
Modulation of Lipid Metabolism by Trans-Anethole in Hepatocytes


Abstract
1. Introduction
2. Results
2.1. Effects of TAO on Cytotoxicity and Cellular Metabolism
2.2. Analysis of the Expression Levels of Lipid Metabolism Genes
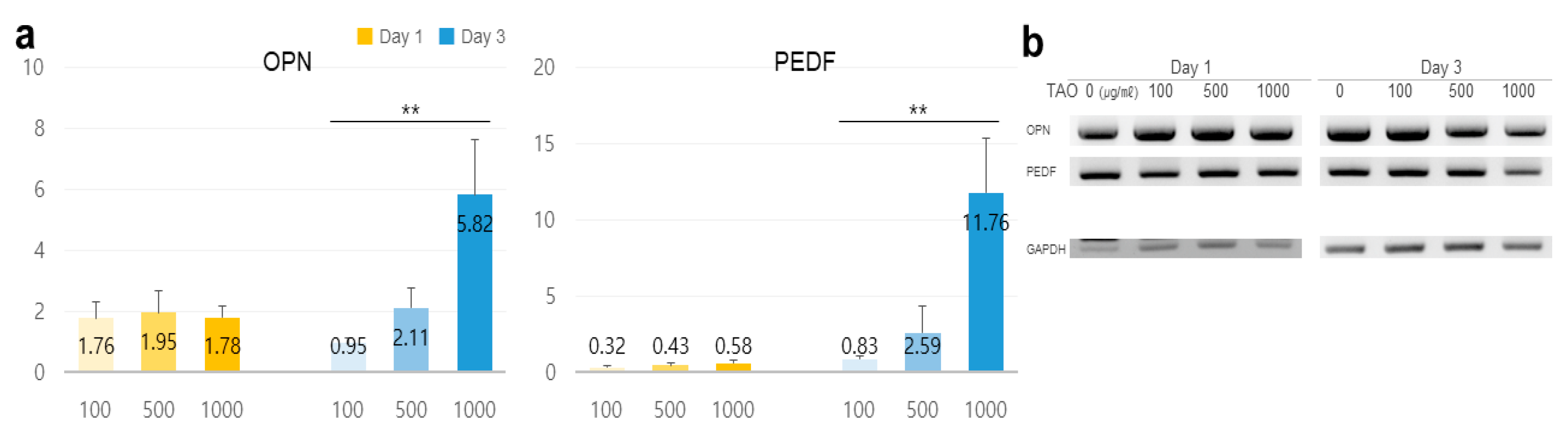
2.3. Potentiality for Reinforcement of the Microenvironment
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Treatment
4.2. Cytotoxicity
4.3. Oil Red O (ORO) Staining
4.4. Conventional Polymerase Chain Reaction (PCR) for Lipid Metabolism Markers
4.5. Flow Cytometry
4.6. Mitochondrial Membrane Potential (MMP) and Cellular Senescence
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Engin, A. Non-Alcoholic Fatty Liver Disease. Obes. Lipotoxicity 2017, 960, 443–467. [Google Scholar] [CrossRef]
- Ipsen, D.H.; Lykkesfeldt, J.; Tveden-Nyborg, P. Molecular mechanisms of hepatic lipid accumulation in non-alcoholic fatty liver disease. Cell. Mol. Life Sci. 2018, 75, 3313–3327. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.-T.; Kuo, P.-L.; Meng-Hsuan, H.; Chen, Y.-Y.; Yeh, M.-L.; Huang, C.-I.; Yang, J.-F.; Lin, C.-I.; Hsieh, M.-H.; Hsieh, M.-Y.; et al. Nonalcoholic fatty liver disease severity is associated with the ratios of total cholesterol and triglycerides to high-density lipoprotein cholesterol. J. Clin. Lipidol. 2016, 10, 420–425.e1. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, K.; Reue, K. Biochemistry, physiology, and genetics of GPAT, AGPAT, and lipin enzymes in triglyceride synthesis. Am. J. Physiol. Metab. 2009, 296, E1195–E1209. [Google Scholar] [CrossRef] [PubMed]
- Yen, C.-L.E.; Stone, S.J.; Koliwad, S.; Harris, C.; Farese, R.V., Jr. Thematic Review Series: Glycerolipids.DGAT enzymes and triacylglycerol biosynthesis. J. Lipid Res. 2008, 49, 2283–2301. [Google Scholar] [CrossRef]
- Tong, L. Acetyl-coenzyme A carboxylase: Crucial metabolic enzyme and attractive target for drug discovery. Cell. Mol. Life Sci. 2005, 62, 1784–1803. [Google Scholar] [CrossRef]
- Kraemer, F.B.; Shen, W.-J. Hormone-sensitive lipase. J. Lipid Res. 2002, 43, 1585–1594. [Google Scholar] [CrossRef]
- Furuhashi, M.; Saitoh, S.; Shimamoto, K.; Miura, T. Fatty Acid-Binding Protein 4 (FABP4): Pathophysiological Insights and Potent Clinical Biomarker of Metabolic and Cardiovascular Diseases. Clin. Med. Insights Cardiol. 2014, 8, CMC.S17067-33. [Google Scholar] [CrossRef]
- Garcia, D.; Shaw, R.J. AMPK: Mechanisms of Cellular Energy Sensing and Restoration of Metabolic Balance. Mol. Cell 2017, 66, 789–800. [Google Scholar] [CrossRef]
- Sztalryd, C.; Xu, G.; Dorward, H.; Tansey, J.T.; Contreras, J.A.; Kimmel, A.R.; Londos, C. Perilipin A is essential for the translocation of hormone-sensitive lipase during lipolytic activation. J. Cell Biol. 2003, 161, 1093–1103. [Google Scholar] [CrossRef]
- Moczulski, D.; Majak, I.; Mamczur, D. An overview of beta-oxidation disorders. Postępy Higieny i Medycyny Doświadczalnej 2009, 63, 266–277. [Google Scholar] [PubMed]
- Nakamura, M.T.; Yudell, B.E.; Loor, J.J. Regulation of energy metabolism by long-chain fatty acids. Prog. Lipid Res. 2014, 53, 124–144. [Google Scholar] [CrossRef] [PubMed]
