
Chili Pepper Carotenoids: Nutraceutical Properties and Mechanisms of Action



Abstract
1. Introduction
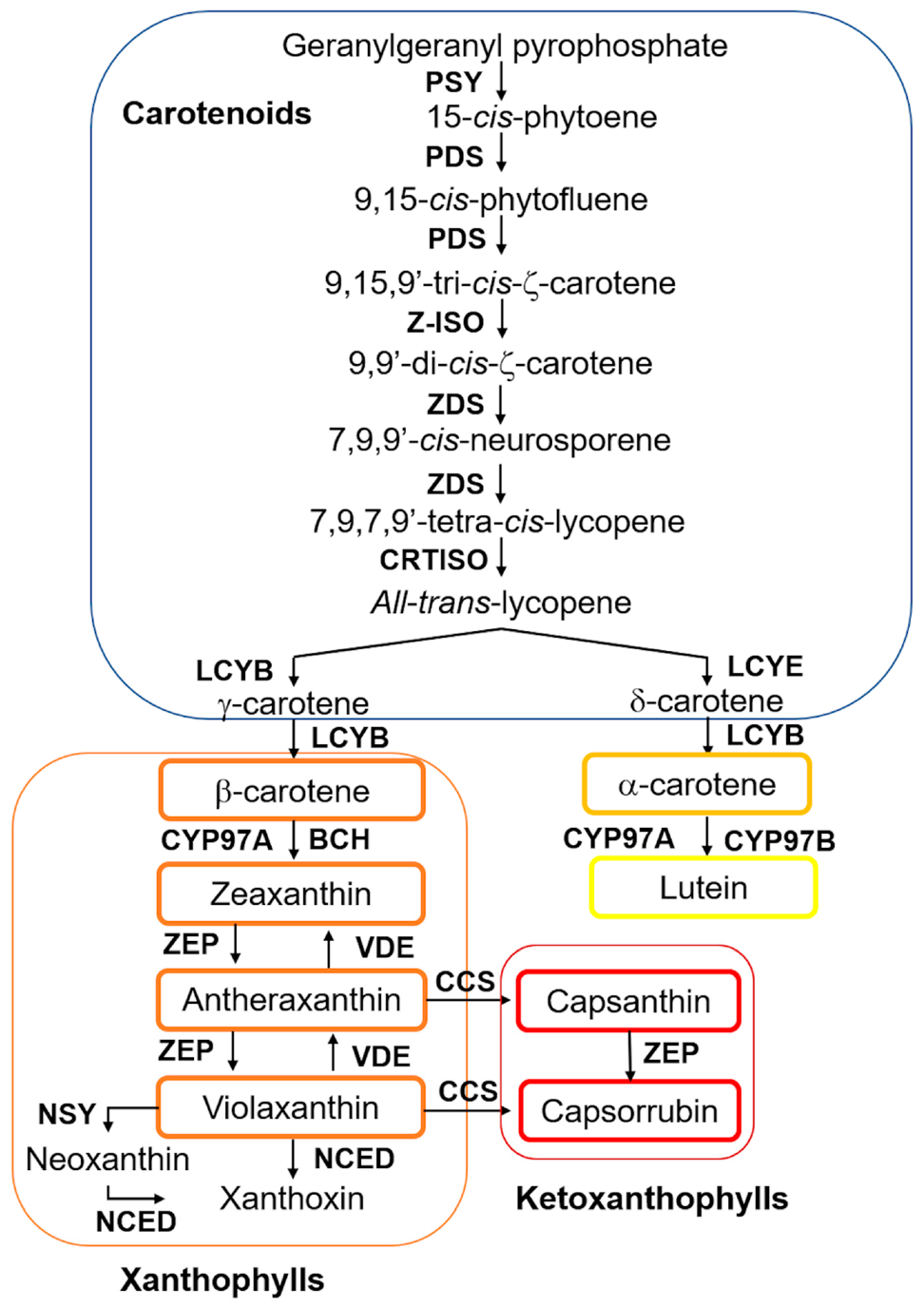
2. Biosynthetic Pathway of Carotenoids in Chili Pepper Fruits
3. Carotenoid Types and Contents in Chili Pepper Fruits
4. Effects of Processing on the Carotenoid Content
5. Carotenoid Fate after Ingestion
6. Carotenoids as Antioxidants
7. Carotenoids and Cancer
8. Carotenoids as Anti-Inflammatory Agents
9. Carotenoids and Obesity
10. Miscellaneous Effects
11. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. Crops; FAO: Roma, Italy, 2018. [Google Scholar]
- Tripodi, P.; Kumar, S. The Capsicum Crop: An introduction. In The Capsicum Genome; Ramchiary, N., Kole, C., Eds.; Springer International Publishing: Basel, Switzerland, 2019. [Google Scholar] [CrossRef]
- Jarret, R.L.; Barboza, G.E.; Costa Batista, F.R.d.; Berke, T.; Chou, Y.-Y.; Hulse-Kemp, A.; Ochoa-Alejo, N.; Tripodi, P.; Veres, A.; Garcia, C.C.; et al. Capsicum—An abbreviated compendium. J. Am. Soc. Hort. Sci. 2019, 144, 3–22. [Google Scholar] [CrossRef]
- Carrizo Garcia, C.; Barfuss, M.H.; Sehr, E.M.; Barboza, G.E.; Samuel, R.; Moscone, E.A.; Ehrendorfer, F. Phylogenetic relationships, diversification and expansion of chili peppers (Capsicum, Solanaceae). Ann. Bot. 2016, 118, 35–51. [Google Scholar] [CrossRef] [PubMed]
- Antonio, A.S.; Wiedemann, L.S.M.; Veiga Junior, V.F. The genus Capsicum: A phytochemical review of bioactive secondary metabolites. RSC Adv. 2018, 8, 25767–25784. [Google Scholar] [CrossRef]
- Baenas, N.; Belovic, M.; Ilic, N.; Moreno, D.A.; Garcia-Viguera, C. Industrial use of pepper (Capsicum annum L.) derived products: Technological benefits and biological advantages. Food Chem. 2019, 274, 872–885. [Google Scholar] [CrossRef] [PubMed]
- Batiha, G.E.; Alqahtani, A.; Ojo, O.A.; Shaheen, H.M.; Wasef, L.; Elzeiny, M.; Ismail, M.; Shalaby, M.; Murata, T.; Zaragoza-Bastida, A.; et al. Biological properties, bioactive constituents, and pharmacokinetics of some Capsicum spp. and capsaicinoids. Int. J. Mol. Sci. 2020, 21, 5179. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.V.; de Andrade Mattietto, R.; de Oliveira Rios, A.; de Almeida Maciel, R.; Moresco, K.S.; de Souza Oliveira, T.C. Bioactive compounds and antioxidant activity of pepper (Capsicum sp.) genotypes. J. Food Sci. Technol. 2015, 52, 7457–7464. [Google Scholar] [CrossRef]
- Badia, A.D.; Spina, A.A.; Vassalotti, G. Capsicum annuum L.: An overview of biological activities and potential nutraceutical properties in humans and animals. J. Nutr. Ecol. Food Res. 2017, 4, 167–177. [Google Scholar] [CrossRef]
- Morales-Soriano, E.; Panozzo, A.; Ugás, R.; Grauwet, T.; Van Loey, A.; Hendrickx, M. Carotenoid profile and basic structural indicators of native Peruvian chili peppers. Eur. Food Res. Technol. 2018, 245, 717–732. [Google Scholar] [CrossRef]
- Alcaíno, J.; Baeza, M.; Cifuentes, V. Carotenoid distribution in nature. In Carotenoids in Nature. Biosynthesis, Regulation and Function; Stange, C., Ed.; Springer: Cham, Seitzerland, 2016; Volume 79, pp. 3–33. [Google Scholar]
- Moise, A.R.; Al-Babili, S.; Wurtzel, E.T. Mechanistic aspects of carotenoid biosynthesis. Chem. Rev. 2014, 114, 164–193. [Google Scholar] [CrossRef]
- Rodríguez-Concepción, M.; Avalos, J.; Bonet, M.L.; Boronat, A.; Gómez-Gómez, L.; Hornero-Méndez, D.; Limon, M.C.; Melendez-Martínez, A.J.; Olmedilla-Alonso, B.; Palou, A.; et al. A global perspective on carotenoids: Metabolism, biotechnology, and benefits for nutrition and health. Prog. Lipid Res. 2018, 70, 62–93. [Google Scholar] [CrossRef]
- Maoka, T. Carotenoids as natural functional pigments. J. Nat. Med. 2020, 74, 1–16. [Google Scholar] [CrossRef]
- Kim, J.H.; Lee, J.; Choi, I.J.; Kim, Y.I.; Kwon, O.; Kim, H.; Kim, J. Dietary carotenoids intake and the risk of gastric cancer: A case-control study in Korea. Nutrients 2018, 10, 1031. [Google Scholar] [CrossRef] [PubMed]
- Molnar, J.; Gyemant, N.; Mucsi, I.; Molnar, A.; Szabo, M.; Kortvelyesi, T.; Varga, A.; Molnar, P.; Toth, G. Modulation of multidrug resistance and apoptosis of cancer cells by selected carotenoids. In Vivo 2004, 18, 237–244. [Google Scholar] [PubMed]
- Boyko, Y.; Kravchenko, I.; Shandra, A.; Boiko, I. Extraction, identification and anti-inflammatory activity of carotenoids out of Capsicum Anuum L. J. Herbmed Pharmacol. 2017, 6, 10–15. [Google Scholar]
- Tuzcu, M.; Orhan, C.; Muz, O.E.; Sahin, N.; Juturu, V.; Sahin, K. Lutein and zeaxanthin isomers modulates lipid metabolism and the inflammatory state of retina in obesity-induced high-fat diet rodent model. BMC Ophthalmol. 2017, 17, 129. [Google Scholar] [CrossRef] [PubMed]
- Nwachukwu, I.D.; Udenigwe, C.C.; Aluko, R.E. Lutein and zeaxanthin: Production technology, bioavailability, mechanisms of action, visual function, and health claim status. Trends Food Sci. Technol. 2016, 49, 74–84. [Google Scholar] [CrossRef]
