
Assessment of Insecticidal Activity of Benzylisoquinoline Alkaloids from Chilean Rhamnaceae Plants against Fruit-Fly Drosophila melanogaster and the Lepidopteran Crop Pest Cydia pomonella

 , , and
, , and
Abstract
1. Introduction
2. Results
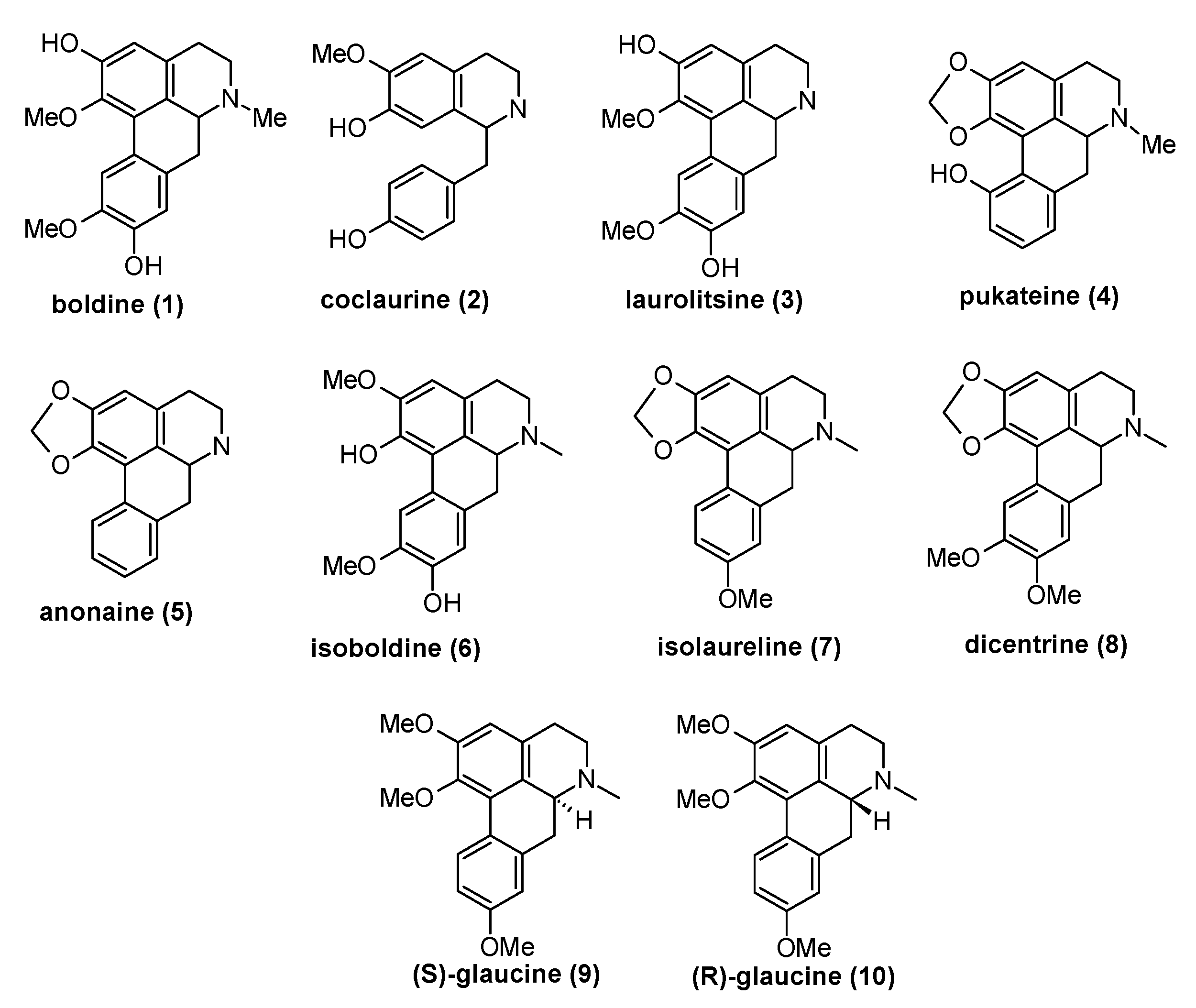
2.1. Phytochemical Analysis
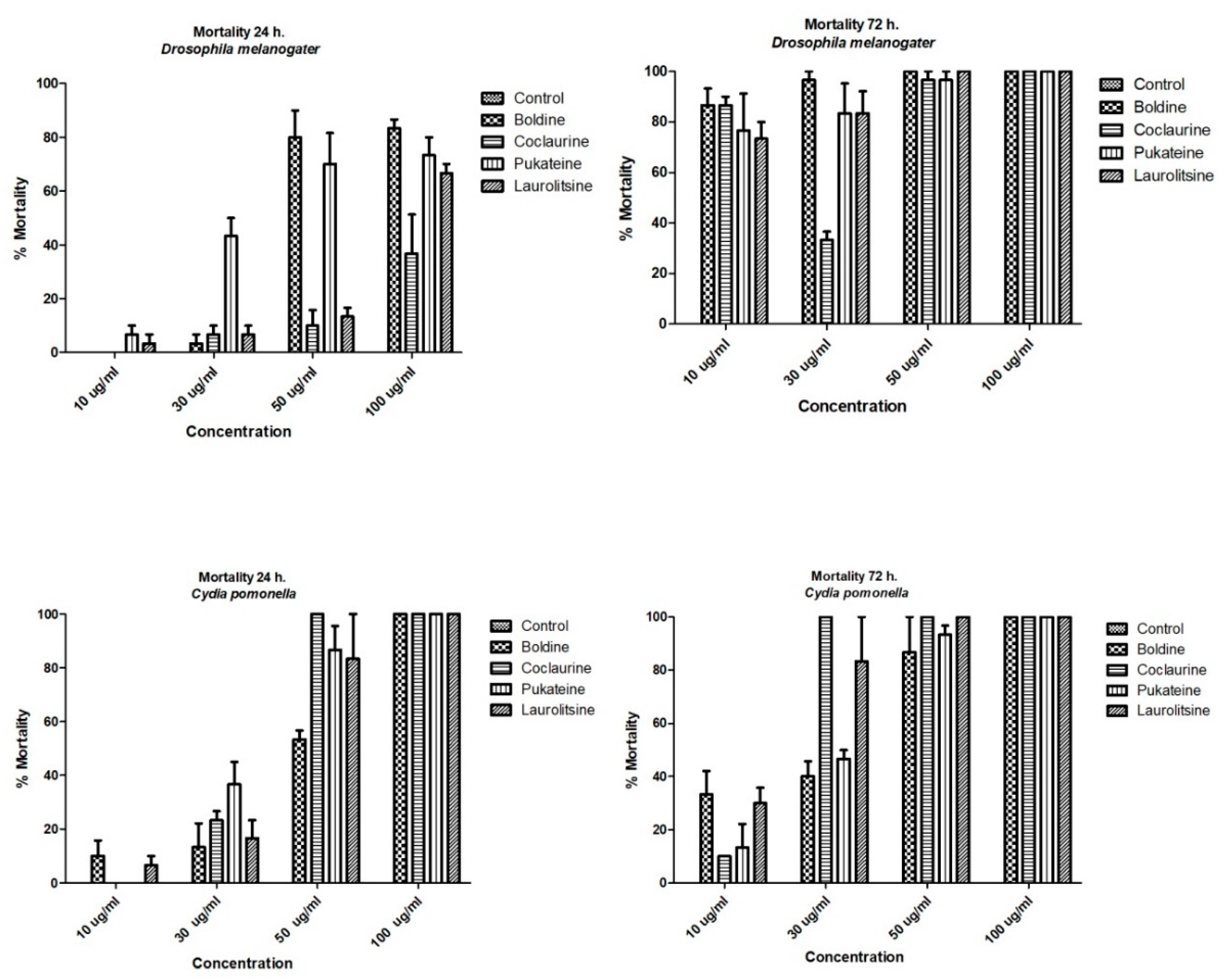
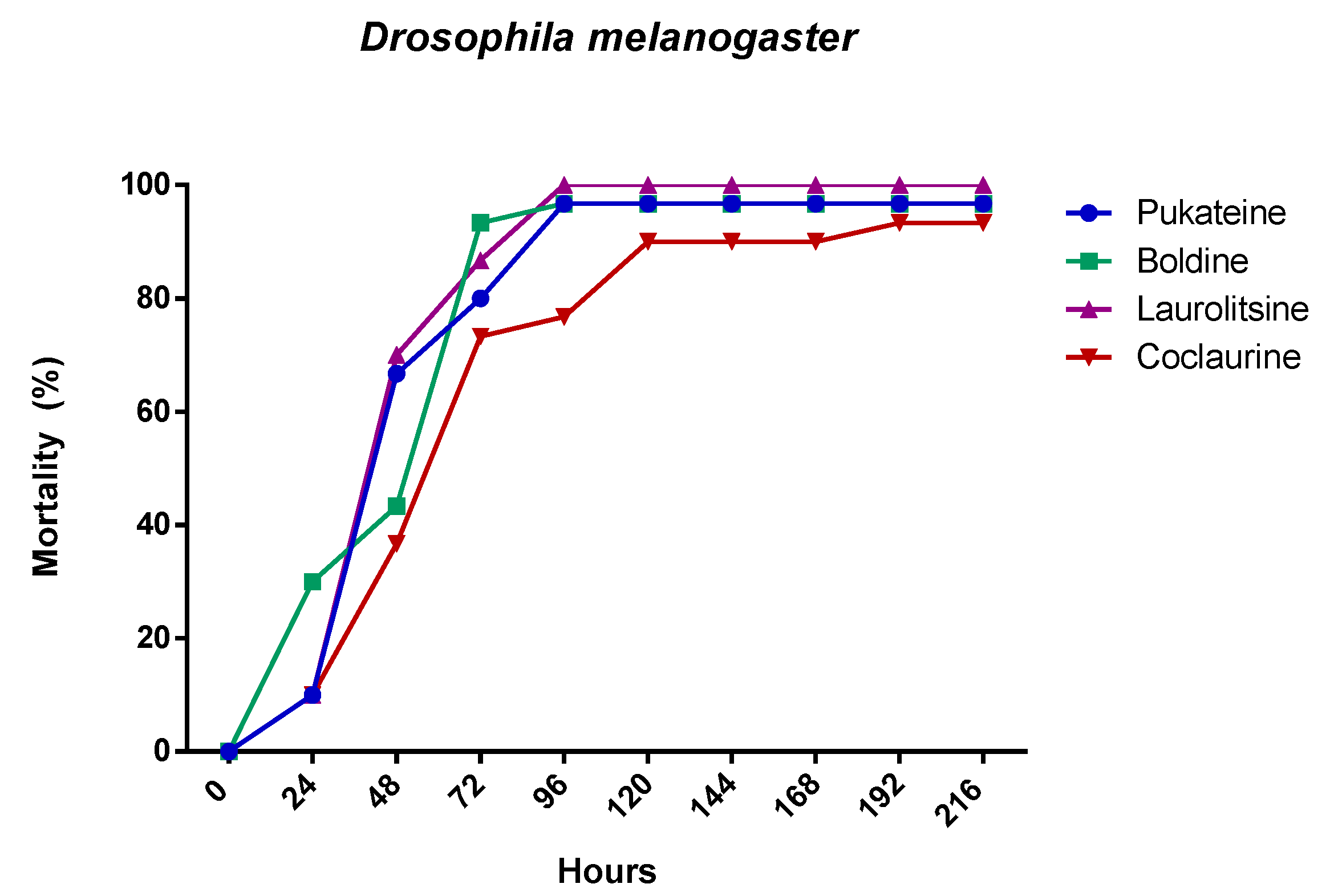
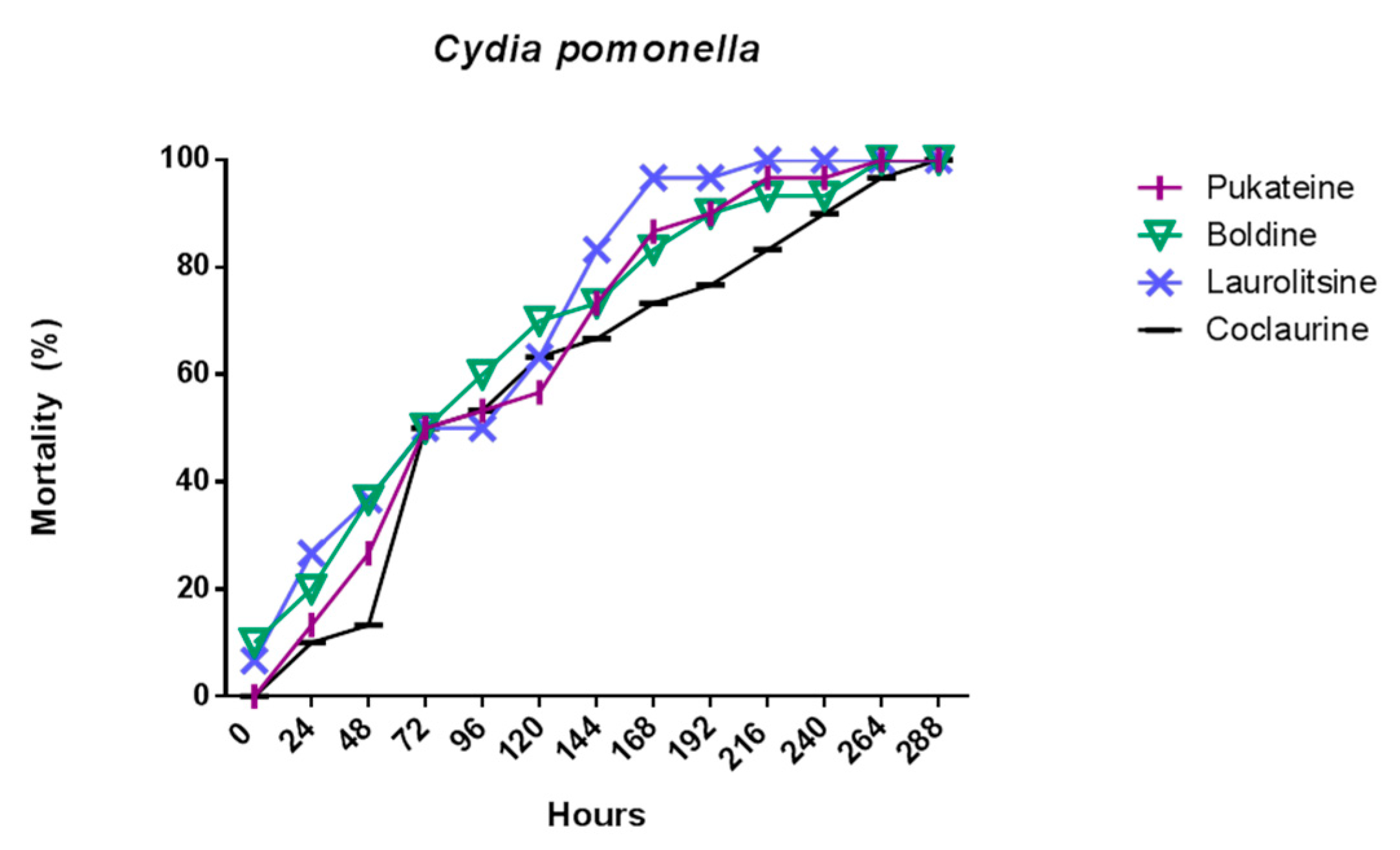
2.2. Insecticidal Activity
2.2.1. Larval Toxicity of the Alkaloids
2.2.2. Influence of the Alkaloids on Insect Growth
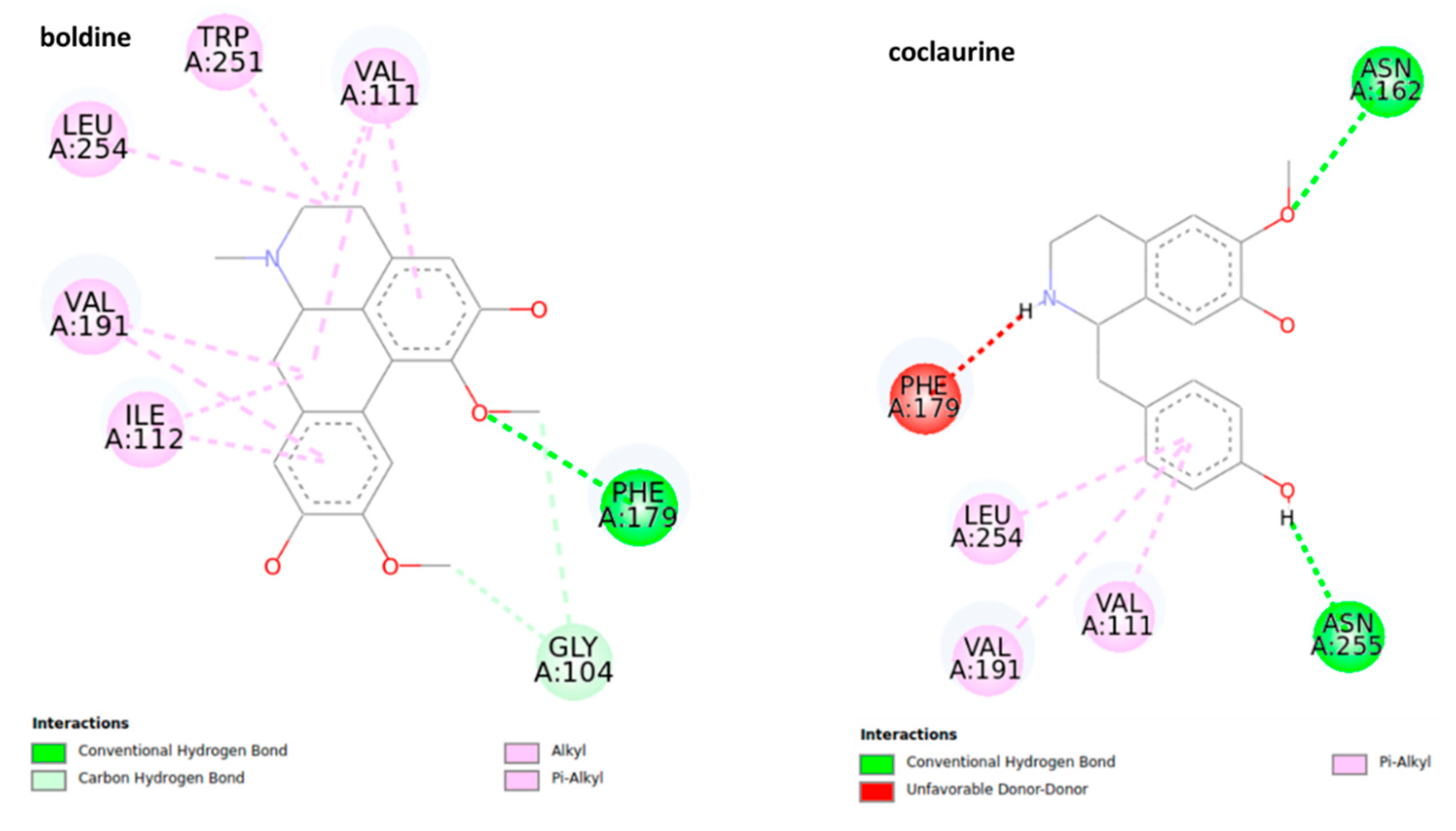
2.3. Docking Study
2.3.1. Octopamine Receptor
2.3.2. Ecdysteroid Receptor
3. Discussion
4. Materials and Methods
4.1. Equipment and General Experimental Procedures
4.2. Plant Material
4.3. Extraction and Isolation
4.4. Insect Bioassays
4.5. Bioassay for Insecticidal Activity against Larvae of D. melanogaster
4.6. Bioassays for Insecticidal Activity against Larvae of C. pomonella
4.7. Growth Regulation Assay
4.8. Ligand Construction and Docking Studies on the Ecdysone Receptor EcR and OAMB Receptor
4.8.1. Receptor Modeling and Ligand Preparation
4.8.2. Molecular Docking
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- ODEPA. Oficina de Estudios y Políticas Agrarias. Boletín de Frutas y Hortalizas Procesadas. 2019. Available online: https://www.odepa.gob.cl (accessed on 8 October 2020).
- Athanassiou, C.G.; Kavallieratos, N.G.; Sciarretta, A.; Palyvos, N.E.; Trematerra, P. Spatial associations of insects and mites in stored wheat. J. Econ. Entomol. 2011, 104, 1752–1764. [Google Scholar] [CrossRef]
- Emameh, R.Z.; Syrjänen, L.; Barker, H.; Supuran, C.T.; Parkkila, S. Drosophila melanogaster: A model organism for controlling Dipteran vectors and pests. J. Enzym. Inhib. Med. Chem. 2015, 30, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Wilson, T. Drosophila melanogaster (Diptera: Drosophilidae): A Model Insect for Insecticide Resistance Studies. J. Econ. Entomol. 1988, 81, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Benhalima, H.; Chaudhry, M.Q.; Mills, K.A.; Price, N.R. Phosphine resistance instored-product insects collected from various grain storage facilities in Morocco. J. Stored Prod. Res. 2004, 40, 241–249. [Google Scholar] [CrossRef]
- Bughio, F.M.; Wilkins, R.M. Influence of malathion resistance status on survivaland growth of Tribolium castaneum, when fed on flour from insect resistant and susceptible grain rice cultivars. J. Stored Prod. Res. 2005, 40, 65–75. [Google Scholar] [CrossRef]
