
Essential Oil of Calotropis procera: Comparative Chemical Profiles, Antimicrobial Activity, and Allelopathic Potential on Weeds


 ,
,  , ,
, ,  ,
, 

Abstract
1. Introduction
2. Results and Discussion
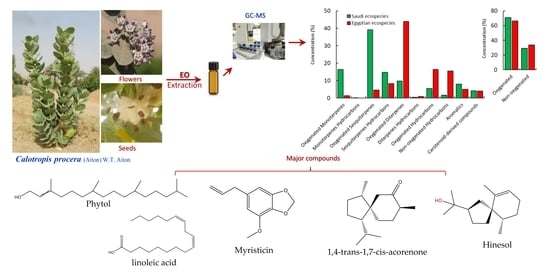
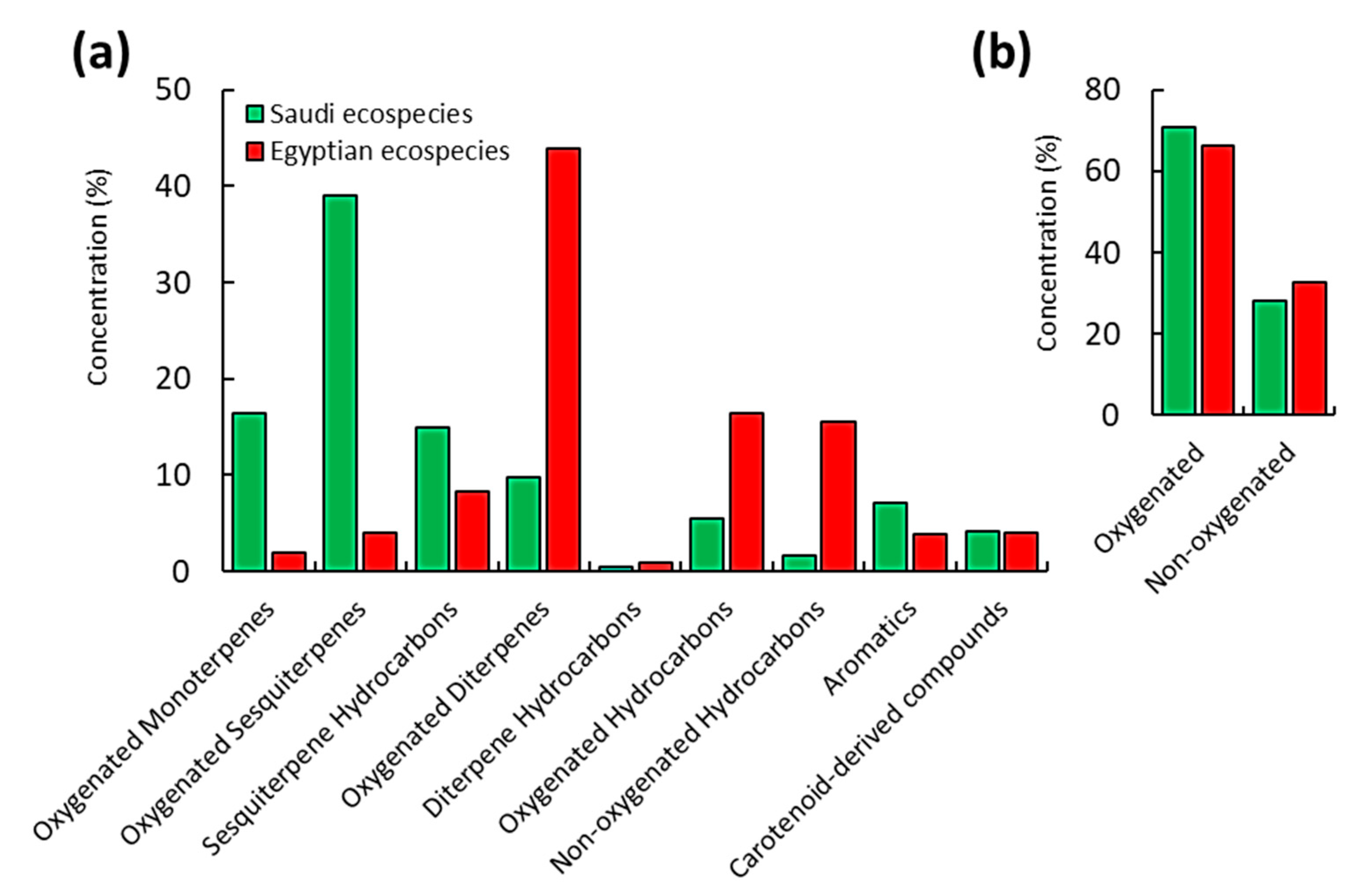
2.1. Chemical Profiles of EOs of C. Procera
2.2. Principal Components Analysis (PCA) and Agglomerative Hierarchical Clustering (AHC)
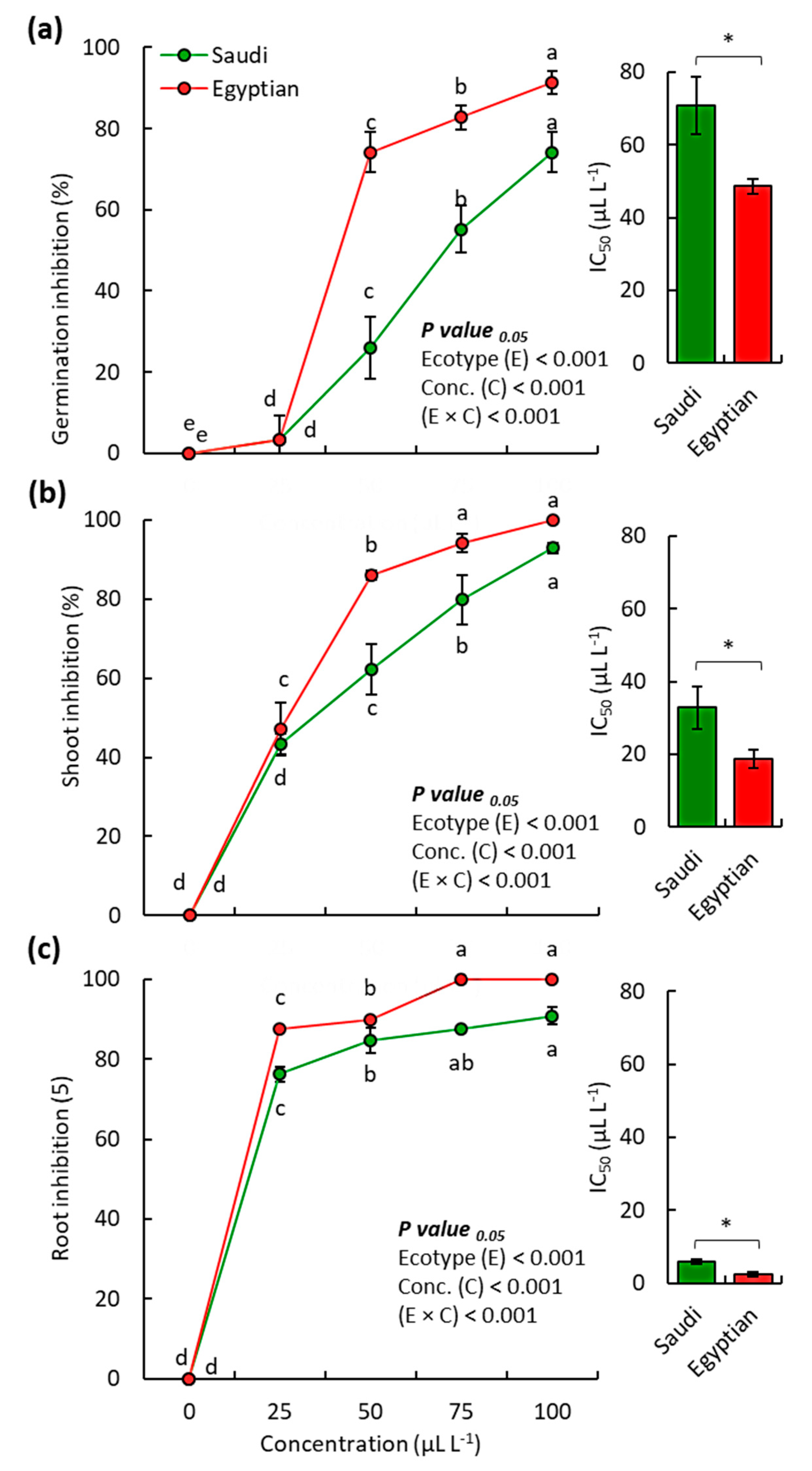
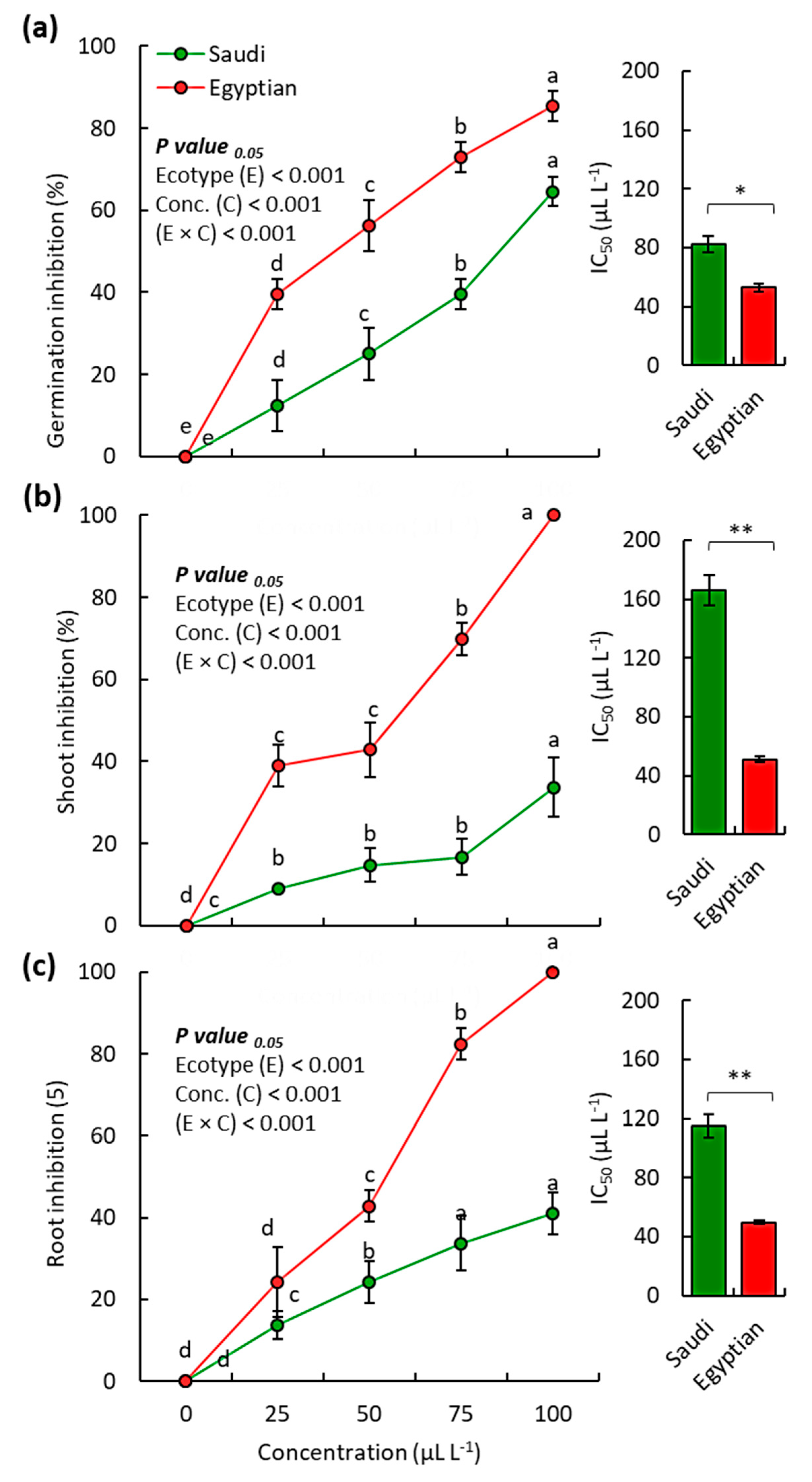
2.3. Allelopathic Activity of the EOs
2.4. Antimicrobial Activities
3. Materials and Methods
3.1. Plant Materials Collection and Identification
3.2. Essential Oil Extraction, GC-MS Analysis, and Constituents’ Identification
3.3. Allelopathic Activity of the EOs
3.4. Antimicrobial Properties of EOs
3.5. Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cumo, C. Plants and People: Origin and Development of Human-Plant Science Relationships; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Elshamy, A.I.; Farrag, A.R.H.; Ayoub, I.M.; Mahdy, K.A.; Taher, R.F.; Gendy, A.E.-N.G.; Mohamed, T.A.; Al-Rejaie, S.S.; Ei-Amier, Y.A.; Abd-EIGawad, A.M. UPLC-qTOF-MS phytochemical profile and antiulcer potential of Cyperus conglomeratus Rottb. alcoholic extract. Molecules 2020, 25, 4234. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wu, Y.; Wang, M. Bioactive Substances of Plant Origin. In Handbook of Food Chemistry; Cheung, P.C.K., Mehta, B.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 967–1008. [Google Scholar]
- Yoon, H.I.; Zhang, W.; Son, J.E. Optimal duration of drought stress near harvest for promoting bioactive compounds and antioxidant capacity in kale with or without UV-B radiation in plant factories. Plants 2020, 9, 295. [Google Scholar] [CrossRef] [PubMed]
- Kliebenstein, D. Plant defense compounds: Systems approaches to metabolic analysis. Ann. Rev. Phytopathol. 2012, 50, 155–173. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Gawad, A.; Zotti, M.; Sarker, T.; Mazzoleni, S.; Bonanomi, G. Does a plant detect its neighbor if it is kin or stranger? Evidence from a common garden experiment. Community Ecol. 2017, 18, 305–310. [Google Scholar] [CrossRef]
- Ramos, M.V.; Demarco, D.; da Costa Souza, I.C.; de Freitas, C.D.T. Laticifers, latex, and their role in plant defense. Trends Plant Sci. 2019, 24, 553–567. [Google Scholar] [CrossRef] [PubMed]
- Yactayo-Chang, J.P.; Tang, H.V.; Mendoza, J.; Christensen, S.A.; Block, A.K. Plant defense chemicals against insect pests. Agronomy 2020, 10, 1156. [Google Scholar] [CrossRef]
- Abd-ElGawad, A.M.; El-Amier, Y.A.; Assaeed, A.M.; Al-Rowaily, S.L. Interspecific variations in the habitats of Reichardia tingitana (L.) Roth leading to changes in its bioactive constituents and allelopathic activity. Saudi J. Biol. Sci. 2020, 27, 489–499. [Google Scholar] [CrossRef]
- Assaeed, A.; Elshamy, A.; El Gendy, A.E.-N.; Dar, B.; Al-Rowaily, S.; Abd-ElGawad, A. Sesquiterpenes-rich essential oil from above ground parts of Pulicaria somalensis exhibited antioxidant activity and allelopathic effect on weeds. Agronomy 2020, 10, 399. [Google Scholar] [CrossRef]
- Elshamy, A.; Abd-ElGawad, A.M.; El-Amier, Y.A.; El Gendy, A.; Al-Rowaily, S. Interspecific variation, antioxidant and allelopathic activity of the essential oil from three Launaea species growing naturally in heterogeneous habitats in Egypt. Flavour Fragr. J. 2019, 34, 316–328. [Google Scholar] [CrossRef]
- Vamanu, E.; Gatea, F. Correlations between microbiota bioactivity and bioavailability of functional compounds: A mini-review. Biomedicines 2020, 8, 39. [Google Scholar] [CrossRef]
- Abd El-Gawad, A.M. Chemical constituents, antioxidant and potential allelopathic effect of the essential oil from the aerial parts of Cullen plicata. Ind. Crops Prod. 2016, 80, 36–41. [Google Scholar] [CrossRef]