- Antwi, S.O.; Steck, S.E.; Su, L.J.; Hebert, J.R.; Zhang, H.; Craft, N.E.; Fontham, E.T.; Smith, G.J.; Bensen, J.T.; Mohler, J.L.; et al. Carotenoid intake and adipose tissue carotenoid levels in relation to prostate cancer aggressiveness among African-American and European-American men in the North Carolina-Louisiana prostate cancer project (PCaP). Prostate 2016, 76, 1053–1066. [Google Scholar] [CrossRef]
- Rowles, J.L., 3rd; Erdman, J.W., Jr. Carotenoids and their role in cancer prevention. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158613. [Google Scholar] [CrossRef]
- Hernández-Ortega, M.; Ortiz-Moreno, A.; Hernandez-Navarro, M.D.; Chamorro-Cevallos, G.; Dorantes-Alvarez, L.; Necoechea-Mondragon, H. Antioxidant, antinociceptive, and anti-inflammatory effects of carotenoids extracted from dried pepper (Capsicum annuum L.). J. Biomed. Biotechnol. 2012, 2012, 524019. [Google Scholar] [CrossRef]
- Coronel, J.; Pinos, I.; Amengual, J. Beta-carotene in obesity research: Technical considerations and current status of the field. Nutrients 2019, 11, 842. [Google Scholar] [CrossRef]
- Lim, J.Y.; Wang, X.D. Mechanistic understanding of beta-cryptoxanthin and lycopene in cancer prevention in animal models. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158652. [Google Scholar] [CrossRef] [PubMed]
- Mounien, L.; Tourniaire, F.; Landrier, J.F. Anti-obesity effect of carotenoids: Direct impact on adipose tissue and adipose tissue-driven indirect effects. Nutrients 2019, 11, 1562. [Google Scholar] [CrossRef]
- Thuphairo, K.; Sornchan, P.; Suttisansanee, U. Bioactive compounds, antioxidant activity and inhibition of key enzymes relevant to Alzheimer’s disease from sweet pepper (Capsicum annuum) extracts. Prev. Nutr. Food Sci. 2019, 24, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Narisawa, T.; Fukaura, Y.; Hasebe, M.; Nomura, S.; Oshima, S.; Inakuma, T. Prevention of N-methylnitrosourea-induced colon carcinogenesis in rats by oxygenated carotenoid capsanthin and capsanthin-rich paprika juice. Proc. Soc. Exp. Biol. Med. 2000, 224, 116–122. [Google Scholar] [CrossRef]
- Maoka, T.; Mochida, K.; Kozuka, M.; Ito, Y.; Fujiwara, Y.; Hashimoto, K.; Enjo, F.; Ogata, M.; Nobukuni, Y.; Tokuda, H.; et al. Cancer chemopreventive activity of carotenoids in the fruits of red paprika Capsicum annuum L. Cancer Lett. 2001, 172, 103–109. [Google Scholar] [CrossRef]
- Erden, Y. Capsanthin stimulates the mitochondrial apoptosis-mediated cell ceath, following DNA damage in MCF-7 cells. Nutr. Cancer 2020. [Google Scholar] [CrossRef]
- Aizawa, K.; Inakuma, T. Dietary capsanthin, the main carotenoid in paprika (Capsicum annuum), alters plasma high-density lipoprotein-cholesterol levels and hepatic gene expression in rats. Br. J. Nutr. 2009, 102, 1760–1766. [Google Scholar] [CrossRef]
- Kim, J.S.; Ha, T.Y.; Kim, S.; Lee, S.J.; Ahn, J. Red paprika (Capsicum annuum L.) and its main carotenoid capsanthin ameliorate impaired lipid metabolism in the liver and adipose tissue of high-fat diet-induced obese mice. J. Funct. Foods 2017, 31, 131–140. [Google Scholar] [CrossRef]
- Jo, S.J.; Kim, J.W.; Choi, H.O.; Kim, J.H.; Kim, H.J.; Woo, S.H.; Han, B.H. Capsanthin inhibits both adipogenesis in 3T3-L1 preadipocytes and weight gain in high-fat diet-induced obese mice. Biomol. Ther. 2017, 25, 329–336. [Google Scholar] [CrossRef]
- Wu, T.; Gao, Y.; Hao, J.; Geng, J.; Zhang, J.; Yin, J.; Liu, R.; Sui, W.; Gong, L.; Zhang, M. Capsanthin extract prevents obesity, reduces serum TMAO levels and modulates the gut microbiota composition in high-fat-diet induced obese C57BL/6J mice. Food Res. Int. 2020, 128, 108774. [Google Scholar] [CrossRef]
- Fernández-Bedmar, Z.; Alonso-Moraga, A. In vivo and in vitro evaluation for nutraceutical purposes of capsaicin, capsanthin, lutein and four pepper varieties. Food Chem. Toxicol. 2016, 98, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Gómez-García, M.R.; Ochoa-Alejo, N. Biochemistry and molecular biology of carotenoid biosynthesis in chili peppers (Capsicum spp.). Int. J. Mol. Sci. 2013, 14, 19025–19053. [Google Scholar] [CrossRef] [PubMed]
- Sathasivam, R.; Radhakrishnan, R.; Kim, J.K.; Park, S.U. An update on biosynthesis and regulation of carotenoids in plants. S. Afr. J. Bot. 2020. [Google Scholar] [CrossRef]
- Bouvier, F.; Hugueney, P.; d’Harlingue, A.; Kuntz, M.; Camara, B. Xanthophyll biosynthesis in chromoplasts: Isolation and molecular cloning of an enzyme catalyzing the conversion of 5,6-epoxycarotenoid into ketocarotenoid. Plant J. 1994, 6, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Ha, S.-H.; Lee, S.-W.; Kim, J.-G.; Hwang, Y.-S. Expression patterns of genes involved in carotenoid biosynthesis in pepper. J. Appl. Biol. Chem. 1999, 42, 92–96. [Google Scholar]
