
Isolation and Characterization of Phenylpropanoid and Lignan Compounds from Peperomia pellucida [L.] Kunth with Estrogenic Activities

 , ,
, ,
Abstract
1. Introduction
2. Results
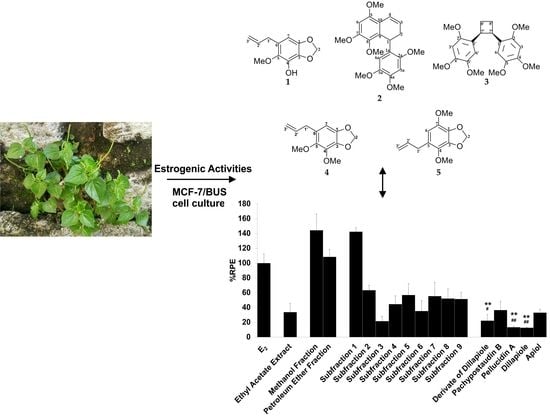
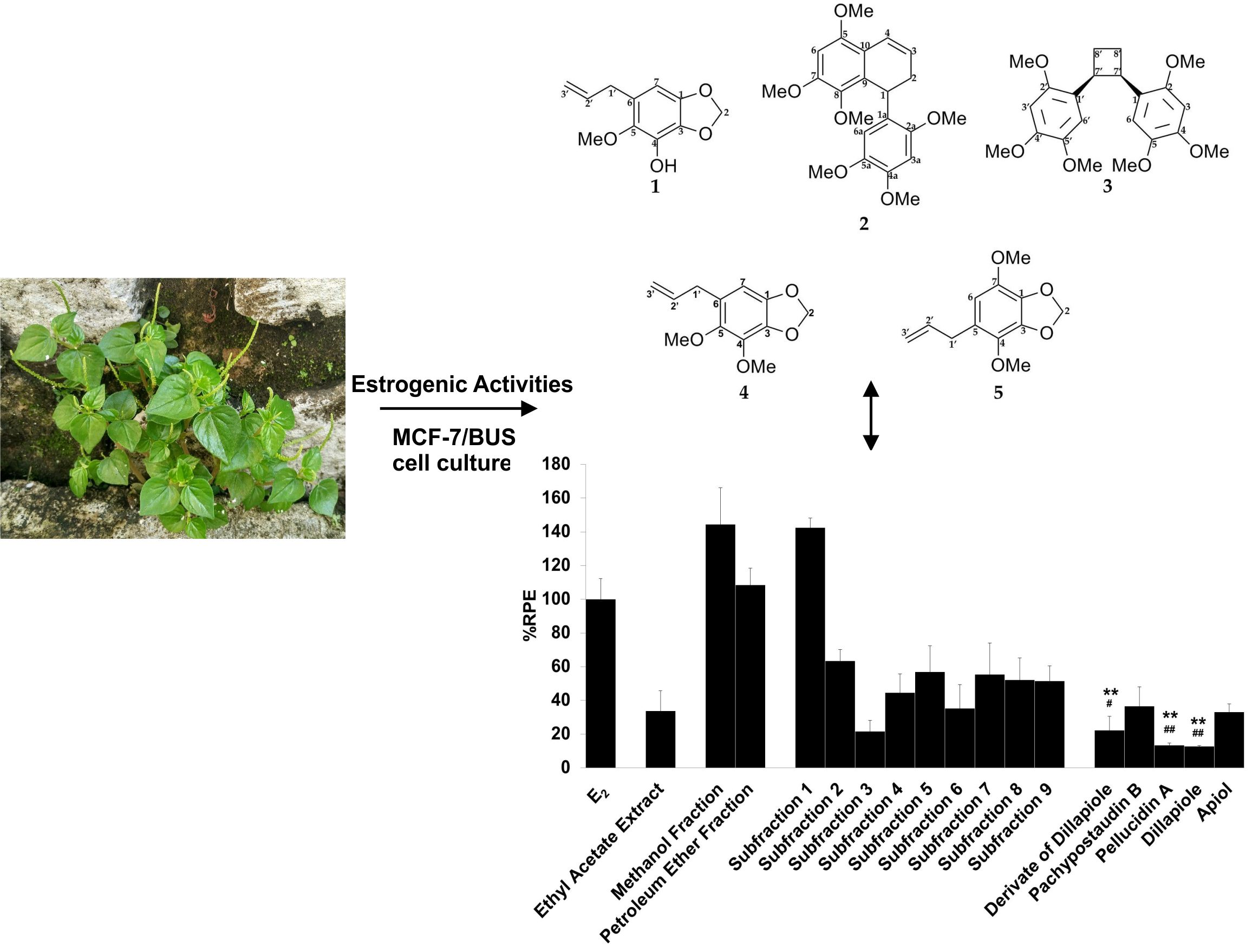
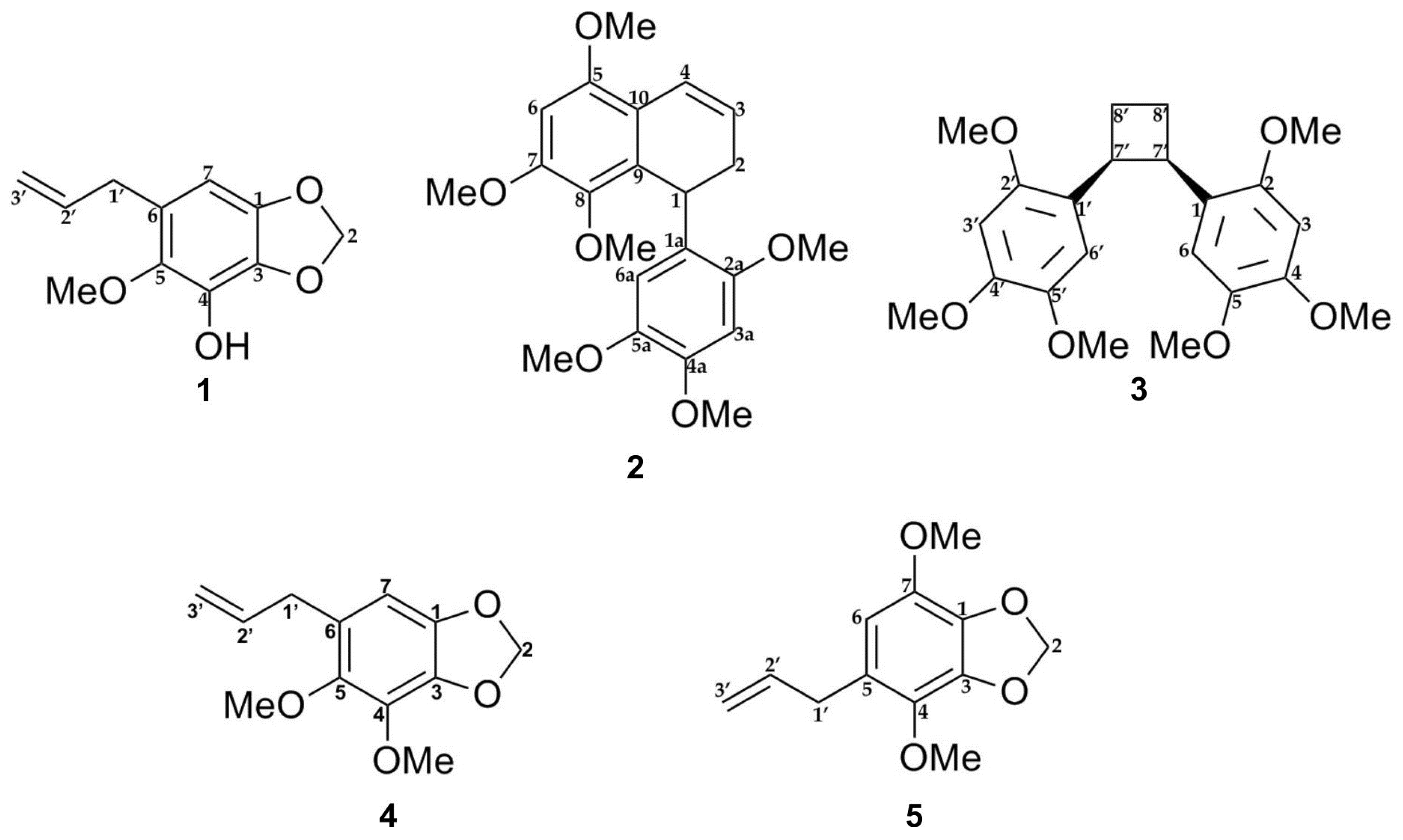
2.1. Isolation and Identification of Compounds from P. pellucida Ethyl Acetate Extract
2.2. Cytotoxicity Assay
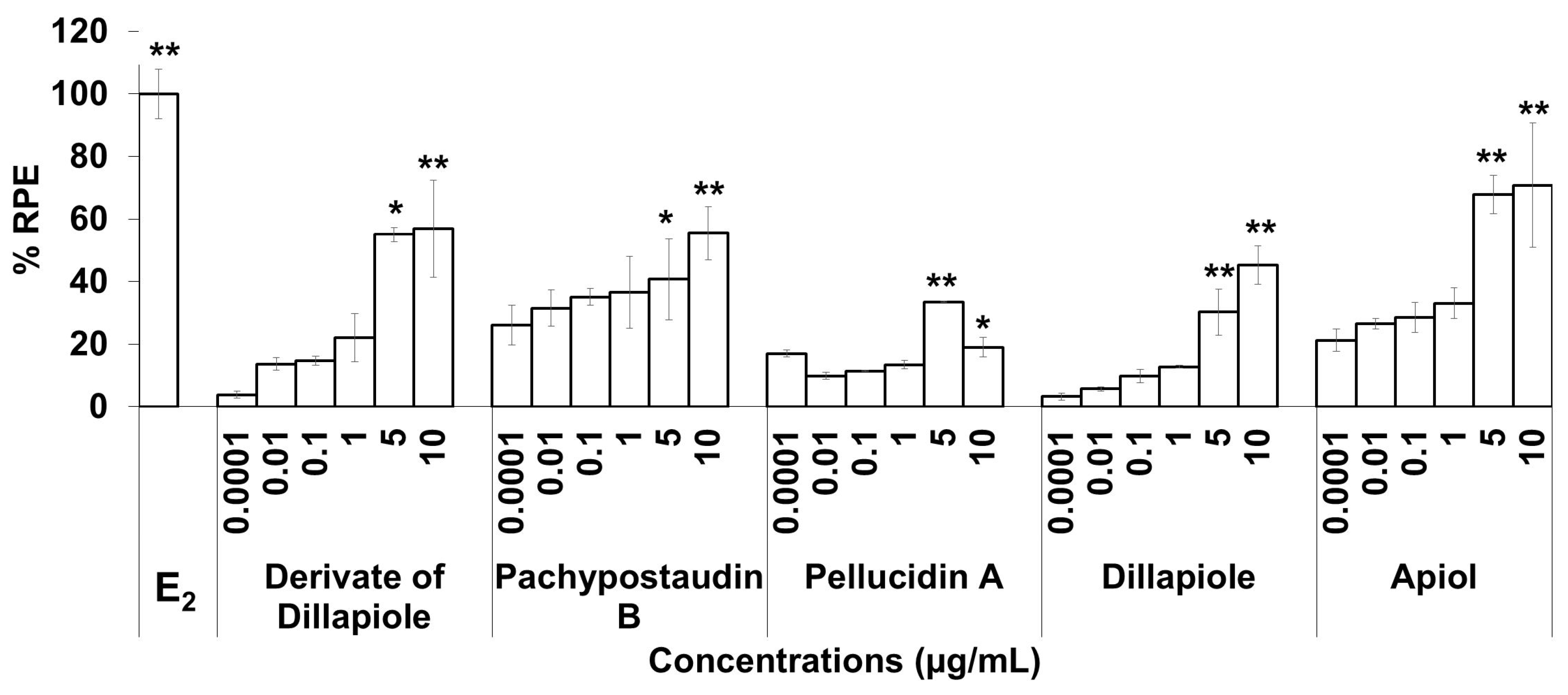
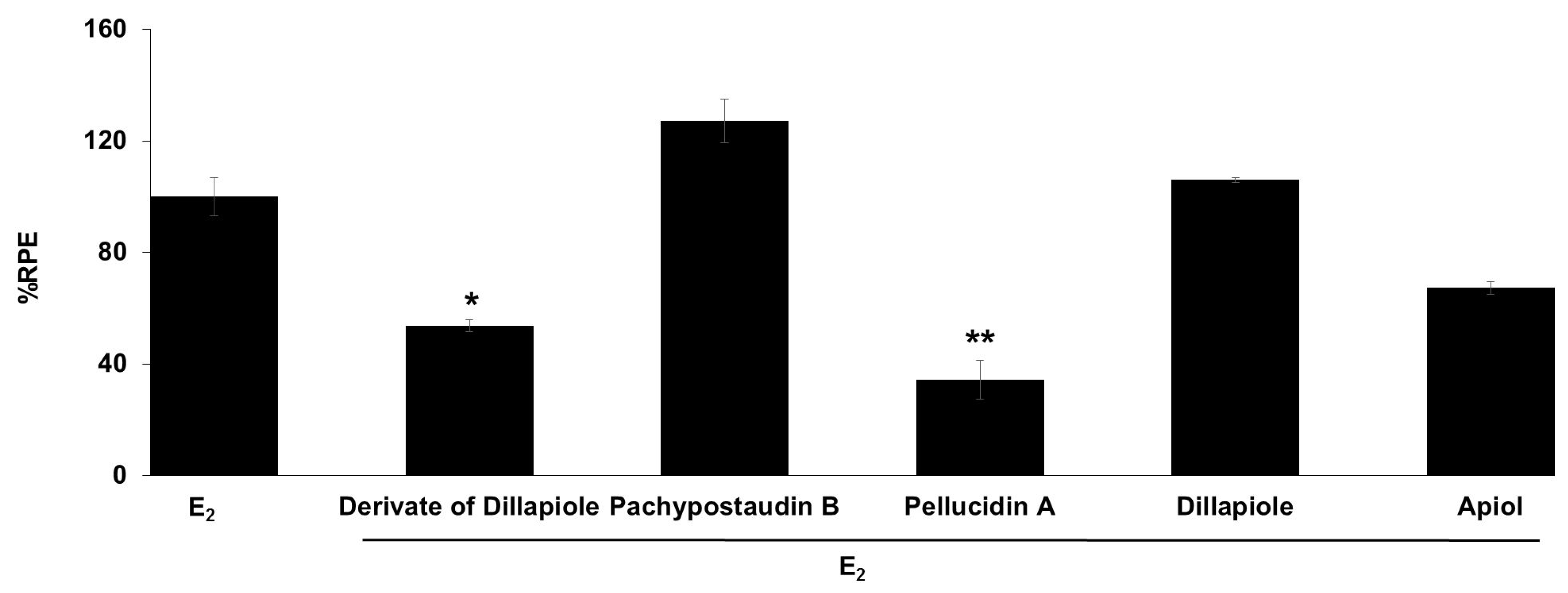
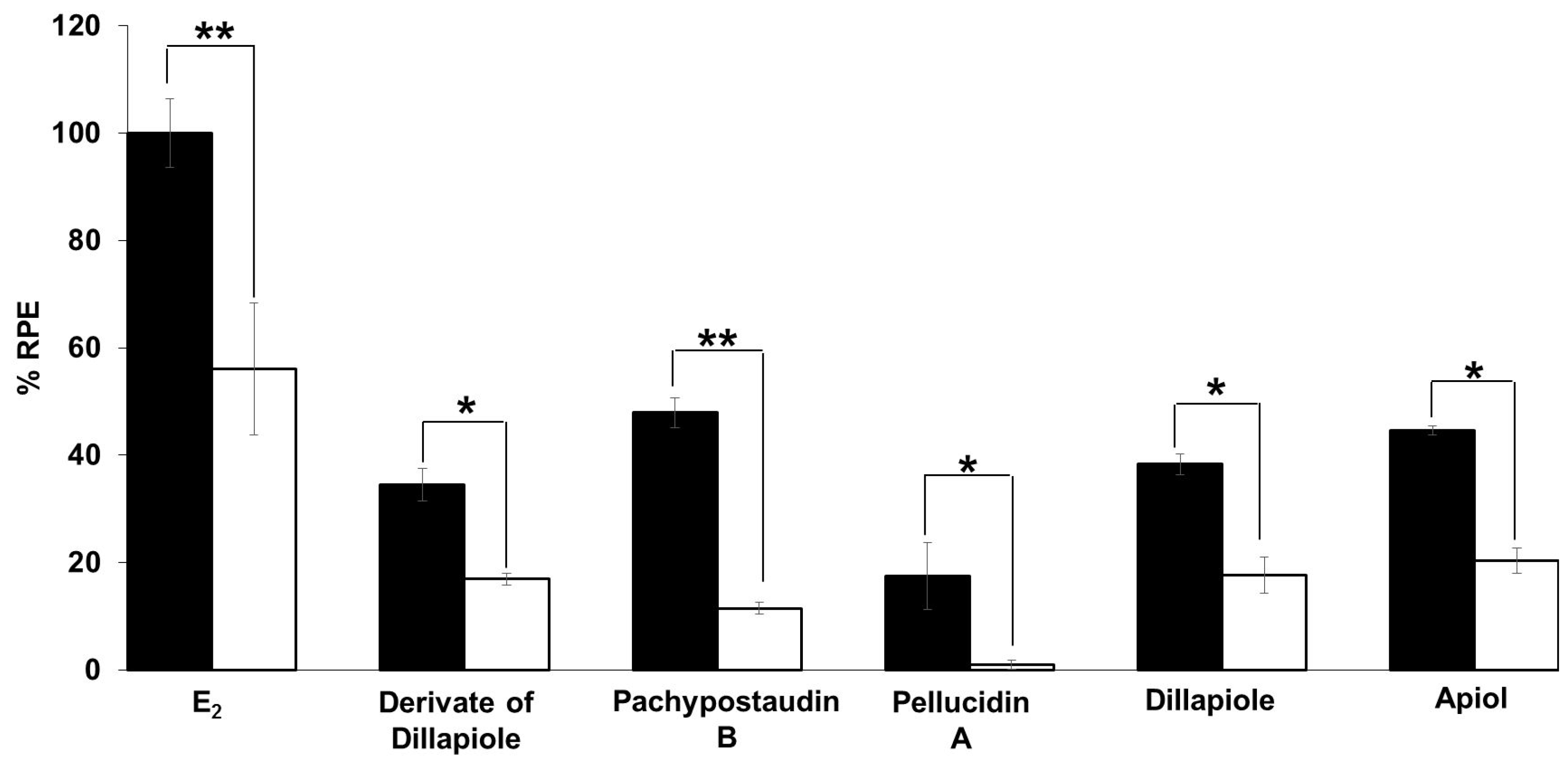
2.3. E-SCREEN Assay
2.4. Molecular Docking
3. Discussion
4. Materials and Methods
4.1. Instruments and Materials
4.2. Extraction Process
4.3. Isolation Process
4.4. Cell Lines
4.5. Cytotoxicity Assay
4.6. E-SCREEN Assay
4.7. Molecular Docking Test
4.8. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Couse, J.F.; Korach, K.S. Estrogen receptor null mice: What have we learned and where will they lead us? Endocr. Rev. 1999, 20, 358–417. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, J.-A. What pharmacologists can learn from recent advances in estrogen signalling. Trends Pharmacol. Sci. 2003, 24, 479–485. [Google Scholar] [CrossRef]