- Elshamy, A.I.; Ammar, N.M.; Hassan, H.A.; Al-Rowaily, S.L.; Raga, T.R.; El Gendy, A.; Abd-ElGawad, A.M. Essential oil and its nanoemulsion of Araucaria heterophylla resin: Chemical characterization, anti-inflammatory, and antipyretic activities. Ind. Crops Prod. 2020, 148, 112272. [Google Scholar] [CrossRef]
- Diniz do Nascimento, L.; Moraes, A.A.B.d.; Costa, K.S.d.; Pereira Galúcio, J.M.; Taube, P.S.; Costa, C.M.L.; Neves Cruz, J.; de Aguiar Andrade, E.H.; Faria, L.J.G.d. Bioactive natural compounds and antioxidant activity of essential oils from spice plants: New findings and potential applications. Biomolecules 2020, 10, 988. [Google Scholar] [CrossRef]
- Amin, S.M.; Hassan, H.M.; El Gendy, A.E.N.G.; El-Beih, A.A.; Mohamed, T.A.; Elshamy, A.I.; Bader, A.; Shams, K.A.; Mohammed, R.; Hegazy, M.E.F. Comparative chemical study and antimicrobial activity of essential oils of three Artemisia species from Egypt and Saudi Arabia. Flavour Fragr. J. 2019, 34, 450–459. [Google Scholar] [CrossRef]
- Baptista-Silva, S.; Borges, S.; Ramos, O.L.; Pintado, M.; Sarmento, B. The progress of essential oils as potential therapeutic agents: A review. J. Essent. Oil Res. 2020, 1–17. [Google Scholar]
- Nea, F.; Kambiré, D.A.; Genva, M.; Tanoh, E.A.; Wognin, E.L.; Martin, H.; Brostaux, Y.; Tomi, F.; Lognay, G.C.; Tonzibo, Z.F. Composition, seasonal variation, and biological activities of Lantana camara essential oils from Côte d’Ivoire. Molecules 2020, 25, 2400. [Google Scholar] [CrossRef]
- Hassan, L.M.; Galal, T.M.; Farahat, E.A.; El-Midany, M.M. The biology of Calotropis procera (Aiton) WT. Trees 2015, 29, 311–320. [Google Scholar] [CrossRef]
- Silva, M.C.C.; da Silva, A.B.; Teixeira, F.M.; de Sousa, P.C.P.; Rondon, R.M.M.; Júnior, J.E.R.H.; Sampaio, L.R.L.; Oliveira, S.L.; Holonda, A.N.M.; de Vasconcelos, S.M.M. Therapeutic and biological activities of Calotropis procera (Ait.) R. Br. Asian Pac. J. Trop. Med. 2010, 3, 332–336. [Google Scholar] [CrossRef]
- Sharma, R.; Thakur, G.S.; Sanodiya, B.S.; Savita, A.; Pandey, M.; Sharma, A.; Bisen, P.S. Therapeutic potential of Calotropis procera: A giant milkweed. IOSR J. Pharm. Biol. Sci. 2012, 4, 42–57. [Google Scholar] [CrossRef]
- Rani, R.; Sharma, D.; Chaturvedi, M.; Yadav, J. Antibacterial activity of twenty different endophytic fungi isolated from Calotropis procera and time kill assay. Clin. Microbiol. 2017, 6, 280. [Google Scholar] [CrossRef]
- Nadeem, M.; Mumtaz, M.W.; Danish, M.; Rashid, U.; Mukhtar, H.; Anwar, F.; Raza, S.A. Calotropis procera: UHPLC-QTOF-MS/MS based profiling of bioactives, antioxidant and anti-diabetic potential of leaf extracts and an insight into molecular docking. J. Food Meas. Charact. 2019, 13, 3206–3220. [Google Scholar] [CrossRef]
- Al-Snafi, A. The constituents and pharmacological properties of Calotropis procera-An Overview. Int. J. Pharm. Rev. Res. 2015, 5, 259–275. [Google Scholar]
- Basu, A.; Chaudhuri, A.N. Preliminary studies on the antiinflammatory and analgesic activities of Calotropis procera root extract. J. Ethnopharmacol. 1991, 31, 319–324. [Google Scholar] [CrossRef]
- Falana, M.B.; Nurudeen, Q.O. Evaluation of phytochemical constituents and in vitro antimicrobial activities of leaves extracts of Calotropis procera against certain human pathogens. Not. Sci. Biol. 2020, 12, 208–221. [Google Scholar] [CrossRef]
- Pattnaik, P.K.; Kar, D.; Chhatoi, H.; Shahbazi, S.; Ghosh, G.; Kuanar, A. Chemometric profile & antimicrobial activities of leaf extract of Calotropis procera and Calotropis gigantea. Nat. Prod. Res. 2017, 31, 1954–1957. [Google Scholar]