- Martínez-López, L.A.; Ochoa-Alejo, N.; Martínez, O. Dynamics of the chili pepper transcriptome during fruit development. BMC Genom. 2014, 15, 143. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.L.; Li, L.; Shah, S.N.M.; Gong, Z.H. The relationship between red fruit colour formation and key genes of capsanthin biosynthesis pathway in Capsicum annuum. Biol. Plant. 2015, 59, 507–513. [Google Scholar] [CrossRef]
- Wang, Q.; Cao, T.J.; Zheng, H.; Zhou, C.F.; Wang, Z.; Wang, R.; Lu, S. Manipulation of carotenoid metabolic flux by lycopene cyclization in ripening red pepper ( Capsicum annuum var. conoides) fruits. J. Agric. Food Chem. 2019, 67, 4300–4310. [Google Scholar] [CrossRef]
- Liu, Y.; Lv, J.; Liu, Z.; Wang, J.; Yang, B.; Chen, W.; Ou, L.; Dai, X.; Zhang, Z.; Zou, X. Integrative analysis of metabolome and transcriptome reveals the mechanism of color formation in pepper fruit (Capsicum annuum L.). Food Chem. 2020, 306, 125629. [Google Scholar] [CrossRef]
- Wang, Q.; Huang, X.Q.; Cao, T.J.; Zhuang, Z.; Wang, R.; Lu, S. Heteromeric geranylgeranyl diphosphate synthase contributes to carotenoid biosynthesis in ripening fruits of red pepper (Capsicum annuum var. conoides). J. Agric. Food Chem. 2018, 66, 11691–11700. [Google Scholar] [CrossRef]
- Simkin, A.J.; Zhu, C.; Kuntz, M.; Sandmann, G. Light-dark regulation of carotenoid biosynthesis in pepper (Capsicum annuum) leaves. J. Plant Physiol. 2003, 160, 439–443. [Google Scholar] [CrossRef]
- Bian, W.; Barsan, C.; Egea, I.; Purgatto, E.; Chervin, C.; Zouine, M.; Latché, A.; Bouzayen, M.; Pech, J.-C. Metabolic and molecular events occurring during chromoplast biogenesis. J. Bot. 2011, 2011, 289859. [Google Scholar] [CrossRef][Green Version]
- Sadali, N.M.; Sowden, R.G.; Ling, Q.; Jarvis, R.P. Differentiation of chromoplasts and other plastids in plants. Plant Cell Rep. 2019, 38, 803–818. [Google Scholar] [CrossRef]
- Sun, T.; Yuan, H.; Cao, H.; Yazdani, M.; Tadmor, Y.; Li, L. Carotenoid metabolism in plants: The role of plastids. Mol. Plant 2018, 11, 58–74. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Q.; Yang, Y.; Fei, Z.; Yuan, H.; Fish, T.; Thannhauser, T.W.; Mazourek, M.; Kochian, L.V.; Wang, X.; Li, L. Proteomic analysis of chromoplasts from six crop species reveals insights into chromoplast function and development. J. Exp. Bot. 2013, 64, 949–961. [Google Scholar] [CrossRef]
- Ytterberg, A.J.; Peltier, J.B.; van Wijk, K.J. Protein profiling of plastoglobules in chloroplasts and chromoplasts. A surprising site for differential accumulation of metabolic enzymes. Plant Physiol. 2006, 140, 984–997. [Google Scholar] [CrossRef] [PubMed]
- Shumskaya, M.; Wurtzel, E.T. The carotenoid biosynthetic pathway: Thinking in all dimensions. Plant Sci. 2013, 208, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Piano, D.; Cocco, E.; Guadalupi, G.; Kalaji, H.M.; Kirkpatrick, J.; Farci, D. Characterization under quasi-native conditions of the capsanthin/capsorubin synthase from Capsicum annuum L. Plant Physiol. Biochem. 2019, 143, 165–175. [Google Scholar] [CrossRef]
- Hornero-Méndez, D.; Gómez-Ladrón De Guevara, R.; Mínguez-Mosquera, M.I. Carotenoid biosynthesis changes in five red pepper (Capsicum annuum L.) cultivars during ripening. Cultivar selection for breeding. J. Agric. Food Chem. 2000, 48, 3857–3864. [Google Scholar] [CrossRef]
- Jeong, H.B.; Kang, M.Y.; Jung, A.; Han, K.; Lee, J.H.; Jo, J.; Lee, H.Y.; An, J.W.; Kim, S.; Kang, B.C. Single-molecule real-time sequencing reveals diverse allelic variations in carotenoid biosynthetic genes in pepper (Capsicum spp.). Plant Biotechnol. J. 2019, 17, 1081–1093. [Google Scholar] [CrossRef]
- Kilcrease, J.; Rodriguez-Uribe, L.; Richins, R.D.; Arcos, J.M.; Victorino, J.; O’Connell, M.A. Correlations of carotenoid content and transcript abundances for fibrillin and carotenogenic enzymes in Capsicum annum fruit pericarp. Plant Sci. 2015, 232, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Berry, H.M.; Rickett, D.V.; Baxter, C.J.; Enfissi, E.M.A.; Fraser, P.D. Carotenoid biosynthesis and sequestration in red chilli pepper fruit and its impact on colour intensity traits. J. Exp. Bot. 2019, 70, 2637–2650. [Google Scholar] [CrossRef] [PubMed]
- Wahyuni, Y.; Ballester, A.R.; Sudarmonowati, E.; Bino, R.J.; Bovy, A.G. Metabolite biodiversity in pepper (Capsicum) fruits of thirty-two diverse accessions: Variation in health-related compounds and implications for breeding. Phytochemistry 2011, 72, 1358–1370. [Google Scholar] [CrossRef]
- Mohd Hassan, N.; Yusof, N.A.; Yahaya, A.F.; Mohd Rozali, N.N.; Othman, R. Carotenoids of capsicum fruits: Pigment profile and health-promoting functional attributes. Antioxidants 2019, 8, 469. [Google Scholar] [CrossRef] [PubMed]
- Biacs, P.A.; Daood, H.G.; Pavisa, A.; Hajdu, F. Studies on the carotenoid pigments of paprika (Capsicum annuum L.var Sz-20). J. Agric. Food Chem. 1989, 37, 350–353. [Google Scholar] [CrossRef]