- Nelson, A.W.; Tilley, W.D.; Neal, D.E.; Carroll, J.S. Estrogen receptor beta in prostate cancer: Friend or foe? Endocr. Relat. Cancer 2014, 21, T219–T234. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.; Edvardsson, K.; Lewandowski, S.A.; Ström, A.; Gustafsson, J.-A. A genome-wide study of the repressive effects of estrogen receptor beta on estrogen receptor alpha signaling in breast cancer cells. Oncogene 2008, 27, 1019–1032. [Google Scholar] [CrossRef] [PubMed]
- Mangalath, D.L.; Sadasivan, C. Selective Estrogen Receptor Modulators (SERMs) from Plants. In Bioactive Natural Products; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2015; pp. 375–386. ISBN 978-3-527-68440-3. [Google Scholar]
- Sirtori, C.R.; Arnoldi, A.; Johnson, S.K. Phytoestrogens: End of a tale? Ann. Med. 2005, 37, 423–438. [Google Scholar] [CrossRef] [PubMed]
- Oseni, T.; Patel, R.; Pyle, J.; Jordan, V.C. Selective estrogen receptor modulators and phytoestrogens. Planta Med. 2008, 74, 1656–1665. [Google Scholar] [CrossRef] [PubMed]
- Lampe, J.W. Isoflavonoid and Lignan Phytoestrogens as Dietary Biomarkers. J. Nutr. 2003, 133, 956S–964S. [Google Scholar] [CrossRef]
- Cornwell, T.; Cohick, W.; Raskin, I. Dietary phytoestrogens and health. Phytochemistry 2004, 65, 995–1016. [Google Scholar] [CrossRef]
- Sirotkin, A.V.; Harrath, A.H. Phytoestrogens and their effects. Eur. J. Pharmacol. 2014, 741, 230–236. [Google Scholar] [CrossRef]
