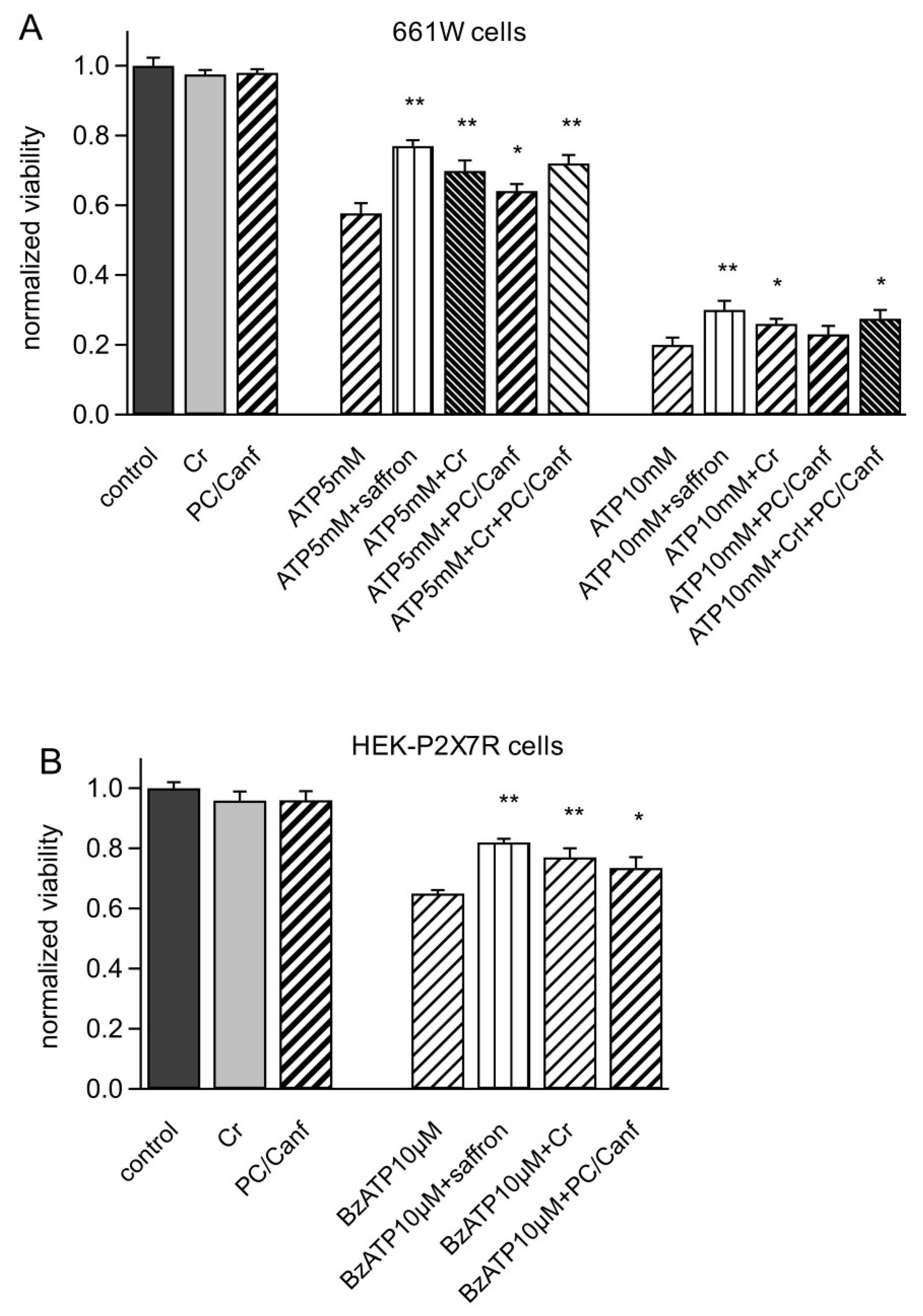
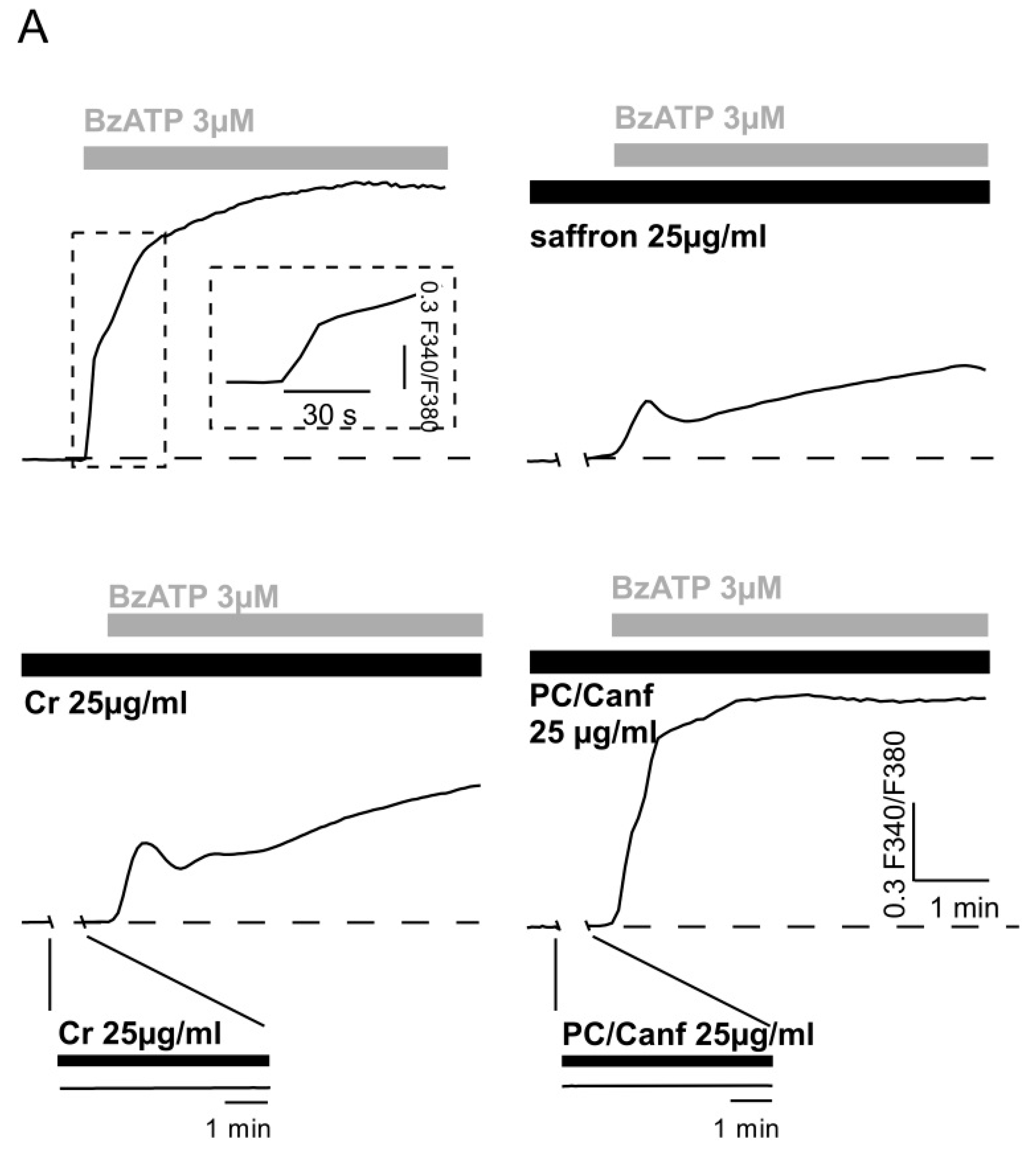
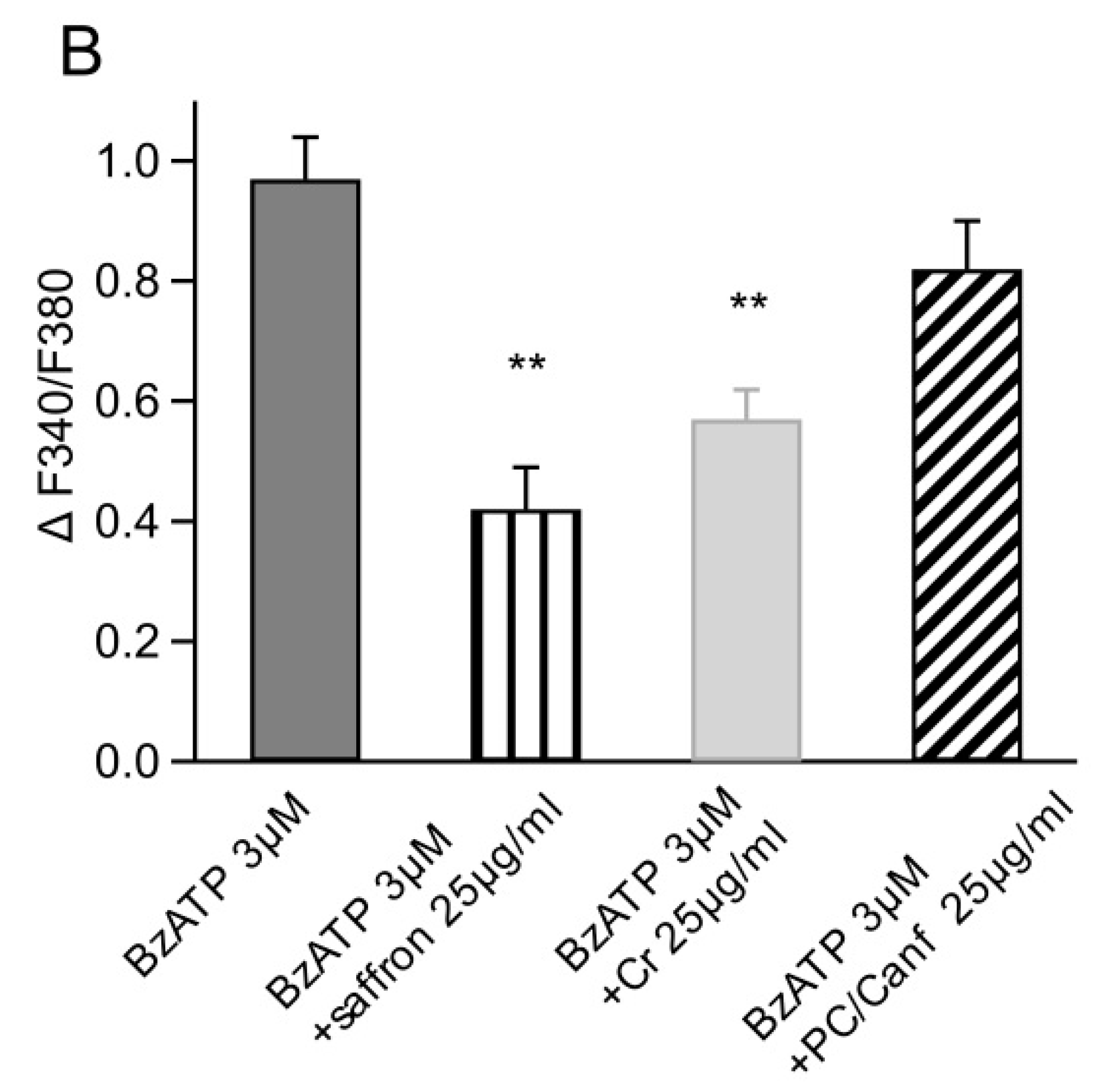
2.2. Saffron Components on Cellular Models