- Deli, J.; Molnar, P.; Matus, Z.; Toth, G. Carotenoid composition in the fruits of red paprika (Capsicum annuum var. lycopersiciforme rubrum) during ripening; biosynthesis of carotenoids in red paprika. J. Agric. Food Chem. 2001, 49, 1517–1523. [Google Scholar] [CrossRef]
- Kim, J.S.; An, C.G.; Park, J.S.; Lim, Y.P.; Kim, S. Carotenoid profiling from 27 types of paprika (Capsicum annuum L.) with different colors, shapes, and cultivation methods. Food Chem. 2016, 201, 64–71. [Google Scholar] [CrossRef]
- Guzman, I.; Hamby, S.; Romero, J.; Bosland, P.W.; O’Connell, M.A. Variability of carotenoid biosynthesis in orange colored Capsicum spp. Plant Sci. 2010, 179, 49–59. [Google Scholar] [CrossRef]
- Rodríguez-Uribe, L.; Hernández, L.; Kilcrease, J.P.; Walker, S.; O’Connell, M.A. Capsaicinoid and carotenoid composition and genetic diversity of Kas I and Ccs in New Mexican Capsicum annuum L. Landraces. HortScience 2014, 49, 1370–1375. [Google Scholar] [CrossRef]
- Rodríguez-Uribe, L.; Guzmán, I.; Rajapakse, W.; Richins, R.D.; O’Connell, M.A. Carotenoid accumulation in orange-pigmented Capsicum annuum fruit, regulated at multiple levels. J. Exp. Bot. 2011, 63, 517–526. [Google Scholar] [CrossRef]
- Borovsky, Y.; Tadmor, Y.; Bar, E.; Meir, A.; Lewinsohn, E.; Paran, I. Induced mutation in beta-carotene hidroxylase results in accumulation of beta-carotene and conversion of red to orange color in pepper fruit. Theor. Appl. Genet. 2013, 126, 557–565. [Google Scholar] [CrossRef] [PubMed]
- da Silveira Agostini-Costa, T.; da Silva Gomes, I.; de Melo, L.A.M.P.; Reifschneider, F.J.B.; da Costa Ribeiro, C.S. Carotenoid and total vitamin C content of peppers from selected Brazilian cultivars. J. Food Compost. Anal. 2017, 57, 73–79. [Google Scholar] [CrossRef]
- Jang, S.-J.; Jeong, H.-B.; Jung, A.; Kang, M.-Y.; Kim, S.; Ha, S.-H.; Kwon, J.-K.; Kang, B.-C. Phytoene synthase 2 can compensate for the absence of PSY1 in the control of color in Capsicum fruit. J. Exp. Bot. 2020, 71, 3417–3427. [Google Scholar] [CrossRef] [PubMed]
- Ha, S.-H.; Kim, J.-B.; Park, J.-S.; Lee, S.-W.; Cho, K.-J. A comparison of the carotenoid accumulation in Capsicum varieties that show different ripening colours: Deletion of the capsanthin-capsorubin synthase gene is not a prerequisite for the formation of a yellow pepper. J. Exp. Bot. 2007, 58, 3135–3144. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, S.; Gui, X.L.; Chang, X.B.; Gong, Z.H. A further analysis of the relationship between yellow ripe-fruit color and the capsanthin-capsorubin synthase gene in pepper (Capsicum sp.) indicated a new mutant variant in C. annuum and a tandem repeat structure in promoter region. PLoS ONE 2013, 8, e61996. [Google Scholar] [CrossRef]
- Adadi, P.; Barakova, N.V.; Krivoshapkina, E.F. Selected methods of extracting carotenoids, characterization, and health concerns: A review. J. Agric. Food. Chem. 2018, 66, 5925–5947. [Google Scholar] [CrossRef]
- Pérez-Galvez, A.; Minguez-Mosquera, M.I. Degradation of non-esterified and esterified xanthophylls by free radicals. Biochim. Biophys. Acta 2002, 1569, 31–34. [Google Scholar] [CrossRef]
- Al-Juhaimi, F.; Ghafoor, K.; Ozcan, M.M.; Jahurul, M.H.A.; Babiker, E.E.; Jinap, S.; Sahena, F.; Sharifudin, M.S.; Zaidul, I.S.M. Effect of various food processing and handling methods on preservation of natural antioxidants in fruits and vegetables. J. Food Sci. Technol. 2018, 55, 3872–3880. [Google Scholar] [CrossRef] [PubMed]
- Giuffrida, D.; Dugo, P.; Torre, G.; Bignardi, C.; Cavazza, A.; Corradini, C.; Dugo, G. Evaluation of carotenoid and capsaicinoid contents in powder of red chili peppers during one year of storage. Food Res. Int. 2014, 65, 163–170. [Google Scholar] [CrossRef]
- Pola, W.; Sugaya, S.; Photchanachai, S. Influence of postharvest temperatures on carotenoid biosynthesis and phytochemicals in mature green chili (Capsicum annuum L.). Antioxidants 2020, 9, 203. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Ambrocio, A.; Guerrero-Beltrán, J.A.; Aparicio-Fernández, X.; Ávila-Sosa, R.; Hernández-Carranza, P.; Cid-Pérez, S.; Ochoa-Velasco, C.E. Effect of blue and ultraviolet-C light irradiation on bioactive compounds and antioxidant capacity of habanero pepper (Capsicum chinense) during refrigeration storage. Postharvest Biol. Technol. 2018, 135, 19–26. [Google Scholar] [CrossRef]
- Pola, W.; Sugaya, S.; Photchanachai, S. Color development and phytochemical changes in mature green chili (Capsicum annuum L.) exposed to red and blue light-emitting diodes. J. Agric. Food. Chem. 2020, 68, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Bains, K. Traditional cooked vegetable dishes as important sources of ascorbic acid and beta-carotene in the diets of Indian urban and rural families. Food Nutr. Bull. 2006, 27, 306–310. [Google Scholar] [CrossRef] [PubMed]