- Kedia, A.; Prakash, B.; Mishra, P.K.; Singh, P.; Dubey, N.K. Botanicals as ecofriendly biorational alternatives of synthetic pesticides against Callosobruchus spp. (Coleoptera: Bruchidae)—A review. J. Food Sci. Technol. 2015, 52, 1239–1257. [Google Scholar] [CrossRef]
- Cespedes, C.L.; Salazar, J.R.; Alarcon, J. Chemistry and biological activities of Calceolaria spp. (Calceolariaceae: Scrophulariaceae). Phytochem. Rev. 2013, 12, 733–749. [Google Scholar] [CrossRef]
- Cespedes, C.L.; Aqueveque, P.M.; Avila, J.G.; Alarcon, J.; Kubo, I. New advances in chemical defenses of plants: Researches in Calceolariaceae. Phytochem. Rev. 2015, 14, 367–380. [Google Scholar] [CrossRef]
- Cespedes, C.L.; Lina-Garcia, L.; Kubo, I.; Salazar, J.R.; Ariza-Castolo, A.; Alarcon, J.; Aqueveque, P.M.; Werner, E.; Seigler, D.S. Calceolaria integrifolia s.l. complex, reduces feeding and growth of Acanthoscelides obtectus, and Epilachna varivestis. A new source of bioactive compounds against dry bean pests. Ind. Crop. Prod. 2016, 89, 257–267. [Google Scholar] [CrossRef]
- Muñoz, E.; Lamilla, C.; Marin, J.C.; Alarcon, J.; Cespedes, C.L. Antifeedant, insect growth regulatory and insecticidal effects of Calceolaria talcana (Calceolariaceae) on Drosophila melanogaster and Spodoptera frugiperda. Ind. Crop. Prod. 2013, 42, 137–144. [Google Scholar] [CrossRef]
- Rios, J.L.; Recio, M.C. Medicinal plants and antimicrobial activity. J. Ethnopharmacol. 2005, 100, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Isman, M.B. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Ann. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef] [PubMed]
- Sosa, M.E.; Tom, C.E. Plant secondary metabolites from Argentinean semiarid lands: Bioactivity against insects. Phytochem. Rev. 2008, 7, 3–24. [Google Scholar] [CrossRef]
- Svetaz, L.; Derita, M.; Rodriguez, M.V.; Postigo, A.; Butassi, E.; Castelli, M.V.; Sortino, M.; Petenatti, E.; Zacchino, S. Antifungal compounds from Latin American plants. In Antifungal Metabolites from Plants; Razzaghi-Abyanch, M., Rai, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 3–26. [Google Scholar]
- Miresmailli, S.; Isman, M.B. Botanical insecticides inspired by plant-herbivore chemical interaction. Trends Plants Sci. 2014, 19, 29–35. [Google Scholar] [CrossRef]
- Hagel, J.M.; Facchini, P.J. Benzylisoquinoline alkaloids metabolism: A century of Discovery and a Brave new world. Plant Cell Physiol. 2013, 54, 647–672. [Google Scholar] [CrossRef]