- Poluzzi, E.; Piccinni, C.; Raschi, E.; Rampa, A.; Recanatini, M.; De Ponti, F. Phytoestrogens in postmenopause: The state of the art from a chemical, pharmacological and regulatory perspective. Curr. Med. Chem. 2014, 21, 417–436. [Google Scholar] [CrossRef]
- Florence, N.T.; Huguette, S.T.S.; Hubert, D.J.; Raceline, G.K.; Desire, D.D.P.; Pierre, K.; Theophile, D. Aqueous extract of Peperomia pellucida (L.) HBK accelerates fracture healing in Wistar rats. BMC Complement. Altern. Med. 2017, 17, 188. [Google Scholar] [CrossRef] [PubMed]
- Ngueguim, F.T.; Khan, M.P.; Donfack, J.H.; Tewari, D.; Dimo, T.; Kamtchouing, P.; Maurya, R.; Chattopadhyay, N. Ethanol extract of Peperomia pellucida (Piperaceae) promotes fracture healing by an anabolic effect on osteoblasts. J. Ethnopharmacol. 2013, 148, 62–68. [Google Scholar] [CrossRef]
- Putri, C.A.; Kartika, A.; Adnyana, I.K. Preventive effect of Peperomia pellucida (L.) Kunth herbs on ovariectomy-induced osteoporotic rats. Chin. Med. 2016, 25, 546–551. [Google Scholar] [CrossRef]
- Khosla, S.; Oursler, M.J.; Monroe, D.G. Estrogen and the Skeleton. Trends Endocrinol. Metab. 2012, 23, 576–581. [Google Scholar] [CrossRef]
- Deroo, B.J.; Korach, K.S. Estrogen receptors and human disease. J. Clin. Investig. 2006, 116, 561–570. [Google Scholar] [CrossRef]