- Gunathilake, K.; Ranaweera, K.; Rupasinghe, H.P.V. Effect of different cooking methods on polyphenols, carotenoids and antioxidant activities of selected edible leaves. Antioxidants 2018, 7, 117. [Google Scholar] [CrossRef] [PubMed]
- Loizzo, M.R.; Pugliese, A.; Bonesi, M.; De Luca, D.; O’Brien, N.; Menichini, F.; Tundis, R. Influence of drying and cooking process on the phytochemical content, antioxidant and hypoglycaemic properties of two bell Capsicum annum L. cultivars. Food Chem. Toxicol. 2013, 53, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Pugliese, A.; O’Callaghan, Y.; Tundis, R.; Galvin, K.; Menichini, F.; O’Brien, N.; Loizzo, M.R. In vitro investigation of the bioaccessibility of carotenoids from raw, frozen and boiled red chili peppers (Capsicum annuum). Eur. J. Nutr. 2014, 53, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Pugliese, A.; Loizzo, M.R.; Tundis, R.; O’Callaghan, Y.; Galvin, K.; Menichini, F.; O’Brien, N. The effect of domestic processing on the content and bioaccessibility of carotenoids from chili peppers (Capsicum species). Food Chem. 2013, 141, 2606–2613. [Google Scholar] [CrossRef]
- Brasili, E.; Chaves, D.F.; Xavier, A.A.; Mercadante, A.Z.; Hassimotto, N.M.; Lajolo, F.M. Effect of pasteurization on flavonoids and carotenoids in Citrus sinensis (L.) Osbeck cv. ‘Cara Cara’ and ‘Bahia’ juices. J. Agric. Food Chem. 2017, 65, 1371–1377. [Google Scholar] [CrossRef]
- Kevrešan, Ž.S.; Mandić, A.P.; Kuhajda, K.N.; Sakač, M.B. Carotenoid content in fresh and dry pepper (Capsicum annuum L.) fruits for paprika production. Food Feed Res. 2009, 36, 21–28. [Google Scholar]
- Campos-Hernández, N.; Jaramillo-Flores, M.E.; Téllez-Medina, D.I.; Alamilla-Beltrán, L. Effect of traditional dehydration processing of pepper jalapeno rayado (Capsicum annuum) on secondary metabolites with antioxidant activity. CyTA J. Food 2018, 16, 316–324. [Google Scholar] [CrossRef]
- Montoya-Ballesteros, L.C.; González-León, A.; García-Alvarado, M.A.; Rodríguez-Jimenes, G.C. Bioactive compounds during drying of chili peppers. Dry. Technol. 2014, 32, 1486–1499. [Google Scholar] [CrossRef]
- Sanatombi, K.; Rajkumari, S. Effect of processing on quality of pepper: A review. Food Rev. Int. 2019, 36, 626–643. [Google Scholar] [CrossRef]
- Andres, V.; Villanueva, M.J.; Tenorio, M.D. The effect of high-pressure processing on colour, bioactive compounds, and antioxidant activity in smoothies during refrigerated storage. Food Chem. 2016, 192, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Cilla, A.; Bosch, L.; Barberá, R.; Alegría, A. Effect of processing on the bioaccessibility of bioactive compounds—A review focusing on carotenoids, minerals, ascorbic acid, tocopherols and polyphenols. J. Food Compost. Anal. 2018, 68, 3–15. [Google Scholar] [CrossRef]
- Eun, J.B.; Maruf, A.; Das, P.R.; Nam, S.H. A review of encapsulation of carotenoids using spray drying and freeze drying. Crit. Rev. Food Sci. Nutr. 2019. [Google Scholar] [CrossRef]
- Kotake-Nara, E.; Nagao, A. Absorption and metabolism of xanthophylls. Mar. Drugs 2011, 9, 1024–1037. [Google Scholar] [CrossRef]
- Harrison, E.H. Mechanisms involved in the intestinal absorption of dietary vitamin A and provitamin A carotenoids. Biochim. Biophys. Acta 2012, 1821, 70–77. [Google Scholar] [CrossRef]
- Kopec, R.E.; Failla, M.L. Recent advances in the bioaccessibility and bioavailability of carotenoids and effects of other dietary lipophiles. J. Food Compos. Anal. 2018, 68, 16–30. [Google Scholar] [CrossRef]
- Etoh, H.; Utsunomiya, Y.; Komori, A.; Murakami, Y.; Oshima, S.; Inakuma, T. Carotenoids in human blood plasma after ingesting paprika juice. Biosci. Biotechnol. Biochem. 2000, 64, 1096–1098. [Google Scholar] [CrossRef]
- Nishino, A.; Ichihara, T.; Takaha, T.; Kuriki, T.; Nihei, H.; Kawamoto, K.; Yasui, H.; Maoka, T. Accumulation of paprika carotenoids in human plasma and erythrocytes. J. Oleo Sci. 2015, 64, 1135–1142. [Google Scholar] [CrossRef]
- Umigai, N.; Murakami, K.; Shimizu, R.; Takeda, R.; Azuma, T. Safety evaluation and plasma carotenoid accumulation in healthy adult subjects after 12 weeks of paprika oleoresin supplementation. J. Oleo Sci. 2018, 67, 225–234. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, L.; Jiwan, M.A.; Daly, T.; O’Brien, N.M.; Aherne, S.A. Bioaccessibility, uptake, and transport of carotenoids from peppers (Capsicum spp.) using the coupled in vitro digestion and human intestinal Caco-2 cell model. J. Agric. Food Chem. 2010, 58, 5374–5379. [Google Scholar] [CrossRef] [PubMed]
- Pugliese, A.; O’Callaghan, Y.; Tundis, R.; Galvin, K.; Menichini, F.; O’Brien, N.; Loizzo, M.R. In vitro assessment of the bioaccessibility of carotenoids from sun-dried chilli peppers. Plant Foods Hum. Nutr. 2014, 69, 8–17. [Google Scholar] [CrossRef]