- Seigler, D.S. Plant Secondary Metabolism; Kluwer Academic Publishers: Boston, MA, USA, 1998; pp. 506–512. [Google Scholar]
- Martin, M.L.; Diaz, M.T.; Montero, M.J.; Prieto, P.; San Ramon, L.; Cortes, D. Antispasmodic activity of benzylisoquinoline alkaloids analogous to papaverine. Planta Med. 1993, 59, 63–67. [Google Scholar] [CrossRef]
- Fedurco, M.; Gregorová, J.; Šebrlová, K.; Kantorová, J.; Peš, O.; Baur, R.; Sigel, E.; Táborská, E. Modulatory Effects of Eschscholzia californica Alkaloids on Recombinant GABAA Receptors. Biochem. Res. Int. 2015, 2015, 617620. [Google Scholar] [CrossRef]
- Exley, R.; Iturriaga-Vasquez, P.; Lukas, R.J.; Sher, E.; Cassels, B.K.; Bermudez, I. Evaluation of benzyltetrahydroisoquinolines as ligands for neuronal nicotinic acetylcholine receptors. Br. J. Pharmacol. 2005, 146, 15–24. [Google Scholar] [CrossRef]
- Lai, J.H. Immunomodulatory effects and mechanisms of plant alkaloid tetrandrine in autoimmune diseases. Acta Pharmacol. Sin. 2005, 23, 1093–1101. [Google Scholar]
- Kwan, C.Y.; Achike, F.I. Tetrandrine and related bis-benzylisoquinoline alkaloids from medicinal herbs: Cardiovascular effects and mechanisms of action. Acta Pharmacol. Sin. 2002, 23, 1057–1068. [Google Scholar]
- Chulia, S.; Ivorra, M.D.; Martinez, S.; Elorriaga, M.; Valiente, M.; Noiguera, M.A.; Lugnier, C.; Advenier, C.; D’Ocon, P. Relationships between structure and vascular activity in a series of benzylisoquinolines. Br. J. Pharmacol. 1997, 122, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, T. Alkaloids. In Herbivores: Their Interactions with Secondary Plant Metabolites, 2nd ed.; Rosenthal, G.A., Berenbaum, M.R., Eds.; Academic Press: San Diego, CA, USA, 1991; Volume 1, pp. 79–121. [Google Scholar]
- Quiroz-Carreño, S.; Cespedes-Acuña, C.L.; Seigler, D.S.; Alarcón-Enos, J. Identification of structurally diverse alkaloids in Talguenea quinquinervia (Gill. et Hook) by liquid chromatography/electrospray ionization tandem mass spectroscopy and insecticidal activity. Phytochem. Anal. 2019, 30, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Alarcón, J.; Molina, S.; Villalobos, N.; Lillo, L.; Lamilla, C.; Cepedes, C.L.; Seigler, D.S. Insecticidal activity of Chilean Rhamnaceae: Talguenea quinquenervia (Gill. et Hook). Bol. Latinoam. Caribe Plant. Med. Aromat. 2011, 10, 389–394. [Google Scholar]