- Villalobos, M.; Olea, N.; Brotons, J.A.; Olea-Serrano, M.F.; Ruiz de Almodovar, J.M.; Pedraza, V. The E-screen assay: A comparison of different MCF7 cell stocks. Environ. Health Perspect. 1995, 103, 844–850. [Google Scholar] [CrossRef]
- Liu, Z.; Yu, X.; Shaikh, Z.A. Rapid activation of ERK1/2 and AKT in human breast cancer cells by cadmium. Toxicol. Appl. Pharmacol. 2007, 228, 286–294. [Google Scholar] [CrossRef]
- Desouza, J.; Gadkar, S.; Jagtap, D.; Sachdeva, G. Size, site, and signaling: Three attributes of estrogen receptors. Biomed. Res. J. 2019, 6, 37. [Google Scholar] [CrossRef]
- Levin, E.R. Plasma membrane estrogen receptors. Trends Endocrinol. Metab. 2009, 20, 477–482. [Google Scholar] [CrossRef]
- Pedram, A.; Razandi, M.; Levin, E.R. Nature of functional estrogen receptors at the plasma membrane. Mol. Endocrinol. 2006, 20, 1996–2009. [Google Scholar] [CrossRef] [PubMed]
- Siersbæk, R.; Kumar, S.; Carroll, J.S. Signaling pathways and steroid receptors modulating estrogen receptor α function in breast cancer. Genes Dev. 2018, 32, 1141–1154. [Google Scholar] [CrossRef]
- Fuentes, N.; Silveyra, P. Estrogen receptor signaling mechanisms. Adv. Protein Chem. Struct. Biol. 2019, 116, 135–170. [Google Scholar] [CrossRef]