- Singh, M.; Javed, K. Chemical characterization and antimicrobial activity of Calotropis gigantea Linn. flower essential oil collected from northern plain of India. Int. J. Adv. Biotechnol. Res. 2013, 4, 533–541. [Google Scholar]
- Kubmarawa, D.; Ogunwande, I.A. Composition of the leaves essential oil of Calotropis procera (R. Br.) from Nigeria. J. Essent. Oil Bear. Plants 2008, 11, 75–78. [Google Scholar] [CrossRef]
- Okiei, W.; Ogunlesi, M.; Ofor, E.; Osibote, E. Analysis of essential oil constituents in hydro-distillates of Calotropis procera. Res. J. Phytochem. 2009, 3, 44–53. [Google Scholar] [CrossRef][Green Version]
- Wahba, H.E.; Khalid, A.K. Comparative study on essential oil composition in various organs of Sodom apple (Calotropis procera) grown wild in Egypt. Asian J. Plant Sci. 2018, 17, 85–90. [Google Scholar] [CrossRef][Green Version]
- Abd El-Gawad, A.M.; Elshamy, A.I.; El Gendy, A.E.-N.; Gaara, A.; Assaeed, A.M. Volatiles profiling, allelopathic activity, and antioxidant potentiality of Xanthium strumarium leaves essential oil from Egypt: Evidence from chemometrics analysis. Molecules 2019, 24, 584. [Google Scholar] [CrossRef]
- Abd-ElGawad, A.M.; Elshamy, A.I.; Al-Rowaily, S.L.; El-Amier, Y.A. Habitat affects the chemical profile, allelopathy, and antioxidant properties of essential oils and phenolic enriched extracts of the invasive plant Heliotropium curassavicum. Plants 2019, 8, 482. [Google Scholar] [CrossRef]
- Abd-ElGawad, A.M.; El Gendy, A.E.-N.G.; Assaeed, A.M.; Al-Rowaily, S.L.; Omer, E.A.; Dar, B.A.; Al-Taisan, W.a.A.; Elshamy, A.I. Essential oil enriched with oxygenated constituents from invasive plant Argemone ochroleuca exhibited potent phytotoxic effects. Plants 2020, 9, 998. [Google Scholar] [CrossRef]
- Abd-ElGawad, A.M.; Elshamy, A.; El Gendy, A.E.-N.; Al-Rowaily, S.L.; Assaeed, A.M. Preponderance of oxygenated sesquiterpenes and diterpenes in the volatile oil constituents of Lactuca serriola L. revealed antioxidant and allelopathic activity. Chem. Biodivers. 2019, 16, e1900278. [Google Scholar] [CrossRef]
- Verdeguer, M.; Blázquez, M.A.; Boira, H. Phytotoxic effects of Lantana camara, Eucalyptus camaldulensis and Eriocephalus africanus essential oils in weeds of Mediterranean summer crops. Biochem. Syst. Ecol. 2009, 37, 362–369. [Google Scholar] [CrossRef]
- de Almeida, L.F.R.; Frei, F.; Mancini, E.; De Martino, L.; De Feo, V. Phytotoxic activities of Mediterranean essential oils. Molecules 2010, 15, 4309–4323. [Google Scholar] [CrossRef]
- Mancini, E.; Arnold, N.A.; De Martino, L.; De Feo, V.; Formisano, C.; Rigano, D.; Senatore, F. Chemical composition and phytotoxic effects of essential oils of Salvia hierosolymitana Boiss. and Salvia multicaulis Vahl. var. simplicifolia Boiss. growing wild in Lebanon. Molecules 2009, 14, 4725–4736. [Google Scholar] [CrossRef]
- Elshamy, A.I.; Abd El-Gawad, A.M.; El Gendy, A.E.-N.G.; Assaeed, A.M. Chemical characterization of Euphorbia heterophylla L. essential oils and their antioxidant activity and allelopathic potential on Cenchrus echinatus L. Chem. Biodivers. 2019, 16, e1900051. [Google Scholar] [CrossRef]
- Bozok, F.; Ulukanli, Z. Volatiles from the aerial parts of east Mediterranean clary sage: Phytotoxic activity. J. Essent. Oil Bear. Plants 2016, 19, 1192–1198. [Google Scholar] [CrossRef]
- Bali, A.S.; Batish, D.R.; Singh, H.P.; Kaur, S.; Kohli, R.K. Chemical characterization and phytotoxicity of foliar volatiles and essential oil of Callistemon viminalis. J. Essent. Oil Bear. Plants 2017, 20, 535–545. [Google Scholar] [CrossRef]
- Ulukanli, Z.; Cenet, M.; Ince, H.; Yilmaztekin, M. Antimicrobial and herbicidal activities of the essential oil from the Mediterranean Thymus eigii. J. Essent. Oil Bear. Plants 2018, 21, 214–222. [Google Scholar] [CrossRef]