- Rauschenbach, I.Y.; Chentsova, N.A.; Alekseev, A.A.; Gruntenko, N.E.; Adonyeva, N.V.; Karpova, E.K.; Komarova, T.N.; Vasiliev, V.G.; Bownes, M. Dopamine and octopamine regulate 20-hydroxyecdysone level in vivo in Drosophila. Arch. Insect Biochem. Physiol. 2007, 65, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Evans, P.D.; Maqueira, B. Insect octopamine receptors: A new classification scheme based on studies of cloned Drosophila G-protein coupled receptors. Invertebr. Neurosci. 2005, 5, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Bogomolova, E.V.; Adonyeva, N.V.; Gruntenko, N.E.; Rauschenbach, I.Y. Effects of 20-hydroxyecdysone and juvenile hormone on octopamine metabolism in females of Drosophila. Arch. Insect Biochem. Physiol. 2009, 70, 244–253. [Google Scholar] [CrossRef]
- Niwa, Y.S.; Niwa, R. Transcriptional regulation of insect steroid hormone biosynthesis and its role in controlling timing of molting and metamorphosis. Dev. Growth Differ. 2016, 58, 94–105. [Google Scholar] [CrossRef]
- Riddiford, L.M.; Cherbas, P.; Truman, J.W. Ecdysone receptors and their biological actions. Vitam. Horm. 2000, 60, 1–73. [Google Scholar]
- Maqueira, B.; Chatwin, H.; Evans, P.D. Identification and characterization of a novel family of Drosophila β-adrenergic-like octopamine G-protein coupled receptors. J. Neurochem. 2005, 94, 547–560. [Google Scholar] [CrossRef]
- Correa, C.; Urzua, A.; Torres, R. 1,2,11-trimethoxynoraporphine from Discaria chacaye (G. DON)Tort. Boletín Soc. Chil. Química 1987, 32, 105–106. [Google Scholar]
- Pacheco, P.; Albonico, S.M.; Silva, M. Alkaloids, triterpenes and other constituents of Discaria crenata. Phytochemistry 1973, 12, 954–955. [Google Scholar] [CrossRef]
- Torres, R.; Delle Monache, F.; Marini, G.B. Alkaloids from Discaria serratifolia. J. Nat. Prod. 1979, 42, 430–431. [Google Scholar] [CrossRef]
- Rivera, A.; Urzua, A.; Torres, R. 1,2-dimethoxy-11-hydroxyaporphine from Discaria serratifolia var. Montana. J. Nat. Prod. 1984, 47, 1040–1041. [Google Scholar] [CrossRef]
- Sanchez, E.; Comin, J. Quaternary bases from Colletia spinossisima GMEL. Tetrahedron 1967, 23, 1139–1143. [Google Scholar]
- Castro-Saavedra, S.; Fuentes-Barros, G.; Tirapegui, C.; Acevedo-Fuentes, W.; Cassel, B.; Barriga, A.; Vilches-Herrera, M. Phytochemical analysis of alkaloids from the Chilean endemic tree Cryptocarya alba. J. Chil. Chem. Soc. 2016, 61, 3076–3080. [Google Scholar] [CrossRef]
- Fuentes-Barros, G.; Castro-Saavedra, S.; Liberona, L.; Acevedo-Fuentes, W.; Tirapegui, C.; Mattar, C.; Cassels, B.K. Variation of the alkaloid content of Peumus boldus (boldo). Fitoterapia 2018, 127, 179–185. [Google Scholar] [CrossRef]