- Marczell, I.; Balogh, P.; Nyiro, G.; Kiss, A.L.; Kovacs, B.; Bekesi, G.; Racz, K.; Patocs, A. Membrane-bound estrogen receptor alpha initiated signaling is dynamin dependent in breast cancer cells. Eur. J. Med. Res. 2018, 23, 31. [Google Scholar] [CrossRef] [PubMed]
- Björnström, L.; Sjöberg, M. Mechanisms of Estrogen Receptor Signaling: Convergence of Genomic and Nongenomic Actions on Target Genes. Mol. Endocrinol. 2005, 19, 833–842. [Google Scholar] [CrossRef]
- Ward, R.S. Recent Advances in the Chemistry of Lignans. In Studies in Natural Products Chemistry; Bioactive Natural Products (Part E); Atta-ur-Rahman, Ed.; Elsevier: Amsterdam, The Netherlands, 2000; Volume 24, pp. 739–798. [Google Scholar]
- Simpson, D.; Amos, S. Chapter 12—Other Plant Metabolites. In Pharmacognosy; Badal, S., Delgoda, R., Eds.; Academic Press: Boston, MA, USA, 2017; pp. 267–280. ISBN 978-0-12-802104-0. [Google Scholar]
- Cui, Q.; Du, R.; Liu, M.; Rong, L. Lignans and Their Derivatives from Plants as Antivirals. Molecules 2020, 25, 183. [Google Scholar] [CrossRef] [PubMed]
- Ayres, D.C.; Loike, J.D. Lignans: Chemical, Biological and Clinical Properties; Cambridge University Press: Cambridge, UK, 1990; ISBN 978-0-521-30421-4. [Google Scholar]
- Ward, R.S. Lignans, neolignans, and related compounds. Nat. Prod. Rep. 1993, 10, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Umezawa, T. Biosynthesis of lignans and norlignans. J. Wood Sci. 2007, 53, 273–284. [Google Scholar] [CrossRef]