- McClements, D.J. Nanoscale nutrient delivery systems for food applications: Improving bioactive dispersibility, stability, and bioavailability. J. Food Sci. 2015, 80, N1602–N1611. [Google Scholar] [CrossRef] [PubMed]
- Jomova, K.; Valko, M. Health protective effects of carotenoids and their interactions with other biological antioxidants. Eur. J. Med. Chem. 2013, 70, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, D.; Freitas, M.; Silva, A.M.S.; Carvalho, F.; Fernandes, E. Antioxidant and pro-oxidant activities of carotenoids and their oxidation products. Food Chem. Toxicol. 2018, 120, 681–699. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D.A. Dietary carotenoids and human immune function. Nutrition 2001, 17, 823–827. [Google Scholar] [CrossRef]
- Kim, J.S.; Ahn, J.; Lee, S.J.; Moon, B.; Ha, T.Y.; Kim, S. Phytochemicals and antioxidant activity of fruits and leaves of paprika (Capsicum Annuum L., var. special) cultivated in Korea. J. Food Sci. 2011, 76, C193–C198. [Google Scholar] [CrossRef]
- Iqbal, Q.; Amjad, M.; Asi, M.R.; Arino, A. Characterization of capsaicinoids and antioxidants in hot peppers as influenced by hybrid and harvesting stage. Plant Foods Hum. Nutr. 2013, 68, 358–363. [Google Scholar] [CrossRef]
- Meckelmann, S.W.; Riegel, D.W.; van Zonneveld, M.; Ríos, L.; Peña, K.; Mueller-Seitz, E.; Petz, M. Capsaicinoids, flavonoids, tocopherols, antioxidant capacity and color attributes in 23 native Peruvian chili peppers (Capsicum spp.) grown in three different locations. Eur. Food Res. Tech. 2014, 240, 273–283. [Google Scholar] [CrossRef]
- Sinisgalli, C.; Faraone, I.; Vassallo, A.; Caddeo, C.; Bisaccia, F.; Armentano, M.F.; Milella, L.; Ostuni, A. Phytochemical profile of Capsicum annuum L. cv Senise, incorporation into liposomes, and evaluation of cellular antioxidant activity. Antioxidants 2020, 9, 428. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Lee, W.M.; Rhee, H.C.; Kim, S. Red paprika (Capsicum annuum L.) and its main carotenoids, capsanthin and beta-carotene, prevent hydrogen peroxide-induced inhibition of gap-junction intercellular communication. Chem. Biol. Interact. 2016, 254, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Matsufuji, H.; Nakamura, H.; Chino, M.; Takeda, M. Antioxidant activity of capsanthin and the fatty acid esters in paprika (Capsicum annuum). J. Agric. Food Chem. 1998, 46, 3468–3472. [Google Scholar] [CrossRef]
- Murakami, A.; Nakashima, M.; Koshiba, T.; Maoka, T.; Nishino, H.; Yano, M.; Sumida, T.; Kim, O.K.; Koshimizu, K.; Ohigashi, H. Modifying effects of carotenoids on superoxide and nitric oxide generation from stimulated leukocytes. Cancer Lett. 2000, 149, 115–123. [Google Scholar] [CrossRef]
- Kim, S.; Ha, T.Y.; Hwang, I.K. Analysis, bioavailability, and potential healthy effects of capsanthin, natural red pigment from Capsicum spp. Food Rev. Int. 2009, 25, 198–213. [Google Scholar] [CrossRef]
- Elvira-Torales, L.I.; García-Alonso, J.; Periago-Castón, M.J. Nutritional importance of carotenoids and their effect on liver health: A review. Antioxidants 2019, 8, 229. [Google Scholar] [CrossRef] [PubMed]
- Terasaki, M.; Mutoh, M.; Fujii, G.; Takahashi, M.; Ishigamori, R.; Masuda, S. Potential ability of xanthophylls to prevent obesity-associated cancer. World J. Pharmacol. 2014, 3, 140–152. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y.S.; Daglia, M.; Rengasamy, K.R. Dietary carotenoids in cancer chemoprevention and chemotherapy: A review of emerging evidence. Pharmacol. Res. 2020, 157, 104830. [Google Scholar] [CrossRef]
- Shin, J.; Song, M.H.; Oh, J.W.; Keum, Y.S.; Saini, R.K. Pro-oxidant actions of carotenoids in triggering apoptosis of cancer cells: A review of emerging evidence. Antioxidants 2020, 9, 532. [Google Scholar] [CrossRef]
- Koklesova, L.; Liskova, A.; Samec, M.; Buhrmann, C.; Samuel, S.M.; Varghese, E.; Ashrafizadeh, M.; Najafi, M.; Shakibaei, M.; Busselberg, D.; et al. Carotenoids in cancer apoptosis-The road from bench to bedside and back. Cancers 2020, 12, 2425. [Google Scholar] [CrossRef]
- Antwi, S.O.; Steck, S.E.; Zhang, H.; Stumm, L.; Zhang, J.; Hurley, T.G.; Hebert, J.R. Plasma carotenoids and tocopherols in relation to prostate-specific antigen (PSA) levels among men with biochemical recurrence of prostate cancer. Cancer Epidemiol. 2015, 39, 752–762. [Google Scholar] [CrossRef]
- Voutilainen, S.; Nurmi, T.; Mursu, J.; Rissanen, T.H. Carotenoids and cardiovascular health. Am. J. Clin. Nutr. 2006, 83, 1265–1271. [Google Scholar] [CrossRef] [PubMed]
- Pietro, N. Carotenoids in cardiovascular disease prevention. JSM Atheroscler. 2016, 1, 1002. [Google Scholar] [CrossRef]