- Dacanay, F.N.; Ladra, M.C.; Junio, H.A.; Nellas, R.B. Molecular Affinityof Mabolo Extracts to an Octopamine Receptor of a Fruit Fly. Molecules 2017, 22, 1677. [Google Scholar] [CrossRef]
- Hirashima, A.; Huang, H. Homology modeling, agonist binding site identification, and docking in octopamine receptor of Periplaneta americana. Comput. Biol. Chem. 2008, 32, 185–190. [Google Scholar] [CrossRef]
- Kastner, K.W.; Shoue, D.A.; Estiu, G.L.; Wolford, J.; Fuerst, M.F.; Markley, L.D.; Izaguirre, J.A.; McDowell, M. Characterization of the Anopheles gambiae octopamina receptor and discovery of potential agonists and antagonists using a combined computational-experimental approach. Malar. J. 2014, 13, 434. [Google Scholar] [CrossRef]
- Dhadialla, T.S.; Carlson, G.R.; Le, D.P. New insecticides with ecdysteroidal and juvenile hormone activity. Annu. Rev. Entomol. 1988, 43, 545–569. [Google Scholar] [CrossRef]
- Li, H.-T.; Wu, H.-M.; Chen, H.-L.; Liu, C.-M.; Chen, C.-Y. The Pharmacological activities of (-)-Anonaine. Molecules 2013, 18, 8257–8263. [Google Scholar] [CrossRef] [PubMed]
- Harper, S.H.; Potter, C.; Gillham, E.M. Annona species as insecticides. Ann. Appl. Biol. 1947, 34, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Crosby, D.G. Minor insecticides of plant origin. In Naturally Occurring Insecticides; Jacobson, M., Crosby, D.G., Eds.; Marcel Dekker: New York, NY, USA, 1971; pp. 177–239. [Google Scholar]
- Saxena, R.C.; Harshan, V.; Saxena, A.; Sukumaran, P.; Sharma, M.C.; Kumar, M.L. Larvicidal and chemosterilant activity of Annona squamosa alkaloids against Anopheles stephensi. J. Am. Mosq. Control Assoc. 1993, 9, 84–87. [Google Scholar] [PubMed]
- Indra, B.; Matsunaga, K.; Hoshino, O.; Susuki, M.; Ogasawara, H.; Ohizumi, Y. Structure activity realtionship studies with (±)-nantenine derivatives for α1-adrenoreceptor antagonist activity. Eur. J. Pharmacol. 2002, 437, 173–178. [Google Scholar] [CrossRef]
- Valiente, M.; D’Ocon, P.; Noguera, M.A.; Cassels, B.K.; Lugnier, C.; Ivorra, M.D. Vascular activity of (-)-anonaine, (-)-roemerine and (-)-pukateine. Tree natural 6-(R)-1,2-methylenedioxyaporphines with different affinities for α1-adrenoceptor subtypes. Planta Med. 2004, 70, 603–609. [Google Scholar] [CrossRef]