- Frezza, C.; Venditti, A.; Toniolo, C.; De Vita, D.; Franceschin, M.; Ventrone, A.; Tomassini, L.; Foddai, S.; Guiso, M.; Nicoletti, M.; et al. Nor-Lignans: Occurrence in Plants and Biological Activities—A Review. Molecules 2020, 25, 197. [Google Scholar] [CrossRef]
- Bayma, J.D.; Arruda, M.S.; Müller, A.H.; Arruda, A.C.; Canto, W.C. A dimeric ArC2 compound from Peperomia pellucida. Phytochemistry 2000, 55, 779–782. [Google Scholar] [CrossRef]
- Ragasa, C.; Dumato, M.; Rideout, J. Antifungal compounds from Peperomia pellucida. ACGC Chem. Res. Commun. 1998, 7, 54–61. [Google Scholar]
- Ngadjui, B.T.; Lontsi, D.; Ayafor, J.F.; Sondengam, B.L. Pachypophyllin and pachypostaudins A and B: Three bisnorlignans from pachypodanthium staudtii. Phytochemistry 1989, 28, 231–234. [Google Scholar] [CrossRef]
- Eklund, P.; Raitanen, J.-E. 9-Norlignans: Occurrence, Properties and Their Semisynthetic Preparation from Hydroxymatairesinol. Molecules 2019, 24, 220. [Google Scholar] [CrossRef] [PubMed]
- Kartika, I.G.A.A.; Insanu, M.; Riani, C.; Chung, K.H.; Adnyana, I.K. Polarity Difference and The Presence of Phytoestrogen Compounds Affecting Estrogenic Activity of Peperomia pellucida Extracts. Sains Malays, accepted.