- Rui, L. Energy Metabolism in the Liver. Compr. Physiol. 2014, 4, 177–197. [Google Scholar] [CrossRef] [PubMed]
- Hillgartner, F.; Charron, T.; Chesnut, K.A. Triiodothyronine Stimulates and Glucagon Inhibits Transcription of the Acetyl-CoA Carboxylase Gene in Chick Embryo Hepatocytes: Glucose and Insulin Amplify the Effect of Triiodothyronine. Arch. Biochem. Biophys. 1997, 337, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Elgendy, E.; Khayyat, S. Oxidation reactions of some natural volatile aromatic compounds: Anethole and eugenol. Russ. J. Org. Chem. 2008, 44, 823–829. [Google Scholar] [CrossRef]
- Bilke, S.; Mosandl, A. 2H/1H and13C/12C Isotope Ratios oftrans-Anethole Using Gas Chromatography−Isotope Ratio Mass Spectrometry. J. Agric. Food Chem. 2002, 50, 3935–3937. [Google Scholar] [CrossRef]
- Muckensturm, B.; Foechterlen, D.; Reduron, J.-P.; Danton, P.; Hildenbrand, M. Phytochemical and chemotaxonomic studies of Foeniculum vulgare. Biochem. Syst. Ecol. 1997, 25, 353–358. [Google Scholar] [CrossRef]
- Shin, Y.J.; Jung, M.J.; Kim, N.S.; Kim, K.; Lee, D.S. Characterization of Fennel Flavors by Solid Phase Trapping-Solvent Extraction and Gas Chromatography-Mass Spectrometry. Bull. Korean Chem. Soc. 2007, 28, 2389–2395. [Google Scholar] [CrossRef]
- Pejin, B.; Bianco, A.; Newmaster, S.; Sabovljevic, M.; Vujisic, L.; Tesevic, V.; Vajs, V.; De Rosa, S. Fatty acids of Rhodobryum ontariense (Bryaceae). Nat. Prod. Res. 2011, 26, 696–702. [Google Scholar] [CrossRef]
- De, M.; De, A.K.; Sen, P.; Banerjee, A.B. Antimicrobial properties of star anise (Illicium verum Hook f). Phytother. Res. 2002, 16, 94–95. [Google Scholar] [CrossRef]
- Fujita, K.-I.; Fujita, T.; Kubo, I. Anethole, a potential antimicrobial synergist, converts a fungistatic dodecanol to a fungicidal agent. Phytother. Res. 2006, 21, 47–51. [Google Scholar] [CrossRef]
- Knio, K.; Usta, J.; Dagher, S.; Zournajian, H.; Kreydiyyeh, S. Larvicidal activity of essential oils extracted from commonly used herbs in Lebanon against the seaside mosquito, Ochlerotatus caspius. Bioresour. Technol. 2008, 99, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, B.A.; Pari, L.; Rathinam, A.; Chandramohan, R. Trans-anethole, a terpenoid ameliorates hyperglycemia by regulating key enzymes of carbohydrate metabolism in streptozotocin induced diabetic rats. Biochimie 2015, 112, 57–65. [Google Scholar] [CrossRef]
- Kang, N.H.; Mukherjee, S.; Min, T.; Kang, S.C.; Yun, J.W. Trans-anethole ameliorates obesity via induction of browning in white adipocytes and activation of brown adipocytes. Biochimie 2018, 151, 1–13. [Google Scholar] [CrossRef]
- Dawidar, A.; Mogib, M.A.; El-Ghorab, A.; Mahfouz, M.; Elsaid, F.; Hussien, K. Chemical Composition and Effect of Photo-Oxygenation on Biological Activities of Egyptian Commercial Anise and Fennel Essential Oils. J. Essent. Oil Bear. Plants 2008, 11, 124–136. [Google Scholar] [CrossRef]
- Gorelick, N. Genotoxicity of trans-anethole in vitro. Mutat. Res. Mol. Mech. Mutagen. 1995, 326, 199–209. [Google Scholar] [CrossRef]