- Heng, H.L.; Chee, C.F.; Thy, C.K.; Tee, J.T.; Chin, S.P.; Herr, D.R.; Buckle, M.J.C.; Paterson, I.C.; Doughty, S.W.; Rahman, N.A.; et al. In vitro functional of isolaureline, dicentrine and glaucine enantiomers at 5-HT2 and α1 receptors. Chem. Biol. Drug Des. 2019, 93, 132–138. [Google Scholar] [CrossRef]
- Schwedes, C.; Tulsiani, S.; Carney, G.E. Ecdysone receptor expression and activity in adult Drosophila melanogaster. J. Insect Physiol. 2011, 57, 899–907. [Google Scholar] [CrossRef]
- Ohhara, Y.; Shimada-Niwa, Y.; Niwa, R.; Kayashima, Y.; Hayashi, Y.; Akagi, K.; Ueda, H.; Yamakawa-Kobayashi, K.; Kobayashi, S. Autocrine regulation of ecdysone synthesis by β3-octopamine receptor in the prothoracic gland is essential for Drosophila metamorphosis. Proc. Natl. Acad. Sci. USA 2015, 112, 1452–1457. [Google Scholar] [CrossRef]
- Bianchini, M.C.; Gularte, C.O.A.; Escoto, D.F.; Pereira, G.; Gayer, M.C.; Roehrs, R.; Soares, F.A.A.; Puntel, R.L. Peumus boldus (Boldo) Aqueous Extract Present Better Protective Effect than Boldine Against Manganese-Induced Toxicity in D. melanogaster. Neurochem. Res. 2016, 41, 2699–2707. [Google Scholar] [CrossRef]
- Klimaczewski, C.V.; Ecker, A.; Piccoli, B.; Aschner, M.; Barbosa, N.V.; Rocha, J.B.T. Peumus boldus attenuates copper-induced toxicity in Drosophila melanogaster. Biomed. Pharmacother. 2018, 97, 1–8. [Google Scholar] [CrossRef]
- Dajas-Bailador, F.A.; Asencio, M.; Bonilla, C.; Scorza, M.C.; Echeverry, C.; Reyes-Parada, M.; Silveira, R.; Protais, P.; Russell, G.; Cassels, B.K.; et al. Dopaminergic pharmacology and antioxidant properties of pukateine, a natural product lead for the design of agents increasing dopamine neurotransmission. Gen. Pharmacol. 1999, 32, 373–379. [Google Scholar] [CrossRef]
- Cazzamali, G.; Klaerke, D.A.; Grimmelikhuijzen, C.J. A new family of insect tyramine receptors. Biochem. Biophys. Res. Commun. 2005, 338, 1189–1196. [Google Scholar] [CrossRef] [PubMed]
- Lange, A.B. Tyramine: From octopamine precursor to neuroactive chemical in insects. Gen. Comp. Endocrinol. 2009, 15, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Claßen, G.; Scholz, H. Octopamine Shifts the Behavioral Response from Indecision to Approach or Aversion in Drosophila melanogaster. Front. Behav. Neurosci. 2018, 12, 131. [Google Scholar] [CrossRef]
- Gruntenko, N.; Rauschenbach, I. 20-Hydroxyecdysone, Juvenile Hormone and Biogenic Amines: Mechanisms of Interaction in Control of Drosophila Reproduction under Normal and Stressful Conditions. In Ecdysone: Structures and Functions; Springer: Dordrecht, The Netherlands, 2009; pp. 317–332. [Google Scholar]