- Soto, A.; Sonnenschein, C.L.; Chung, K.; Fernández, M.; Olea, N.; Olea Serrano, F. The E-SCREEN Assay as a Tool to Identify Estrogens: An Update on Estrogenic Environmental Pollutants. Environ. Health Perspect. 1995, 103 (Suppl. 7), 113–122. [Google Scholar] [CrossRef] [PubMed]
- Resende, F.A.; de Oliveira, A.P.S.; de Camargo, M.S.; Vilegas, W.; Varanda, E.A. Evaluation of Estrogenic Potential of Flavonoids Using a Recombinant Yeast Strain and MCF7/BUS Cell Proliferation Assay. PLoS ONE 2013, 8, e74881. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Yanuar, A.; Mulia, K.; Mun’im, A. Extraction of Polyphenolic Content from Peperomia pellucida (L) Kunth Herb with 1-Ethyl-3-methylimidazolium Bromide. Indian J. Pharm. Sci. 2018, 79, 1013–1017. [Google Scholar] [CrossRef]
- Bacciottini, L.; Falchetti, A.; Pampaloni, B.; Bartolini, E.; Carossino, A.M.; Brandi, M.L. Phytoestrogens: Food or drug? Clin. Cases Miner. Bone Metab. 2007, 4, 123–130. [Google Scholar]
- An, K.-C. Selective Estrogen Receptor Modulators. Asian Spine J. 2016, 10, 787–791. [Google Scholar] [CrossRef]
- Hajirahimkhan, A.; Simmler, C.; Yuan, Y.; Anderson, J.R.; Chen, S.-N.; Nikolić, D.; Dietz, B.M.; Pauli, G.F.; van Breemen, R.B.; Bolton, J.L. Evaluation of Estrogenic Activity of Licorice Species in Comparison with Hops Used in Botanicals for Menopausal Symptoms. PLoS ONE 2013, 8, e67947. [Google Scholar] [CrossRef]
- MacGregor, J.I.; Jordan, V.C. Basic guide to the mechanisms of antiestrogen action. Pharmacol. Rev. 1998, 50, 151–196. [Google Scholar]
- Mäkelä, S.; Santti, R.; Salo, L.; McLachlan, J.A. Phytoestrogens are partial estrogen agonists in the adult male mouse. Environ. Health Perspect. 1995, 103, 123–127. [Google Scholar]
- Lecomte, S.; Demay, F.; Ferrière, F.; Pakdel, F. Phytochemicals Targeting Estrogen Receptors: Beneficial Rather Than Adverse Effects? Int. J. Mol. Sci. 2017, 18, 1381. [Google Scholar] [CrossRef]
- Nath, D. Phytoestrogens: A Possible Agent for Controlling Fertility. GJORM 2018, 3. [Google Scholar] [CrossRef]
- Mostrom, M.; Evans, T.J. Chapter 52—Phytoestrogens. In Reproductive and Developmental Toxicology; Gupta, R.C., Ed.; Academic Press: San Diego, CA, USA, 2011; pp. 707–722. ISBN 978-0-12-382032-7. [Google Scholar]
- Mathouet, H.; Elomri, A.; Lameiras, P.; Daïch, A.; Vérité, P. An alkaloid, two conjugate sesquiterpenes and a phenylpropanoid from Pachypodanthium confine Engl. and Diels. Phytochemistry 2007, 68, 1813–1818. [Google Scholar] [CrossRef]
- Manalo, J.B.; Han, B.H.; Han, Y.H.; Park, M.H.; Anzaldo, F.E. Studies on ether-soluble neutral compounds of Peperomia pellucida. Arch. Pharm. Res. 1983, 6, 133–136. [Google Scholar] [CrossRef]
- Budzinski, J.W.; Trudeau, V.L.; Drouin, C.E.; Panahi, M.; Arnason, J.T.; Foster, B.C. Modulation of human cytochrome P450 3A4 (CYP3A4) and P-glycoprotein (P-gp) in Caco-2 cell monolayers by selected commercial-source milk thistle and goldenseal products. Can. J. Physiol. Pharmacol. 2007, 85, 966–978. [Google Scholar] [CrossRef]
- Kwon, Y.S.; Choi, W.G.; Kim, W.J.; Kim, W.K.; Kim, M.J.; Kang, W.H.; Kim, C.M. Antimicrobial constituents of Foeniculum vulgare. Arch. Pharm. Res. 2002, 25, 154–157. [Google Scholar] [CrossRef]
- Monteiro, M. Antibacterial activity of the Piper aduncum oil and dillapiole, Its main constituent, Against multidrug-resistant strains. Bol. Latinoam. Caribe Plantas Med. Aromat. 2014, 13, 517–526. [Google Scholar]
- Parise-Filho, R.; Pastrello, M.; Pereira Camerlingo, C.E.; Silva, G.J.; Agostinho, L.A.; de Souza, T.; Motter Magri, F.M.; Ribeiro, R.R.; Brandt, C.A.; Polli, M.C. The anti-inflammatory activity of dillapiole and some semisynthetic analogues. Pharm. Biol. 2011, 49, 1173–1179. [Google Scholar] [CrossRef] [PubMed]
- Rafael, M.S.; Hereira-Rojas, W.J.; Roper, J.J.; Nunomura, S.M.; Tadei, W.P. Potential control of Aedes aegypti (Diptera: Culicidae) with Piper aduncum L. (Piperaceae) extracts demonstrated by chromosomal biomarkers and toxic effects on interphase nuclei. Genet. Mol. Res. 2008, 7, 772–781. [Google Scholar] [CrossRef]