- Gammone, M.A.; Riccioni, G.; D’Orazio, N. Carotenoids: Potential allies of cardiovascular health? Food Nutr. Res. 2015, 59, 26762. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Leite, J.O.; DeOgburn, R.; Smyth, J.A.; Clark, R.M.; Fernandez, M.L. A lutein-enriched diet prevents cholesterol accumulation and decreases oxidized LDL and inflammatory cytokines in the aorta of guinea pigs. J. Nutr. 2011, 141, 1458–1463. [Google Scholar] [CrossRef] [PubMed]
- Zou, Z.; Xu, X.; Huang, Y.; Xiao, X.; Ma, L.; Sun, T.; Dong, P.; Wang, X.; Lin, X. High serum level of lutein may be protective against early atherosclerosis: The Beijing atherosclerosis study. Atherosclerosis 2011, 219, 789–793. [Google Scholar] [CrossRef]
- Leermakers, E.T.; Darweesh, S.K.; Baena, C.P.; Moreira, E.M.; Melo van Lent, D.; Tielemans, M.J.; Muka, T.; Vitezova, A.; Chowdhury, R.; Bramer, W.M.; et al. The effects of lutein on cardiometabolic health across the life course: A systematic review and meta-analysis1,2. Am. J. Clin. Nutr. 2016, 103, 481–494. [Google Scholar] [CrossRef]
- Hozawa, A.; Jacobs, D.R., Jr.; Steffes, M.W.; Gross, M.D.; Steffen, L.M.; Lee, D.H. Circulating carotenoid concentrations and incident hypertension: The Coronary Artery Risk Development in Young Adults (CARDIA) study. J. Hypertens. 2009, 27, 237–242. [Google Scholar] [CrossRef]
- Lu, C.-C.; Yen, G.-C. Antioxidative and anti-inflammatory activity of functional foods. Curr. Opin. Food Sci. 2015, 2, 1–8. [Google Scholar] [CrossRef]
- Mohammadzadeh Honarvar, N.; Saedisomeolia, A.; Abdolahi, M.; Shayeganrad, A.; Taheri Sangsari, G.; Hassanzadeh Rad, B.; Muench, G. Molecular anti-inflammatory mechanisms of retinoids and carotenoids in Alzheimer’s disease: A review of current evidence. J. Mol. Neurosci. 2017, 61, 289–304. [Google Scholar] [CrossRef]
- Jin, X.H.; Ohgami, K.; Shiratori, K.; Suzuki, Y.; Hirano, T.; Koyama, Y.; Yoshida, K.; Ilieva, I.; Iseki, K.; Ohno, S. Inhibitory effects of lutein on endotoxin-induced uveitis in Lewis rats. Invest. Ophthalmol. Vis. Sci. 2006, 47, 2562–2568. [Google Scholar] [CrossRef] [PubMed]
- Pattison, D.J.; Symmons, D.P.; Lunt, M.; Welch, A.; Bingham, S.A.; Day, N.E.; Silman, A.J. Dietary beta-cryptoxanthin and inflammatory polyarthritis: Results from a population-based prospective study. Am. J. Clin. Nutr. 2005, 82, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Di Tomo, P.; Canali, R.; Ciavardelli, D.; Di Silvestre, S.; De Marco, A.; Giardinelli, A.; Pipino, C.; Di Pietro, N.; Virgili, F.; Pandolfi, A. Beta-carotene and lycopene affect endothelial response to TNF-alpha reducing nitro-oxidative stress and interaction with monocytes. Mol. Nutr. Food Res. 2012, 56, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Pechinskii, S.; Kuregyan, A. The impact of carotenoids on immunity. Pharm. Chem. J. 2014, 47, 509–513. [Google Scholar] [CrossRef]
- Chew, B.P.; Park, J.S. Carotenoid action on the immune response. J. Nutr. 2004, 134, 257S–261S. [Google Scholar] [CrossRef]
- Toti, E.; Chen, C.Y.O.; Palmery, M.; Villaño Valencia, D.; Peluso, I. Non-provitamin A and provitamin A carotenoids as immunomodulators: Recommended dietary allowance, therapeutic index, or personalized nutrition? Oxid. Med. Cell. Longev. 2018, 2018, 4637861. [Google Scholar] [CrossRef]
- Ghodratizadeh, S.; Kanbak, G.; Beyramzadeh, M.; Dikmen, Z.G.; Memarzadeh, S.; Habibian, R. Effect of carotenoid β-cryptoxanthin on cellular and humoral immune response in rabbit. Vet. Res. Commun. 2014, 38, 59–62. [Google Scholar] [CrossRef]
- Bonet, M.L.; Canas, J.A.; Ribot, J.; Palou, A. Carotenoids in adipose tissue biology and obesity. In Carotenoids in Nature: Biosynthesis, Regulation and Function; Stange, C., Ed.; Springer: Cham, Switzerland, 2016; Volume 79, pp. 377–414. [Google Scholar]
- Maeda, H.; Saito, S.; Nakamura, N.; Maoka, T. Paprika pigments attenuate obesity-induced inflammation in 3T3-L1 adipocytes. ISRN Inflamm. 2013, 2013, 763758. [Google Scholar] [CrossRef]
- Kakutani, R.; Hokari, S.; Nishino, A.; Ichihara, T.; Sugimoto, K.; Takaha, T.; Kuriki, T.; Maoka, T. Effect of oral paprika xanthophyll intake on abdominal fat in healthy overweight humans: A randomized, double-blind, placebo-controlled study. J. Oleo. Sci. 2018, 67, 1149–1162. [Google Scholar] [CrossRef]
- Melendez-Martínez, A.J.; Stinco, C.M.; Mapelli-Brahm, P. Skin carotenoids in public health and nutricosmetics: The emerging roles and applications of the UV radiation-absorbing colourless carotenoids phytoene and phytofluene. Nutrients 2019, 11, 1093. [Google Scholar] [CrossRef]
- Nishino, A.; Sugimoto, K.; Sambe, H.; Ichihara, T.; Takaha, T.; Kuriki, T. Effects of dietary paprika xanthophylls on ultraviolet light-induced skin damage: A double-blind placebo-controlled study. J. Oleo Sci. 2018, 67, 863–869. [Google Scholar] [CrossRef] [PubMed]
- Kamoshita, M.; Toda, E.; Osada, H.; Narimatsu, T.; Kobayashi, S.; Tsubota, K.; Ozawa, Y. Lutein acts via multiple antioxidant pathways in the photo-stressed retina. Sci. Rep. 2016, 6, 30226. [Google Scholar] [CrossRef]