- Li, H.-M.; Jiang, H.-B.; Gui, S.-H.; Liu, X.-Q.; Liu, H.; Lu, X.-P.; Smagghe, G.; Wang, J.-J. Characterization of a β-Adrenergic-Like Octopamine Receptor in the Oriental Fruit Fly, Bactrocera dorsalis (Hendel). Int. J. Mol. Sci. 2016, 17, 1577. [Google Scholar] [CrossRef]
- Li, F.; Li, K.; Wu, L.J.; Fan, Y.L.; Liu, T.X. Role of Biogenic Amines in Oviposition by the Diamondback Moth, Plutella xylostella L. Front. Physiol. 2020, 11, 475. [Google Scholar] [CrossRef]
- Beggs, K.T.; Tyndall, J.D.A.; Mercer, A.R. Honey bee dopamine and octopamine receptors linked to intracellular calcium signaling have a close phylogenetic and pharmacological relationship. PLoS ONE 2011, 6. [Google Scholar] [CrossRef]
- Kang, X.-L.; Zhang, J.-Y.; Wang, D.; Zhao, Y.-M.; Han, X.-L.; Wang, J.-X.; Zhao, X.-F. The steroid hormone 20-hydroxyecdysone binds to dopamine receptor to repress lepidopteran insect feeding and promote pupation. PLoS Genet. 2019, 15, e1008331. [Google Scholar] [CrossRef] [PubMed]
- Wada, K.; Munakata, K. Naturally occurring insect control chemicals. Isoboldine, a feeding inhibitor, and cocculolidine, an insecticide in the leaves of Cocculus trilobus DC. J. Agric. Food Chem. 1968, 16, 471–474. [Google Scholar] [CrossRef]
- Ivorra, M.D.; Chulia, S.; Lugnier, C.; D’Ocon, M.P. Selective action of two aporphines at a1-adrenoceptors and potential-operated Ca2+ channels. Eur. J. Pharmacol. 1993, 231, 165–174. [Google Scholar] [CrossRef]
- Ivorra, M.D.; Martinez, F.; Serrano, A.; D’Ocon, P. Different mechanism of relaxation induced by aporphine alkaloids in rat uterus. J. Pharm. Pharmacol. 1993, 45, 439–443. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, M.; Nakamura, Y.; Ishikawa, Y. Insecticidal sesquiterpene from Alpinia oxyphylla against Drosophila melanogaster. J. Agric Food Chem. 2009, 48, 3639–3641. [Google Scholar] [CrossRef]
- Piskroski, R.; Dorn, S. How the oligophage codling moth Cydia pomonella survives on walnut despite its secondary metabolites juglone. J. Insect Physiol. 2011, 57, 744–750. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.L.; Chaudhuri, S.K.; Kubo, I. Quantification of insect growth and its use in screening of naturally occurring insect control agents. J. Chem. Ecol. 1993, 19, 1109–1118. [Google Scholar] [CrossRef] [PubMed]
- MarvinSketch (Version 16.8.15, Calculation Module Developed by ChemAxon). Available online: http://www.chemaxon.com/products/marvin/marvinsketch/ (accessed on 3 October 2017).