- Saleh, I.; Abd-ElGawad, A.; El Gendy, A.E.-N.; Abd El Aty, A.; Mohamed, T.; Kassem, H.; Aldosri, F.; Elshamy, A.; Hegazy, M.-E.F. Phytotoxic and antimicrobial activities of Teucrium polium and Thymus decussatus essential oils extracted using hydrodistillation and microwave-assisted techniques. Plants 2020, 9, 716. [Google Scholar] [CrossRef]
- Azizi, A.; Yan, F.; Honermeier, B. Herbage yield, essential oil content and composition of three oregano (Origanum vulgare L.) populations as affected by soil moisture regimes and nitrogen supply. Ind. Crops Prod. 2009, 29, 554–561. [Google Scholar] [CrossRef]
- Abd-ElGawad, A.M.; Elshamy, A.; El-Amier, Y.A.; El Gendy, A.; Al-Barati, S.; Dar, B.; Al-Rowaily, S.; Assaeed, A. Chemical composition variations, allelopathic, and antioxidant activities of Symphyotrichum squamatum (Spreng.) Nesom essential oils growing in heterogeneous habitats. Arab. J. Chem. 2020, 13, 237–4245. [Google Scholar] [CrossRef]
- Holm, L.G.; Plucknett, D.L.; Pancho, J.V.; Herberger, J.P. The World’s WorstWeeds: Distribution and Biology; University Press of Hawaii: Honolulu, HI, USA, 1977. [Google Scholar]
- Arora, K.; Batish, D.; Kohli, R.; Singh, H. Allelopathic impact of essential oil of Tagetes minuta on common agricultural and wasteland weeds. Innovare J. Agric. Sci. 2017, 5, 1–4. [Google Scholar]
- Menchari, Y.; Camilleri, C.; Michel, S.; Brunel, D.; Dessaint, F.; Le Corre, V.; Délye, C. Weed response to herbicides: Regional-scale distribution of herbicide resistance alleles in the grass weed Alopecurus myosuroides. New Phytol. 2006, 171, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Liu, S.; Ju, X.; Du, J.; Xu, B.; Yuan, H.; Qin, F.; Li, L. The antitumor effect of hinesol, extract from Atractylodes lancea (Thunb.) DC. by proliferation, inhibition, and apoptosis induction via MEK/ERK and NF-κB pathway in non–small cell lung cancer cell lines A549 and NCI-H1299. J. Cell. Biochem. 2019, 120, 18600–18607. [Google Scholar] [CrossRef]
- Satoh, K.; Nagai, F.; Kano, I. Inhibition of H+, K+-ATPase by hinesol, a major component of So-jutsu, by interaction with enzyme in the E1 state. Biochem. Pharmacol. 2000, 59, 881–886. [Google Scholar] [CrossRef]
- Hosni, K.; Hassen, I.; Sebei, H.; Casabianca, H. Secondary metabolites from Chrysanthemum coronarium (Garland) flowerheads: Chemical composition and biological activities. Ind. Crops Prod. 2013, 44, 263–271. [Google Scholar] [CrossRef]
- Ulukanli, Z.; Demirci, S.; Yilmaztekin, M. Essential oil constituents of Tanacetum cilicicum: Antimicrobial and phytotoxic activities. J. Food Qual. 2017, 2017, 6214896. [Google Scholar] [CrossRef]
- Trong Le, N.; Viet Ho, D.; Quoc Doan, T.; Tuan Le, A.; Raal, A.; Usai, D.; Sanna, G.; Carta, A.; Rappelli, P.; Diaz, N. Biological activities of essential oils from leaves of Paramignya trimera (Oliv.) Guillaum and Limnocitrus littoralis (Miq.) Swingle. Antibiotics 2020, 9, 207. [Google Scholar] [CrossRef]
- Mohamed, T.A.; Abd El Aty, A.A.; Shahat, A.A.; Abdel-Azim, N.S.; Shams, K.A.; Elshamy, A.A.; Ahmed, M.M.; Youns, S.H.; El-Wassimy, T.M.; El-Toumy, S.A. New antimicrobial metabolites from the medicinal herb Artemisia herba-Alba. Nat. Prod. Res. 2019, 1–9. [Google Scholar] [CrossRef] [PubMed]
- de Souza, A.G.; Dos Santos, N.M.A.; da Silva Torin, R.F.; dos Santos Rosa, D. Synergic antimicrobial properties of carvacrol essential oil and montmorillonite in biodegradable starch films. Int. J. Biol. Macromol. 2020, 164, 1737–1747. [Google Scholar] [CrossRef]
- Nassar, M.I.; Yassine, Y.M.; Elshamy, A.I.; El-Beih, A.A.; El-Shazly, M.; Singab, A.N.B. Essential oil and antimicrobial activity of aerial parts of Cyperus leavigatus L. (Family: Cyperaceae). J. Essent. Oil Bear. Plants 2015, 18, 416–422. [Google Scholar] [CrossRef]