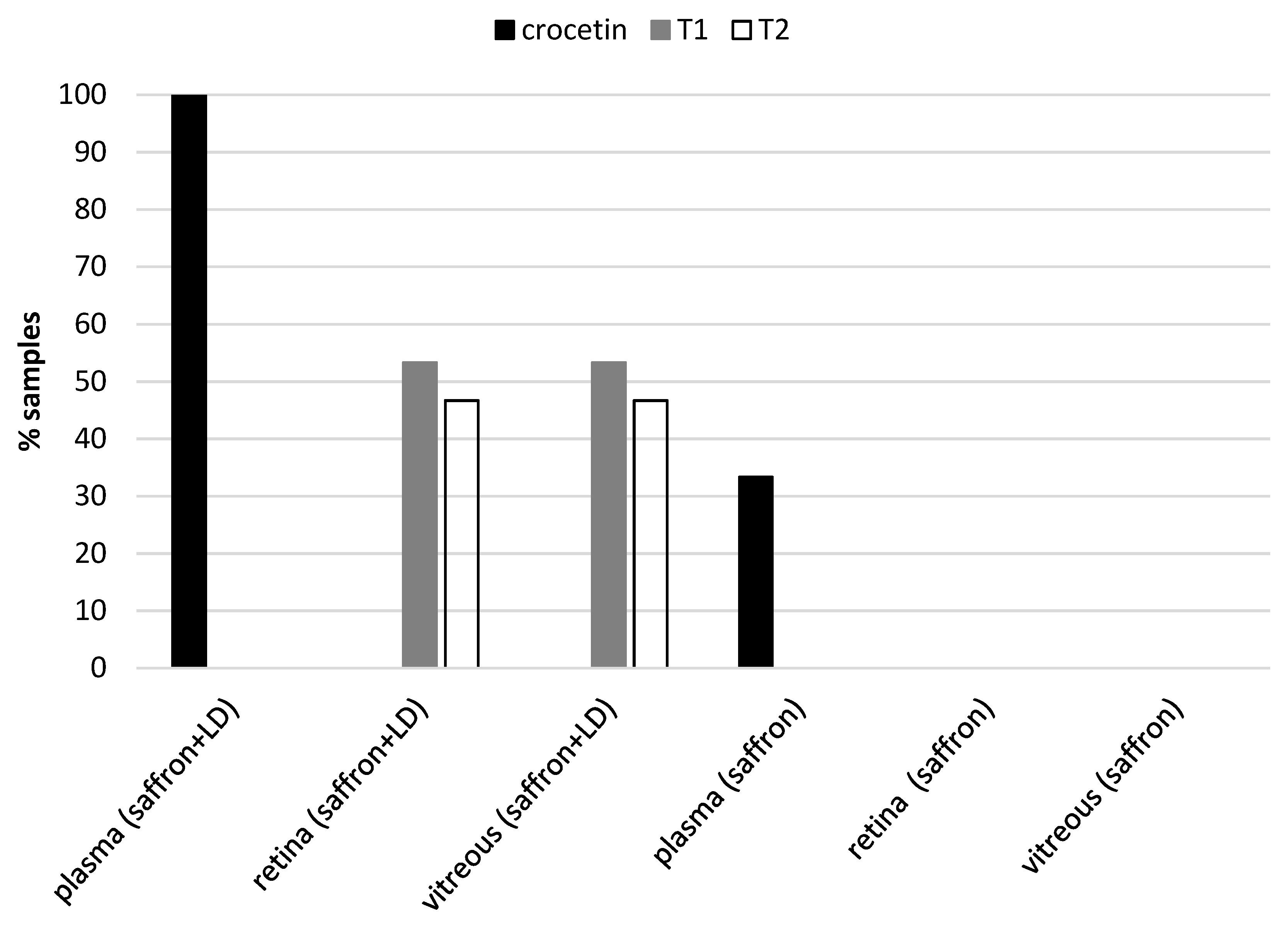
2.3. Metabolites of Saffron in Animal Tissues
3. Discussion
4. Materials and Methods
4.1. Chemical Analysis of Saffron
4.2. Animal Model
4.2.1. Light Damage (LD)
4.2.2. Saffron Treatment
4.2.3. Immunohistochemistry and Morphological Evaluation by Quantitative Histology
4.3. Solid-Phase Extraction (SPE)
- Conditioning: 2 × 5 mL Hexane 2 × 5 mL Methanol (alternating).
- Loading: 15 mL of extract 40 mg recovered in 20 mL H2O.
- Elution A: 2 × 1 mL H2O-EtOH (75:25).
- Elution B: 2 × 1 mL EtOH (100%).