- Sluijs, I.; Cadier, E.; Beulens, J.W.; van der, A.D.; Spijkerman, A.M.; van der Schouw, Y.T. Dietary intake of carotenoids and risk of type 2 diabetes. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 376–381. [Google Scholar] [CrossRef]
- Llopis, S.; Rodrigo, M.J.; Gonzalez, N.; Genoves, S.; Zacarias, L.; Ramon, D.; Martorell, P. Beta-cryptoxanthin reduces body fat and increases oxidative stress response in Caenorhabditis elegans model. Nutrients 2019, 11, 232. [Google Scholar] [CrossRef] [PubMed]
- Cannavale, C.N.; Hassevoort, K.M.; Edwards, C.G.; Thompson, S.V.; Burd, N.A.; Holscher, H.D.; Erdman, J.W., Jr.; Cohen, N.J.; Khan, N.A. Serum lutein is related to relational memory performance. Nutrients 2019, 11, 768. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.S.; Shin, M.; Kim, S.; Lee, S.B. Recent advances in studies on the therapeutic potential of dietary carotenoids in neurodegenerative diseases. Oxid. Med. Cell Longev. 2018, 2018, 4120458. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Carotenoid | Health Effect | Mechanism of Action | Reference |
|---|---|---|---|
| Lutein | Gastric cancer | Not determined (ND) | [15] |
| Cancer cells | Modulation of apoptosis and multidrug resistance | [16] | |
| Edema reduction | Reduction acetylcholinesterase, Increase seromucoids | [17] | |
| Retina damage | Modulation of oxidative stress, and pro-inflammatory gene expression | [18] | |
| Macular degeneration | Absorption of UV radiation, antioxidant | [19] | |
| β-carotene | Prostate cancer | Inverse correlation with prostate-specific antigen (PSA) occurrence | [20,21] |
| Gastric cancer | ND | [15] | |
| Anti-inflammatory, analgesic, antinociceptive | ND | [22] | |
| Edema reduction | Reduction acetylcholinesterase, Increase seromucoids | [17] | |
| Obesity | Promotion of fatty acid oxidation | [23] | |
| β-cryptoxanthin | Prostate cancer | Inverse correlation with PSA occurrence | [20,21] |
| Gastric cancer | ND | [15] | |
| Cancer prevention | Modulation of signaling pathways | [24] | |
| Anti-inflammatory, analgesic, antinociceptive | ND | [22] | |
| Zeaxanthin | Gastric cancer | ND | [15] |
| Obesity | Activation of AMP-activated protein (AMPK) and inhibition of lipogenesis | [25] | |
| Macular degeneration | Absorption of UV radiation, antioxidant | [19] | |
| Alzheimer disease | Inhibition of acetylcholinesterase, butyrylcholinesterase and β-secretase | [26] | |
| Violaxanthin | Cancer cells | Modulation of apoptosis and multidrug resistance | [16] |
| Anti-inflammatory, analgesic, antinociceptive | ND | [22] | |
| Capsanthin | Colon cancer | Inhibitory effect | [27] |
| Skin cancer | Chemopreventive | [28] | |
| Cancer cells | Modulation of apoptosis and multidrug resistance | [16] | |
| Cancer breast(MCF-7 cells) | Oxidative stress, DNA damage, increase p53 and Bax, lipid peroxidation | [29] | |
| Atherosclerosis | increase in the cholesterol efflux | [30] | |
| Edema reduction | Reduction of acetylcholinesterase, Increase seromucoids | [17] | |
| Obesity | Suppression of hepatic lipogenesis, fatty acid oxidation, and gluconeogenesis. Inhibit adipogenesis | [31] | |
| Obesity and insulin sensitizing | Inhibition of adipogenesis, increase of lipolytic activity, accelerated oxidation of fatty acids | [32] | |
| Atherosclerosis | Decrease on serum levels of total cholesterol, triglycerides, low density lipoprotein cholesterol, prebiotic | [33] | |
| Skin health | Counteract the cytotoxic effect of UV radiation by decreasing the formation of DNA strand breaks | [34] | |
| Diabetes | Improvement of glucose tolerance, improvement of insulin sensitivity | [33] | |
| Alzheimer disease | Inhibiting acetylcholinesterase, butyrylcholinesterase and β-secretase | [26] | |
| Capsorubin | Cancer cells | Modulation of apoptosis and multidrug resistance | [16] |
| Skin health | Counteract the cytotoxic effect of UV radiation by decreasing the formation of DNA strand breaks | [34] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villa-Rivera, M.G.; Ochoa-Alejo, N. Chili Pepper Carotenoids: Nutraceutical Properties and Mechanisms of Action. Molecules 2020, 25, 5573. https://doi.org/10.3390/molecules25235573
Villa-Rivera MG, Ochoa-Alejo N. Chili Pepper Carotenoids: Nutraceutical Properties and Mechanisms of Action. Molecules. 2020; 25(23):5573. https://doi.org/10.3390/molecules25235573
Chicago/Turabian StyleVilla-Rivera, Maria Guadalupe, and Neftalí Ochoa-Alejo. 2020. "Chili Pepper Carotenoids: Nutraceutical Properties and Mechanisms of Action" Molecules 25, no. 23: 5573. https://doi.org/10.3390/molecules25235573
APA StyleVilla-Rivera, M. G., & Ochoa-Alejo, N. (2020). Chili Pepper Carotenoids: Nutraceutical Properties and Mechanisms of Action. Molecules, 25(23), 5573. https://doi.org/10.3390/molecules25235573