- Duraipandiyan, V.; Al-Harbi, N.A.; Ignacimuthu, S.; Muthukumar, C. Antimicrobial activity of sesquiterpene lactones isolated from traditional medicinal plant, Costus speciosus (Koen ex. Retz.) Sm. BMC Complement. Altern. Med. 2012, 12, 1–6. [Google Scholar] [CrossRef]
- Labed, F.; Masullo, M.; Mirra, V.; Nazzaro, F.; Benayache, F.; Benayache, S.; Piacente, S. Amino acid-sesquiterpene lactone conjugates from the aerial parts of Centaurea pungens and evaluation of their antimicrobial activity. Fitoterapia 2019, 133, 51–55. [Google Scholar] [CrossRef]
- Noriega, P.; Ballesteros, J.; De la Cruz, A.; Veloz, T. Chemical composition and preliminary antimicrobial activity of the hydroxylated sesquiterpenes in the essential oil from Piper barbatum kunth leaves. Plants 2020, 9, 211. [Google Scholar] [CrossRef]
- Mirjalili, M.; Hadian, J.; Aliahmadi, A.; Kanani, M.; Sonboli, A. Chemical composition and in vitro antimicrobial activity of the essential oil of Cyclotrichium leucotrichum from Iran. Nat. Prod. Res. 2013, 27, 934–937. [Google Scholar] [CrossRef]
- El-Shazly, A.; Dorai, G.; Wink, M. Composition and antimicrobial activity of essential oil and hexaneðether extract of Tanacetum santolinoides (DC.) Feinbr. and Fertig. Zeitschrift für Naturforschung C 2002, 57, 620–623. [Google Scholar] [CrossRef]
- Laouer, H.; Hirèche-Adjal, Y.; Prado, S.; Boulaacheb, N.; Akkal, S.; Singh, G.; Singh, P.; Isidorov, V.A.; Szczepaniak, L. Chemical composition and antimicrobial activity of essential oil of Bupleurum montanum and B. plantagineum. Nat. Prod. Commun. 2009, 4, 1605–1610. [Google Scholar] [CrossRef]
- Yousefbeyk, F.; Gohari, A.R.; Sourmaghi, M.H.S.; Amini, M.; Jamalifar, H.; Amin, M.; Golfakhrabadi, F.; Ramezani, N.; Amin, G. Chemical composition and antimicrobial activity of essential oils from different parts of Daucus littoralis Smith subsp. hyrcanicus Rech. f. J. Essent. Oil Bear. Plants 2014, 17, 570–576. [Google Scholar] [CrossRef]
- Saidana, D.; Mahjoub, M.; Boussaada, O.; Chriaa, J.; Chéraif, I.; Daami, M.; Mighri, Z.; Helal, A. Chemical composition and antimicrobial activity of volatile compounds of Tamarix boveana (Tamaricaceae). Microbiol. Res. 2008, 163, 445–455. [Google Scholar] [CrossRef]
- Nafis, A.; Kasrati, A.; Jamali, C.A.; Custódio, L.; Vitalini, S.; Iriti, M.; Hassani, L. A Comparative study of the in vitro antimicrobial and synergistic effect of essential oils from Laurus nobilis L. and Prunus armeniaca L. from Morocco with Antimicrobial drugs: New approach for health promoting products. Antibiotics 2020, 9, 140. [Google Scholar] [CrossRef] [PubMed]
- Dehghan, G.; Solaimanian, R.; Shahverdi, A.R.; Amin, G.; Abdollahi, M.; Shafiee, A. Chemical composition and antimicrobial activity of essential oil of Ferula szovitsiana DC. Flavour Fragr. J. 2007, 22, 224–227. [Google Scholar] [CrossRef]
- Ghaneian, M.T.; Ehrampoush, M.H.; Jebali, A.; Hekmatimoghaddam, S.; Mahmoudi, M. Antimicrobial activity, toxicity and stability of phytol as a novel surface disinfectant. Environ. Health Eng. Manag. J. 2015, 2, 13–16. [Google Scholar]
- Kačániová, M.; Galovičová, L.; Ivanišová, E.; Vukovic, N.L.; Štefániková, J.; Valková, V.; Borotová, P.; Žiarovská, J.; Terentjeva, M.; Felšöciová, S. Antioxidant, antimicrobial and antibiofilm activity of coriander (Coriandrum sativum L.) essential oil for its application in foods. Foods 2020, 9, 282. [Google Scholar] [CrossRef]
- Chaudhary, S.A. Flora of the Kingdom of Saudi Arabia (Vol. 1); Ministry of Agriculture and Water: Riyadh, Saudi Arabia, 1999. [Google Scholar]
- Collenette, S. Wildflowers of Saudi Arabia; National Commission for Wildlife Conservation and Development (NCWCD): Riyadh, Saudi Arabia, 1999. [Google Scholar]
- Tackholm, V. Students’ Flora of Egypt; Cairo University Press: Cairo, Egypt, 1974. [Google Scholar]