4.4. Cell Cultures
4.4.1. Viability Assay
4.4.2. Intracellular Calcium Measurements
4.5. Tissue Analysis for Saffron Metabolites
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Serrano-Díaz, J.; Sanchez, A.M.; Martínez-Tomé, M.; Winterhalter, P.; Alonso, G.L. Flavonoid Determination in the Quality Control of Floral Bioresidues from Crocus sativus L. J. Agric. Food Chem. 2014, 62, 3125–3133. [Google Scholar] [CrossRef]
- Maccarone, R.; Di Marco, S.; Bisti, S. Saffron Supplement Maintains Morphology and Function after Exposure to Damaging Light in Mammalian Retina. Investig. Opthalmol. Vis. Sci. 2008, 49, 1254–1261. [Google Scholar] [CrossRef]
- Natoli, R.; Zhu, Y.; Valter, K.; Bisti, S.; Eells, J.; Stone, J. Gene and noncoding RNA regulation un-derly-ing photoreceptor protection: Microarray study of dietary antioxidant saffron and photobi-omodulation in rat retina. Mol. Vis. 2010, 16, 1801–1822. [Google Scholar]
- Corso, L.; Cavallero, A.; Baroni, D.; Garbati, P.; Prestipino, G.; Bisti, S.; Nobile, M.; Picco, C. Saffron reduces ATP-induced retinal cytotoxicity by targeting P2X7 receptors. Purinergic Signal. 2016, 12, 161–174. [Google Scholar] [CrossRef]
- Bisti, S.; Maccarone, R.; Falsini, B. Saffron and retina: Neuroprotection and pharmacokinetics. Vis. Neurosci. 2014, 31, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Bisti, S.; Di Marco, S.; Maggi, M.A.; Di Paolo, M.; Piccardi, M.; Falsini, B. Saffron Shifts the Degenera-tive and Inflammatory Phenotype in Photoreceptor Degeneration. In Saffron the Age-Old Panacea in a New Light; Elsevier: Amsterdam, The Netherlands, 2020; Chapter 14. [Google Scholar]
- Maccarone, R.; Rapino, C.; Zerti, D.; Di Tommaso, M.; Battista, N.; Di Marco, S.; Bisti, S.; Maccarrone, M. Modulation of Type-1 and Type-2 Cannabinoid Receptors by Saffron in a Rat Model of Retinal Neurodegeneration. PLoS ONE 2016, 11, e0166827. [Google Scholar] [CrossRef] [PubMed]
- Di Marco, S.; Carnicelli, V.; Franceschini, N.; Di Paolo, M.; Piccardi, M.; Bisti, S.; Falsini, B. Saffron: A Multitask Neuroprotective Agent for Retinal Degenerative Diseases. Antioxidants 2019, 8, 224. [Google Scholar] [CrossRef] [PubMed]
- Piccardi, M.; Fadda, A.; Martelli, F.; Marangoni, D.; Magli, A.; Minnella, A.M.; Bertelli, M.; Di Marco, S.; Bisti, S.; Falsini, B. Antioxidant Saffron and Central Retinal Function in ABCA4-Related Stargardt Macular Dystrophy. Nutrients 2019, 11, 2461. [Google Scholar] [CrossRef] [PubMed]
- Falsini, B.; Piccardi, M.; Minnella, A.; Savastano, M.C.; Capoluongo, E.; Fadda, A.; Balestrazzi, E.; Maccarone, R.; Bisti, S. Influence of Saffron Supplementation on Retinal Flicker Sensitivity in Early Age-Related Macular Degeneration. Investig. Opthalmol. Vis. Sci. 2010, 51, 6118–6124. [Google Scholar] [CrossRef] [PubMed]
- Heitmar, R.; Brown, J.E.; Kyrou, I. Saffron (Crocus sativus L.) in Ocular Diseases: A Narrative Review of the Existing Evidence from Clinical Studies. Nutrients 2019, 11, 649. [Google Scholar] [CrossRef] [PubMed]
- Piccardi, M.; Marangoni, D.; Minnella, A.M.; Savastano, M.C.; Valentini, P.; Ambrosio, L.; Capoluongo, E.D.; Maccarone, R.; Bisti, S.; Falsini, B. A Longitudinal Follow-Up Study of Saffron Supplementation in Early Age-Related Macular Degeneration: Sustained Benefits to Central Retinal Function. Evid. Based Complement. Altern. Med. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Bathaie, S.Z.; Farajzade, A.; Hoshyar, R. A review of the chemistry and uses of crocins and crocetin, the carotenoid natural dyes in saffron, with particular emphasis on applications as colorants including their use as biological stains. Biotech. Histochem. 2014, 89, 401–411. [Google Scholar] [CrossRef]