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Mortality after 24 h (%) 10 µg/mL | Cumulative Mortality a,b,c (%) | Weight Gained (%) | Length Gained (%) | Pupation % | Mean Pupation Time (days) | % IG | Emergence (%) |
|---|---|---|---|---|---|---|---|---|
| Control | 0 | 0 | 100 | 100 | 100 | 8 | 100 | 100 |
| Boldine 1 | 0 | 82.4 ± 3.4 a,b,c | 11.2 ± 0.07 | 68.6 | 3.3 | 14 | 3.3 | 3.3 |
| Coclaurine 2 | 0 | 73.0 ± 3.4 a,b,c | 42.9 ± 0.06 | 72.9 | 6.6 | 12 | 6.6 | 6.6 |
| Laurolitsine 3 | 3.3 | 100 ± 0.0 a,b,c | 0.0 ± 0.0 | 0 | 0 | 0 | 0 | 0 |
| Pukateine 4 | 10 | 83.8 ± 2.1 a,b,c | 77.5 ± 0.03 | 7.9 | 3.3 | 10 | 3.3 | 3.3 |
| Treatment | Mortality after 24 h (%) 10 µg/mL | Cumulative Mortality a,b,c (%) | Weight Gained (%) | Length Gained (%) | Pupation % | Mean Pupation Time (days) | % IG | Emergence (%) |
|---|---|---|---|---|---|---|---|---|
| Control | 0 | 100 | 100 | 100 | 8 | 100 | 100 | |
| Boldine 1 | 10 | 100 ± 0.0 a,b,c | 7.40 ± 0.07 | 68.3 ± 0.07 | 0.0 | - | - | - |
| Coclaurine 2 | 0 | 100 ± 0.0 a,b,c | 98.5 ± 0.06 | 71.4 ± 0.06 | 0.0 | - | - | - |
| Laurolitsine 3 | 6.7 | 100 ± 0.0 a,b,c | 46.0 ± 0.07 | 66.7 ± 0.07 | 0.0 | - | - | - |
| Pukateine 4 | 0 | 100 ± 0.0 a,b,c | 33.6 ± 0.20 | 66.3 ± 0.08 | 0.0 | - | - | - |
Sample Availability: Samples of the compounds are available from the authors. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quiroz-Carreño, S.; Pastene-Navarrete, E.; Espinoza-Pinochet, C.; Muñoz-Núñez, E.; Devotto-Moreno, L.; Céspedes-Acuña, C.L.; Alarcón-Enos, J. Assessment of Insecticidal Activity of Benzylisoquinoline Alkaloids from Chilean Rhamnaceae Plants against Fruit-Fly Drosophila melanogaster and the Lepidopteran Crop Pest Cydia pomonella. Molecules 2020, 25, 5094. https://doi.org/10.3390/molecules25215094
Quiroz-Carreño S, Pastene-Navarrete E, Espinoza-Pinochet C, Muñoz-Núñez E, Devotto-Moreno L, Céspedes-Acuña CL, Alarcón-Enos J. Assessment of Insecticidal Activity of Benzylisoquinoline Alkaloids from Chilean Rhamnaceae Plants against Fruit-Fly Drosophila melanogaster and the Lepidopteran Crop Pest Cydia pomonella. Molecules. 2020; 25(21):5094. https://doi.org/10.3390/molecules25215094
Chicago/Turabian StyleQuiroz-Carreño, Soledad, Edgar Pastene-Navarrete, Cesar Espinoza-Pinochet, Evelyn Muñoz-Núñez, Luis Devotto-Moreno, Carlos L. Céspedes-Acuña, and Julio Alarcón-Enos. 2020. "Assessment of Insecticidal Activity of Benzylisoquinoline Alkaloids from Chilean Rhamnaceae Plants against Fruit-Fly Drosophila melanogaster and the Lepidopteran Crop Pest Cydia pomonella" Molecules 25, no. 21: 5094. https://doi.org/10.3390/molecules25215094
APA StyleQuiroz-Carreño, S., Pastene-Navarrete, E., Espinoza-Pinochet, C., Muñoz-Núñez, E., Devotto-Moreno, L., Céspedes-Acuña, C. L., & Alarcón-Enos, J. (2020). Assessment of Insecticidal Activity of Benzylisoquinoline Alkaloids from Chilean Rhamnaceae Plants against Fruit-Fly Drosophila melanogaster and the Lepidopteran Crop Pest Cydia pomonella. Molecules, 25(21), 5094. https://doi.org/10.3390/molecules25215094