- Europe Co. Active Ingredients Used in Cosmetics: Safety Survey; Committee of Experts on Cosmetic Products; Council of Europe: Strasbourg, France, 2008. [Google Scholar]
- Barbier-Torres, L.; Fortner, K.A.; Iruzubieta, P.; Delgado, T.C.; Giddings, E.; Chen, Y.; Champagne, D.; Fernández-Ramos, D.; Mestre, D.; Gomez-Santos, B.; et al. Silencing hepatic MCJ attenuates non-alcoholic fatty liver disease (NAFLD) by increasing mitochondrial fatty acid oxidation. Nat. Commun. 2020, 11. [Google Scholar] [CrossRef]
- Deng, J.; Liu, S.; Zou, L.; Xu, C.; Geng, B.; Xu, G. Lipolysis Response to Endoplasmic Reticulum Stress in Adipose Cells. J. Biol. Chem. 2012, 287, 6240–6249. [Google Scholar] [CrossRef]
- Winder, W.W.; Hardie, D.G. AMP-activated protein kinase, a metabolic master switch: Possible roles in Type 2 diabetes. Am. J. Physiol. Metab. 1999, 277, E1–E10. [Google Scholar] [CrossRef]
- Lakhani, H.V.; Sharma, D.; Dodrill, M.W.; Nawab, A.; Sharma, N.; Cottrill, C.L.; Shapiro, J.I.; Sodhi, K. Phenotypic Alteration of Hepatocytes in Non-Alcoholic Fatty Liver Disease. Int. J. Med Sci. 2018, 15, 1591–1599. [Google Scholar] [CrossRef] [PubMed]
- Minokoshi, Y.; Kahn, C.R.; Kahn, B.B. Tissue-specific Ablation of the GLUT4 Glucose Transporter or the Insulin Receptor Challenges Assumptions about Insulin Action and Glucose Homeostasis. J. Biol. Chem. 2003, 278, 33609–33612. [Google Scholar] [CrossRef] [PubMed]
- Karim, S.; Liaskou, E.; Fear, J.; Garg, A.; Reynolds, G.; Claridge, L.; Adams, D.H.; Newsome, P.N.; Lalor, P.F. Dysregulated hepatic expression of glucose transporters in chronic disease: Contribution of semicarbazide-sensitive amine oxidase to hepatic glucose uptake. Am. J. Physiol. Liver Physiol. 2014, 307, G1180-90. [Google Scholar] [CrossRef] [PubMed]
- Lund, S.; Holman, G.D.; Schmitz, O.; Pedersen, O. Contraction stimulates translocation of glucose transporter GLUT4 in skeletal muscle through a mechanism distinct from that of insulin. Proc. Natl. Acad. Sci. USA 1995, 92, 5817–5821. [Google Scholar] [CrossRef] [PubMed]
- Shan, W.-F.; Chen, B.-Q.; Zhu, S.-J.; Jiang, L.; Zhou, Y.-F. Effects of GLUT4 expression on insulin resistance in patients with advanced liver cirrhosis. J. Zhejiang Univ. Sci. B 2011, 12, 677–682. [Google Scholar] [CrossRef]
- Nuñez-Garcia, M.; Gomez-Santos, B.; Buqué, X.; García-Rodríguez, J.L.; Romero, M.R.; Marin, J.J.; Arteta, B.; García-Monzón, C.; Castaño, L.; Syn, W.-K.; et al. Osteopontin regulates the cross-talk between phosphatidylcholine and cholesterol metabolism in mouse liver. J. Lipid Res. 2017, 58, 1903–1915. [Google Scholar] [CrossRef]
- Wen, Y.; Feng, D.; Wu, H.; Liu, W.; Li, H.; Wang, F.; Xia, Q.; Gao, W.-Q.; Kong, X. Defective Initiation of Liver Regeneration in Osteopontin-Deficient Mice after Partial Hepatectomy due to Insufficient Activation of IL-6/Stat3 Pathway. Int. J. Biol. Sci. 2015, 11, 1236–1247. [Google Scholar] [CrossRef]