- Lozano, P.; Castellar, M.; Simancas, M.; Iborra, J.L. A quantitative high-performance liquid chromatographic method to analyse commercial saffron (Crocus sativus L.) products. J. Chromatogr. A 1999, 830, 477–483. [Google Scholar] [CrossRef]
- Rubert, J.; Lacina, O.; Zachariasova, M.; Hajslova, J. Saffron authentication based on liquid chromatography high resolution tandem mass spectrometry and multivariate data analysis. Food Chem. 2016, 204, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Wanrooij, J.; Van Bommel, M.; Quye, A. Characterisation of chemical components for identifying historical Chinese textile dyes by ultra high performance liquid chromatography photodiode array electrospray ionisation mass spectrometer. J. Chromatogr. A 2017, 1479, 87–96. [Google Scholar] [CrossRef]
- Moras, B.; Loffredo, L.; Rey, S. Quality assessment of saffron (Crocus sativus L.) extracts via UHPLC-DAD-MS analysis and detection of adulteration using gardenia fruit extract (Gardenia jasminoides Ellis). Food Chem. 2018, 257, 325–332. [Google Scholar] [CrossRef]
- D’Archivio, A.A.; Di Donato, F.; Foschi, M.; Maggi, M.A.; Ruggieri, F. UHPLC Analysis of Saffron (Crocus sativus L.): Optimization of Separation Using Chemometrics and Detection of Minor Crocetin Esters. Molcules 2018, 23, 1851. [Google Scholar] [CrossRef]
- Bouvier, F.; Suire, C.; Mutterer, J.; Camara, B. Oxidative Remodeling of Chromoplast Carotenoids: Identi-fication of the carotenoid dioxygenase CsCCD and CsZCD genes involved in Crocus secondary metab-olite biogenesis. Plant Cell 2003, 15, 47–62. [Google Scholar] [CrossRef]
- Moraga Ángela, R.; Nohales, P.F.; Pérez, J.A.F.; Gómez-Gómez, L. Glucosylation of the saffron apocarotenoid crocetin by a glucosyltransferase isolated from Crocus sativus stigmas. Planta 2004, 219, 955–966. [Google Scholar] [CrossRef]
- Himeno, H.; Sano, K. Synthesis of Crocin, Picrocrocin and Safranal by Saffron Stigma-like Structures Proliferated in Vitro. Agric. Biol. Chem. 1987, 51, 2395–2400. [Google Scholar] [CrossRef]
- Carmona, M.; Zalacain, A.; Sanchez, A.M.; Novella, A.J.L.; Alonso, G.L. Crocetin Esters, Picrocrocin and Its Related Compounds Present in Crocus sativus Stigmas and Gardenia jasminoides Fruits. Tentative Identification of Seven New Compounds by LC-ESI-M S. J. Agric. Food Chem. 2006, 54, 973–979. [Google Scholar] [CrossRef] [PubMed]
- Cossignani, L.; Urbani, E.; Simonetti, M.S.; Maurizi, A.; Chiesi, C.; Blasi, F. Characterisation of secondary metabolites in saffron from central Italy (Cascia, Umbria). Food Chem. 2014, 143, 446–451. [Google Scholar] [CrossRef] [PubMed]
- Tarantilis, P.A.; Tsoupras, G.; Polissiou, M. Determination of saffron (Crocus sativus L.) components in crude plant extract using high-performance liquid chromatography-UV-visible photodiode-array detection-mass spectrometry. J. Chromatogr. A 1995, 699, 107–118. [Google Scholar] [CrossRef]
- Lech, K.; Witowska-Jarosz, J.; Jarosz, M. Saffron yellow: Characterization of carotenoids by high performance liquid chromatography with electrospray mass spectrometric detection. J. Mass Spectrom. 2009, 44, 1661–1667. [Google Scholar] [CrossRef] [PubMed]