- Boulos, L. Flora of Egypt (Vol. 3); Al Hadara Publishing: Cairo, Egypt, 2002. [Google Scholar]
- Lorian, V. Antibiotics in Laboratory Medicine; Lippincott Williams & Wilkins: Baltimore, PA, USA, 2005. [Google Scholar]
- Ronald, M. Microbiologia; Compania editorial continental SA de CV: Mexico, Mexico, 1990; Volume 505. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Rt. | KI | Compound Name | Conc. (%) | Identification | ||
|---|---|---|---|---|---|---|---|
| Exp | Lit | Saudi Arabia | Egypt | ||||
| Oxygenated Monoterpenes | |||||||
| 1 | 7.06 | 1030 | 1031 | Eucalyptol | 0.37 ± 0.03 | 0.21 ± 0.01 | a & b |
| 2 | 11.12 | 1123 | 1122 | α-Cyclocitral | 0.15 ± 0.02 | --- | a & b |
| 3 | 11.45 | 1139 | 1137 | trans-Pinocarveol | 0.88 ± 0.03 | --- | a & b |
| 4 | 12.00 | 1143 | 1143 | Camphor | 1.50 ± 0.06 | --- | a & b |
| 5 | 12.75 | 1146 | 1145 | Verbenol | 0.33 ± 0.02 | --- | a & b |
| 6 | 13.48 | 1189 | 1189 | 4-Terpineol | 0.16 ± 0.02 | --- | a & b |
| 7 | 13.61 | 1201 | 1202 | Safranal | 0.35 ± 0.02 | --- | a & b |
| 8 | 15.74 | 1235 | 1234 | trans-Chrysanthenyl acetate | 12.33 ± 0.09 | 1.09 ± 0.04 | a & b |
| 9 | 16.95 | 1336 | 1338 | α-Terpinyl propionate | 0.41 ± 0.02 | --- | a & b |
| 10 | 25.03 | 1453 | 1455 | Neryl acetone | --- | 0.61 ± 0.03 | a & b |
| Oxygenated Sesquiterpenes | |||||||
| 11 | 24.10 | 1499 | 1501 | β-Himachalene | 0.30 ± 0.01 | --- | a & b |
| 12 | 24.32 | 1439 | 1440 | α-Guaiene | 0.17 ± 0.01 | --- | a & b |
| 13 | 24.40 | 1511 | 1512 | Germacrene D-4-ol | 0.24 ± 0.02 | --- | a & b |
| 14 | 24.90 | 1517 | 1516 | 6-Epishyobunol | 0.28 ± 0.01 | --- | a & b |
| 15 | 25.53 | 1535 | 1533 | Nerolidol | 0.64 ± 0.03 | --- | a & b |
| 16 | 26.80 | 1551 | 1554 | Diepicedrene-1-oxide | 1.95 ± 0.06 | --- | a & b |
| 17 | 27.14 | 1558 | 1557 | Dihydro-α-agarofuran | 0.86 ± 0.04 | --- | a & b |
| 18 | 27.73 | 1563 | 1561 | Hexahydrofarnesol | 0.23 ± 0.02 | --- | a & b |
| 19 | 27.95 | 1564 | 1562 | Epiglobulol | 0.28 ± 0.01 | --- | a & b |
| 20 | 28.29 | 1567 | 1568 | Palustrol | 0.40 ± 0.03 | --- | a & b |
| 21 | 28.52 | 1575 | 1580 | Caryophyllene oxide | --- | 0.41 ± 0.02 | a & b |
| 22 | 28.77 | 1578 | 1579 | Spathulenol | 0.46 ± 0.02 | --- | a & b |
| 23 | 28.94 | 1586 | 1584 | Viridiflorol | 2.47 ± 0.07 | --- | a & b |
| 24 | 29.58 | 1588 | 1588 | Calarene epoxide | 0.21 ± 0.01 | --- | a & b |
| 25 | 29.66 | 1594 | 1594 | Isoaromadendrene epoxide | 0.27 ± 0.01 | --- | a & b |
| 26 | 29.85 | 1596 | 1597 | α-Cedrol | 0.56 ± 0.03 | --- | a & b |
| 27 | 30.73 | 1604 | 1606 | Cedrenol | 0.59 ± 0.02 | --- | a & b |
| 28 | 30.94 | 1619 | 1622 | Humulane-1,6-dien-3-ol | 0.78 ± 0.01 | --- | a & b |
| 29 | 31.09 | 1625 | 1625 | Aromadendrene oxide (1) | 0.45 ± 0.02 | --- | a & b |
| 30 | 31.39 | 1627 | 1628 | 4-epi-cubedol | 1.82 ± 0.06 | --- | a & b |
| 31 | 31.81 | 1632 | 1631 | 1,4-trans-1,7-cis-Acorenone | 7.62 ± 0.05 | --- | a & b |
| 32 | 32.21 | 1638 | 1638 | Hinesol | 13.50 ± 0.08 | --- | a & b |
| 33 | 32.48 | 1649 | 1651 | β-Eudesmol | 0.60 ± 0.02 | --- | a & b |
| 34 | 33.24 | 1657 | 1656 | α-Acorenol | 1.95 ± 0.05 | 0.35 ± 0.02 | a & b |
| 35 | 33.76 | 1669 | 1671 | Cedr-8-en-15-ol | 0.36 ± 0.02 | --- | a & b |
| 36 | 34.23 | 1689 | 1687 | Cedr-8-en-13-ol | 0.78 ± 0.03 | 0.45 ± 0.01 | a & b |
| 37 | 36.89 | 1691 | 1692 | Juniper camphor | 0.18 ± 0.01 | 0.40 ± 0.02 | a & b |
| 38 | 38.66 | 1885 | 1885 | (8S,14)-Cedrandiol | 0.43 ± 0.02 | 1.38 ± 0.04 | a & b |
| 39 | 45.69 | 1922 | 1925 | (E,E)-Farnesyl acetone | 0.16 ± 0.01 | --- | a & b |
| 40 | 40.79 | 2045 | 2005 | Isochiapin B | 0.56 ± 0.02 | 1.03 ± 0.05 | a & b |
| Sesquiterpenes Hydrocarbons | |||||||
| 41 | 19.26 | 1351 | 1351 | α-Cubebene | 2.19 ± 0.06 | --- | a & b |
| 42 | 20.48 | 1376 | 1378 | α-Copaene | 0.84 ± 0.02 | --- | a & b |