- Rychli, K.; Huber, K.; Wojta, J. Pigment epithelium-derived factor (PEDF) as a therapeutic target in cardiovascular disease. Expert Opin. Ther. Targets 2009, 13, 1295–1302. [Google Scholar] [CrossRef]
- Ma, S.; Wang, S.; Li, M.; Zhang, Y.; Zhu, P. The effects of pigment epithelium-derived factor on atherosclerosis: Putative mechanisms of the process. Lipids Heal. Dis. 2018, 17, 240. [Google Scholar] [CrossRef]
- Song, A.; Kim, B.; Kim, E.; Sung, J.H.; Park, Y.; Park, S.; Park, T.; Kim, J.K.; Jeong, Y.; Lee, S.G. Protective Functions of Group 3 Late Embryogenesis Abundant (G3LEA) Proteins in Enterococcus faecium During Vancomycin Treatment. Indian J. Microbiol. 2020, 60, 1–9. [Google Scholar] [CrossRef]
- Park, Y.; Lee, K.; Lee, C.; Song, A.; Kim, J.; Kim, B.; Lee, S.G. Protection and immune modulation of activated human vaginal epithelial cells by Aurea helianthus extract. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Function | F/R | Seq (5′ → 3′) | Product Size |
|---|---|---|---|---|
| AMPK | Lipid oxidation | F | CGCCTTGATTCTTTTGAGGCTT | 190 |
| R | AGGATCAGACTACACCTGGCT | |||
| GLUT4 | Glucose uptake | F | TCTCCAACTGGACGAGCAAC | 269 |
| R | AGTTATGCCACTGGTGCGTT | |||
| ACS | Lipid oxidation | F | CCTGGGATCTCTCTCATGGC | 289 |
| R | CCCCAACAACTTGCAGTGAT | |||
| CPT2 | Lipid oxidation | F | GACTCGGCAGTGTTCTGTCT | 674 |
| R | GTCAGCTGGCCATGGTACTTG | |||
| ACADS | Lipid oxidation | F | TTCATCAAGGAGCCGGCAAT | 306 |
| R | AGGGTAAAGGCACATGGCTC | |||
| HSL | Triglyceride lipolysis enzyme | F | AGCTGAGACACTTAGCCCCT | 383 |
| R | CACTCCGGAGCTCTTTTTCC | |||
| AP2 | Lipolysis | F | TGGTGGTGGTGAGTATCTTCT | 574 |
| R | GGTCAACGTCCCTTGGCTTA | |||
| LPL | Triglyceride lipolysis enzyme | F | GGCAGCTTCATGCATTCCTC | 326 |
| R | CAGCCAGAACGGCAACTACT | |||
| ACC1 | Lipid synthesis | F | GCACATCTTCACACTCCTGAA | 110 |
| R | GTACCACTCACCTGCCGTAT | |||
| GPAT | Triglyceride synthesis | F | TGGGTGAAGAATTCTGGTGGA | 292 |
| R | CATGAGGGGTGCAGGTGTAG | |||
| OPN | Changing micro-environment | F | GAATCTCCTAGCCCCACAGACC | 379 |
| R | GTGTGAGGTGATGTCCTCGTC | |||
| PEDF | Changing micro-environment | F | GCTGAGTTACGAAGGCGAAGT | 102 |
| R | GCTGAGTTACGAAGGCGAAGT | |||
| GAPDH | Housekeeping | F | GTGGTCTCCTCTGACTTCAACA | 210 |
| R | CTCTTCCTCTTGTGCTCTTGCT |
Sample Availability: Samples of the compound is not available from the authors. | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, A.; Park, Y.; Kim, B.; Lee, S.G. Modulation of Lipid Metabolism by Trans-Anethole in Hepatocytes. Molecules 2020, 25, 4946. https://doi.org/10.3390/molecules25214946
Song A, Park Y, Kim B, Lee SG. Modulation of Lipid Metabolism by Trans-Anethole in Hepatocytes. Molecules. 2020; 25(21):4946. https://doi.org/10.3390/molecules25214946
Chicago/Turabian StyleSong, Ahran, Yoonjin Park, Boyong Kim, and Seung Gwan Lee. 2020. "Modulation of Lipid Metabolism by Trans-Anethole in Hepatocytes" Molecules 25, no. 21: 4946. https://doi.org/10.3390/molecules25214946
APA StyleSong, A., Park, Y., Kim, B., & Lee, S. G. (2020). Modulation of Lipid Metabolism by Trans-Anethole in Hepatocytes. Molecules, 25(21), 4946. https://doi.org/10.3390/molecules25214946