- Koulakiotis, N.S.; Pittenauer, E.; Halabalaki, M.; Tsarbopoulos, A.; Allmaier, G. Comparison of different tandem mass spectrometric techniques (ESI-IT, ESI- and IP-MALDI-QRTOF and vMALDI-TOF/RTOF) for the analysis of crocins and picrocrocin from the stigmas of Crocus sativus L. Rapid Commun. Mass Spectrom. 2012, 26, 670–678. [Google Scholar] [CrossRef]
- Li, N.; Lin, G.; Kwan, Y.W.; Min, Z.-D. Simultaneous quantification of five major biologically active ingredients of saffron by high-performance liquid chromatography. J. Chromatogr. A 1999, 849, 349–355. [Google Scholar] [CrossRef]
- Sanchez, A.M.; Carmona, M.; Zalacain, A.; Carot, J.M.; Jabaloyes, J.M.; Alonso, G.L.; Carmona, M. Rapid Determination of Crocetin Esters and Picrocrocin from Saffron Spice (Crocus sativus L.) Using UV-Visible Spectrophotometry for Quality Control. J. Agric. Food Chem. 2008, 56, 3167–3175. [Google Scholar] [CrossRef]
- ISO 3632-2. Saffron (Crocus sativus L.) Part 2 (Test Methods); International Organization for Stand-Ardiza-Tion Genève: Genève, Switzerland, 2010. [Google Scholar]
- Yamauchi, M.; Tsuruma, K.; Imai, S.; Nakanishi, T.; Umigai, N.; Shimazawa, M.; Hara, H. Crocetin prevents retinal degeneration induced by oxidative and endoplasmic reticulum stresses via inhibition of caspase activity. Eur. J. Pharmacol. 2011, 650, 110–119. [Google Scholar] [CrossRef]
- Ohno, Y.; Nakanishi, T.; Umigai, N.; Tsuruma, K.; Shimazawa, M.; Hara, H. Oral administration of crocetin prevents inner retinal damage induced by N-methyl-d-aspartate in mice. Eur. J. Pharmacol. 2012, 690, 84–89. [Google Scholar] [CrossRef]
- Fernández-Sánchez, L.; Lax, P.; Esquiva, G.; Martín-Nieto, J.; Pinilla, I.; Cuenca, N. Safranal, a Saffron Constituent, Attenuates Retinal Degeneration in P23H Rats. PLoS ONE 2012, 7, e43074. [Google Scholar] [CrossRef]
- Surprenant, A.; Rassendren, F.; Kawashima, E.; North, R.A.; Buell, G. The Cytolytic P2Z Receptor for Extracellular ATP Identified as a P2X Receptor (P2X7). Science 1996, 272, 735–738. [Google Scholar] [CrossRef] [PubMed]
- Giaccio, M. Crocetin from Saffron: An Active Component of an Ancient Spice. Crit. Rev. Food Sci. Nutr. 2004, 44, 155–172. [Google Scholar] [CrossRef] [PubMed]
- Poma, A.; Fontecchio, G.; Carlucci, G.; Chichiricco, G. Anti-Inflammatory Properties of Drugs from saffron crocus. Anti Inflamm. Anti-Allergy Agents Med. Chem. 2012, 11, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Sung, Y.-Y.; Kim, H.K. Crocin Ameliorates Atopic Dermatitis Symptoms by down Regulation of Th2 Response via Blocking of NF-κB/STAT6 Signaling Pathways in Mice. Nutrients 2018, 10, 1625. [Google Scholar] [CrossRef] [PubMed]
- Sperlágh, B.; Vizi, E.S.; Wirkner, K.; Illes, P. P2X7 receptors in the nervous system. Prog. Neurobiol. 2006, 78, 327–346. [Google Scholar] [CrossRef] [PubMed]
- Housley, G.D.; Bringmann, A.; Reichenbach, A. Purinergic signaling in special senses. Trends Neurosci. 2009, 32, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Puthussery, T.; Fletcher, E. Extracellular ATP induces retinal photoreceptor apoptosis through activation of purinoceptors in rodents. J. Comp. Neurol. 2009, 513, 430–440. [Google Scholar] [CrossRef]
- Vessey, K.A.; Jobling, A.; Greferath, U.; Fletcher, E. The Role of the P2X7 Receptor in the Retina: Cell Signalling and Dysfunction. Biol. Mammary Gland 2011, 723, 813–819. [Google Scholar] [CrossRef]