| 43 | 21.26 | 1409 | 1409 | α-Cedrene | 0.44 ± 0.03 | --- | a & b |
| 44 | 21.71 | 1410 | 1412 | α-Gurjunene | 0.32 ± 0.01 | --- | a & b |
| 45 | 22.29 | 1418 | 1418 | trans-Caryophyllene | 1.44 ± 0.07 | 3.07 ± 0.06 | a & b |
| 46 | 22.78 | 1429 | 1429 | cis-Thujopsene | 0.61 ± 0.03 | 0.83 ± 0.02 | a & b |
| 47 | 22.96 | 1462 | 1460 | α-Humulene | 0.26 ± 0.02 | --- | a & b |
| 48 | 23.55 | 1480 | 1483 | α-Muurolene | 0.15 ± 0.01 | --- | a & b |
| 49 | 23.81 | 1484 | 1486 | Germacrene-D | 0.19 ± 0.01 | --- | a & b |
| 50 | 23.95 | 1496 | 1489 | Aromadendrene | 0.43 ± 0.02 | --- | a & b |
| 51 | 24.64 | 1493 | 1496 | β-Muurolene | 0.56 ± 0.02 | --- | a & b |
| 52 | 25.16 | 1500 | 1502 | Bicyclogermacrene | --- | 4.37 ± 0.08 | a & b |
| 53 | 25.22 | 1517 | 1515 | α-Selinene | 0.95 ± 0.04 | --- | a & b |
| 54 | 25.63 | 1521 | 1524 | cis-Calamenene | 0.74 ± 0.03 | --- | a & b |
| 55 | 26.22 | 1532 | 1533 | γ-Cadinene | 1.86 ± 0.07 | --- | a & b |
| 56 | 26.39 | 1537 | 1537 | α-Cadinene | 3.31 ± 0.06 | --- | a & b |
| 57 | 26.56 | 1555 | 1557 | Junipene | 0.52 ± 0.03 | --- | a & b |
| 58 | 27.45 | 1548 | 1546 | α-Calacorene | 0.16 ± 0.01 | --- | a & b |
| Oxygenated Diterpenes | |||||||
| 59 | 44.31 | 1942 | 1944 | Isophytol | 0.25 ± 0.01 | 2.66 ± 0.08 | a & b |
| 60 | 46.99 | 1949 | 1950 | Phytol | 8.73 ± 0.09 | 38.02 ± 0.13 | a & b |
| 61 | 47.95 | 2201 | 2203 | trans-Geranyl geraniol | 0.80 ± 0.02 | 3.24 ± 0.08 | a & b |
| Diterpenes Hydrocarbons | |||||||
| 62 | 44.81 | 2064 | 2062 | Kaur-16-ene | 0.39 ± 0.03 | 0.90 ± 0.02 | a & b |
| Oxygenated Hydrocarbons | |||||||
| 63 | 35.57 | 1635 | 1636 | 1-Heptatriacontanol | 0.47 ± 0.03 | --- | a & b |
| 64 | 41.35 | 1754 | 1756 | Hexyl cinnamic aldehyde | 0.91 ± 0.04 | --- | a & b |
| 65 | 46.78 | 1927 | 1927 | Methyl palmitate | 2.86 ± 0.07 | 2.70 ± 0.05 | a & b |
| 66 | 47.23 | 2108 | 2109 | Linoleic acid, methyl ester | --- | 0.72 ± 0.03 | a & b |
| 67 | 47.69 | 2128 | 2128 | Methyl stearate | --- | 0.98 ± 0.04 | a & b |
| 68 | 47.78 | 2144 | 2145 | Z-7-Hexadecenal | 0.49 ± 0.03 | 2.10 ± 0.06 | a & b |
| 69 | 48.57 | 2152 | 2152 | Linoleic acid | 0.15 ± 0.01 | 6.36 ± 0.07 | a & b |
| 70 | 54.87 | 2161 | 2161 | Oleic Acid | 0.59 ± 0.02 | 3.51 ± 0.09 | a & b |
| Non-oxygenated Hydrocarbons | |||||||
| 71 | 32.55 | 1900 | 1901 | n-Nonadecane | --- | 0.45 ± 0.01 | a & b |
| 72 | 39.99 | 2000 | 2000 | n-Eicosane | --- | 0.67 ± 0.02 | a & b |
| 73 | 43.24 | 2100 | 2101 | n-Heneicosane | --- | 1.30 ± 0.05 | a & b |
| 74 | 49.33 | 2200 | 2200 | n-Docosane | --- | 6.86 ± 0.11 | a & b |
| 75 | 52.45 | 2300 | 2303 | n-Tricosane | 0.84 ± 0.03 | --- | a & b |
| 76 | 57.85 | 2500 | 2502 | n-Pentacosane | 0.76 ± 0.02 | 6.31 ± 0.07 | a & b |
| Aromatics | |||||||
| 77 | 19.93 | 1355 | 1353 | 1,1,6-Trimethyl-1,2-dihydronaphthalene | 0.37 ± 0.02 | --- | a & b |
| 78 | 36.99 | 1438 | 1436 | Bolandiol | 0.17 ± 0.01 | 0.15 ± 0.01 | a |
| 79 | 38.42 | 1845 | 1843 | Myristicin | 6.13 ± 0.08 | 2.09 ± 0.05 | a & b |
| 80 | 40.73 | 1457 | 1457 | Myristic acid | 0.50 ± 0.02 | --- | a & b |
| 81 | 44.51 | 2277 | 2478 | Ethyl iso-allocholate | --- | 1.58 ± 0.06 | a |
| Carotenoid-derived compounds | |||||||
| 82 | 17.41 | 1284 | 1283 | Dihydroedulan II | 0.16 ± 0.01 | --- | a & b |
| 83 | 20.94 | 1351 | 1354 | α-Damascenone | 1.42 ± 0.03 | 0.71 ± 0.01 | a & b |
| 84 | 23.67 | 1426 | 1426 | trans-α-Ionone | 2.00 ± 0.05 | 2.91 ± 0.04 | a & b |
| 85 | 32.07 | 1473 | 1472 | α-Iso methyl ionone | 0.38 ± 0.03 | 0.38 ± 0.02 | a & b |
| 86 | 33.01 | 1518 | 1519 | Methyl-α-ionone | 0.19 ± 0.02 | --- | a & b |
| Total identified | 99.11 | 98.80 | |||||
| Strains | EO Concentration (µg mL−1) | MIC b (µg mL−1) | Antibiotic (10 µg mL−1) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 2.50 | 1.25 | 0.625 | 0.312 | 0.156 | 0.078 | ||||