- Zhang, X.; Laties, A.M.; Mitchell, C.H. Stimulation of P2X7 Receptors Elevates Ca2+ and Kills Retinal Ganglion Cells. Investig. Opthalmol. Vis. Sci. 2005, 46, 2183–2191. [Google Scholar] [CrossRef]
- Schmidt, M.V.; Betti, G.; Hensel, A. Saffron in phytotherapy: Pharmacology and clinical uses. Wien. Med. Wochenschr. 2007, 157, 315–319. [Google Scholar] [CrossRef]
- Ayatollahi, H.; Javan, A.O.; Khajedaluee, M.; Shahroodian, M.; Hosseinzadeh, H. Effect of Crocus sativus L. (Saffron) on Coagulation and Anticoagulation Systems in Healthy Volunteers. Phytother. Res. 2013, 28, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Xi, L.; Qian, Z.; Du, P.; Fu, J. Pharmacokinetic properties of crocin (crocetin digentiobiose ester) following oral administration in rats. Phytomedicine 2007, 14, 633–636. [Google Scholar] [CrossRef] [PubMed]
- Asai, A.; Nakano, T.; Takahashi, M.; Nagao, A. Orally Administered Crocetin and Crocins Are Absorbed into Blood Plasma as Crocetin and Its Glucuronide Conjugates in Mice. J. Agric. Food Chem. 2005, 53, 7302–7306. [Google Scholar] [CrossRef] [PubMed]
- Umigai, N.; Murakami, K.; Ulit, M.; Antonio, L.; Shirotori, M.; Morikawa, H.; Nakano, T. The pharmacokinetic profile of crocetin in healthy adult human volunteers after a single oral administration. Phytomedicine 2011, 18, 575–578. [Google Scholar] [CrossRef] [PubMed]
- Kanakis, C.D.; Tarantilis, P.A.; Tajmir-Riahi, H.A.; Polissiou, M.G. Crocetin, Dimethylcrocetin, and Safranal Bind Human Serum Albumin: Stability and Antioxidative Properties. J. Agric. Food Chem. 2007, 55, 970–977. [Google Scholar] [CrossRef] [PubMed]
- Stone, J.; Mitrofanis, J.; Johnstone, D.M.; Falsini, B.; Bisti, S.; Adam, P.; Nuevo, A.B.; George-Weinstein, M.; Mason, R.; Eells, J. Acquired Resilience: An Evolved System of Tissue Protection in Mammals. Dose-Rsponse 2018, 16, 1–40. [Google Scholar] [CrossRef]
- D’Archivio, A.A.; Giannitto, A.; Maggi, M.A.; Ruggieri, F. Geographical classification of Italian saffron (Crocus sativus L.) based on chemical constituents determined by high-performance liquid-chromatography and by using linear discriminant analysis. Food Chem. 2016, 212, 110–116. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Saffron 1 | Saffron 2 | Saffron 3 | Control | Light Damage | |
|---|---|---|---|---|---|
| Saffron 1 | Not significant | p < 0.001 | p < 0.001 | Not significant | |
| Saffron 2 | Not significant | p < 0.001 | p < 0.001 | Not significant | |
| Saffron 3 | p < 0.001 | p < 0.001 | p < 0.005 | p < 0.001 | |
| Control | p < 0.001 | p < 0.001 | p < 0.005 | p < 0.001 | |
| Light damage | Not significant | Not significant | p < 0.001 | p < 0.001 |
Sample Availability: Samples of the compounds are available from the authors. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maggi, M.A.; Bisti, S.; Picco, C. Saffron: Chemical Composition and Neuroprotective Activity. Molecules 2020, 25, 5618. https://doi.org/10.3390/molecules25235618
Maggi MA, Bisti S, Picco C. Saffron: Chemical Composition and Neuroprotective Activity. Molecules. 2020; 25(23):5618. https://doi.org/10.3390/molecules25235618
Chicago/Turabian StyleMaggi, Maria Anna, Silvia Bisti, and Cristiana Picco. 2020. "Saffron: Chemical Composition and Neuroprotective Activity" Molecules 25, no. 23: 5618. https://doi.org/10.3390/molecules25235618
APA StyleMaggi, M. A., Bisti, S., & Picco, C. (2020). Saffron: Chemical Composition and Neuroprotective Activity. Molecules, 25(23), 5618. https://doi.org/10.3390/molecules25235618