| Bacterial | Gentamycin | ||||||||
| S.aureus | SA | 35.20 ± 0.60 a | 34.20 ± 0.95 | 33.20 ± 1.15 | 31.80 ± 1.00 | 30.10 ± 0.50 | 17.20 ± 0.80 | 15.75 | 36.00 ± 1.16 |
| Eg | 23.00 ± 1.40 | 22.10 ± 0.77 | 20.20 ± 1.35 | 19.20 ± 1.30 | 16.00 ± 1.10 | 14.40 ± 0.80 | 15.75 | ||
| S. pyogenes | SA | 22.00 ± 0.82 | 21.10 ± 0.85 | 18.40 ± 0.30 | 17.98 ± 0.89 | 16.00 ± 0.16 | 14.10 ± 1.20 | 20.58 | 30.00 ± 0.94 |
| Eg | 22.80 ± 1.0 | 21.10 ± 1.43 | 19.00 ± 0.82 | 16.68 ± 0.80 | 14.90 ± 0.87 | 12.45 ± 0.90 | 15.79 | ||
| S.epidermidis | SA | 20.00 ± 1.00 | 31.20 ± 0.90 | 28.10 ± 0.10 | 26.00 ± 0.65 | 24.60 ± 0.50 | 16.30 ± 0.98 | 15.75 | 34.00 ± 0.84 |
| Eg | 33.00 ± 1.25 | 18.00 ± 1.00 | 17.20 ± 1.00 | 15.90 ± 1.00 | 14.68 ± 0.77 | 10.60 ± 0.35 | 15.81 | ||
| S. typhi | SA | 37.00 ± 0.88 | 31.80 ± 0.65 | 29.20 ± 0.30 | 27.30 ± 0.25 | 25.70 ± 0.35 | 15.10 ± 0.73 | 20.58 | 37.00 ± 0.88 |
| Eg | 32.30 ± 0.7 | 35.10 ± 0.85 | 33.78 ± 0.85 | 32.80 ± 0.70 | 30.90 ± 1.47 | 16.20 ± 1.20 | 15.75 | ||
| E. coli | SA | 21.00 ± 1.10 | 9.10 ± 0.37 | 7.98 ± 0.10 | 6.97 ± 0.25 | 5.60 ± 0.19 | 6.20 ± 0.33 | 23.25 | 21.00 ± 1.10 |
| Eg | 10.00 ± 0.52 | 20.20 ± 1.10 | 19.95 ± 1.07 | 17.20 ± 1.10 | 14.90 ± 0.56 | 8.50 ± 0.80 | 15.75 | ||
| Shigella spp. | SA | 28.00 ± 1.10 | 37.00 ± 0.15 | 35.60 ± 0.40 | 33.60 ± 0.50 | 30.80 ± 0.35 | 17.50 ± 0.40 | 20.58 | 28.00 ± 0.60 |
| Eg | 39.00 ± 0.7 | 26.20 ± 1.15 | 25.90 ± 1.10 | 25.00 ± 1.10 | 23.80 ± 0.64 | 11.00 ± 1.50 | 15.88 | ||
| P. aeruginosa | SA | 31.00 ± 0.90 | 26.80 ± 0.95 | 25.20 ± 0.20 | 23.40 ± 0.25 | 30.10 ± 0.50 | 14.35 ± 0.70 | 31.25 | 31.00 ± 0.90 |
| Eg | 28.00 ± 1.00 | 29.20 ± 0.90 | 27.00 ± 0.90 | 25.90 ± 0.90 | 23.50 ± 0.92 | 12.50 ± 1.30 | 23.25 | ||
| Fungal | Amphotericin | ||||||||
| T. shoenlenii | SA | 31.00 ± 0.90 | 29.20 ± 0.90 | 28.10 ± 1.10 | 25.90 ± 1.10 | 23.00 ± 1.10 | 15.00 ± 0.80 | 21.75 | 21.00 ± 1.10 |
| Eg | 31.30 ± 1.15 | 30.10 ± 1.10 | 27.00 ± 0.90 | 25.90 ± 0.90 | 23.50 ± 0.96 | 12.50 ± 1.30 | 15.75 | ||
| A. fumigatus | SA | MI | MI | MI | MI | MI | 9.00 ± 1.50 | 21.30 | 28.00 ± 0.60 |
| Eg | MI | MI | MI | MI | MI | 35.00 ± 1.10 | 15.79 | ||
Sample Availability: Samples of the compounds are not available from the authors. | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Rowaily, S.L.; Abd-ElGawad, A.M.; Assaeed, A.M.; Elgamal, A.M.; Gendy, A.E.-N.G.E.; Mohamed, T.A.; Dar, B.A.; Mohamed, T.K.; Elshamy, A.I. Essential Oil of Calotropis procera: Comparative Chemical Profiles, Antimicrobial Activity, and Allelopathic Potential on Weeds. Molecules 2020, 25, 5203. https://doi.org/10.3390/molecules25215203
Al-Rowaily SL, Abd-ElGawad AM, Assaeed AM, Elgamal AM, Gendy AE-NGE, Mohamed TA, Dar BA, Mohamed TK, Elshamy AI. Essential Oil of Calotropis procera: Comparative Chemical Profiles, Antimicrobial Activity, and Allelopathic Potential on Weeds. Molecules. 2020; 25(21):5203. https://doi.org/10.3390/molecules25215203
Chicago/Turabian StyleAl-Rowaily, Saud L., Ahmed M. Abd-ElGawad, Abdulaziz M. Assaeed, Abdelbaset M. Elgamal, Abd El-Nasser G. El Gendy, Tarik A. Mohamed, Basharat A. Dar, Tahia K. Mohamed, and Abdelsamed I. Elshamy. 2020. "Essential Oil of Calotropis procera: Comparative Chemical Profiles, Antimicrobial Activity, and Allelopathic Potential on Weeds" Molecules 25, no. 21: 5203. https://doi.org/10.3390/molecules25215203
APA StyleAl-Rowaily, S. L., Abd-ElGawad, A. M., Assaeed, A. M., Elgamal, A. M., Gendy, A. E.-N. G. E., Mohamed, T. A., Dar, B. A., Mohamed, T. K., & Elshamy, A. I. (2020). Essential Oil of Calotropis procera: Comparative Chemical Profiles, Antimicrobial Activity, and Allelopathic Potential on Weeds. Molecules, 25(21), 5203. https://doi.org/10.3390/molecules25215203