- Vizcaíno Páez, S.; Pineda Mejía, R.; Garcia, C.; Gil, J.; Durango, D. Metabolism and antifungal activity of safrole, dillapiole, and derivatives against Botryodiplodia theobromae and Colletotrichum acutatum. Bol. Latinoam. Caribe Plantas Med. Aromat. 2016, 15, 1–17. [Google Scholar]
- Zhou, G.-D.; Moorthy, B.; Bi, J.; Donnelly, K.C.; Randerath, K. DNA adducts from alkoxyallylbenzene herb and spice constituents in cultured human (HepG2) cells. Environ. Mol. Mutagen. 2007, 48, 715–721. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Martínez, R.; Arrieta, J.; Cruz-Antonio, L.; Arrieta-Baez, D.; Velázquez-Méndez, A.; Sánchez-Mendoza, M.; Rojas-Martínez, R.; Arrieta, J.; Cruz-Antonio, L.; Arrieta-Baez, D.; et al. Dillapiole, Isolated from Peperomia pellucida, Shows Gastroprotector Activity against Ethanol-Induced Gastric Lesions in Wistar Rats. Molecules 2013, 18, 11327–11337. [Google Scholar] [CrossRef] [PubMed]
- Song, H.Y.; Yang, J.Y.; Suh, J.W.; Lee, H.S. Acaricidal Activities of Apiol and Its Derivatives from Petroselinum sativum Seeds against Dermatophagoides pteronyssinus, Dermatophagoides farinae, and Tyrophagus putrescentiae. J. Agric. Food Chem. 2011, 59, 7759–7764. [Google Scholar] [CrossRef] [PubMed]
- Stefano, V.D.; Pitonzo, R.; Schillaci, D. Antimicrobial and antiproliferative activity of Athamanta sicula L. (Apiaceae). Pharmacogn. Mag. 2011, 7, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Creusot, N.; Budzinski, H.; Balaguer, P.; Kinani, S.; Porcher, J.-M.; Aït-Aïssa, S. Effect-directed analysis of endocrine-disrupting compounds in multi-contaminated sediment: Identification of novel ligands of estrogen and pregnane X receptors. Anal. Bioanal. Chem. 2013, 405, 2553–2566. [Google Scholar] [CrossRef]
- Katzenellenbogen, B.S.; Miller, M.A.; Eckert, R.L.; Sudo, K. Antiestrogen pharmacology and mechanism of action. J. Steroid Biochem. 1983, 19, 59–68. [Google Scholar] [CrossRef]
- Mueller, S.O.; Simon, S.; Chae, K.; Metzler, M.; Korach, K.S. Phytoestrogens and their human metabolites show distinct agonistic and antagonistic properties on estrogen receptor alpha (ERalpha) and ERbeta in human cells. Toxicol. Sci. 2004, 80, 14–25. [Google Scholar] [CrossRef]
- Xu, S.; Li, N.; Ning, M.-M.; Zhou, C.-H.; Yang, Q.-R.; Wang, M.-W. Bioactive Compounds from Peperomia pellucida. J. Nat. Prod. 2006, 69, 247–250. [Google Scholar] [CrossRef]
- Chang, E.C.; Frasor, J.; Komm, B.; Katzenellenbogen, B.S. Impact of estrogen receptor beta on gene networks regulated by estrogen receptor alpha in breast cancer cells. Endocrinology 2006, 147, 4831–4842. [Google Scholar] [CrossRef]
- Mak, P.; Leav, I.; Pursell, B.; Bae, D.; Yang, X.; Taglienti, C.A.; Gouvin, L.M.; Sharma, V.M.; Mercurio, A.M. ERbeta impedes prostate cancer EMT by destabilizing HIF-1alpha and inhibiting VEGF-mediated snail nuclear localization: Implications for Gleason grading. Cancer Cell 2010, 17, 319–332. [Google Scholar] [CrossRef] [PubMed]
- McPherson, S.J.; Hussain, S.; Balanathan, P.; Hedwards, S.L.; Niranjan, B.; Grant, M.; Chandrasiri, U.P.; Toivanen, R.; Wang, Y.; Taylor, R.A.; et al. Estrogen receptor-beta activated apoptosis in benign hyperplasia and cancer of the prostate is androgen independent and TNFalpha mediated. Proc. Natl. Acad. Sci. USA 2010, 107, 3123–3128. [Google Scholar] [CrossRef] [PubMed]
- Paterni, I.; Granchi, C.; Katzenellenbogen, J.A.; Minutolo, F. Estrogen receptors alpha (ERα) and beta (ERβ): Subtype-selective ligands and clinical potential. Steroids 2014, 90, 13–29. [Google Scholar] [CrossRef]
- Pinton, G.; Thomas, W.; Bellini, P.; Manente, A.G.; Favoni, R.E.; Harvey, B.J.; Mutti, L.; Moro, L. Estrogen receptor β exerts tumor repressive functions in human malignant pleural mesothelioma via EGFR inactivation and affects response to gefitinib. PLoS ONE 2010, 5, e14110. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, A.; Toth, C.; Hoffmeister, M.; Roth, W.; Herpel, E.; Jansen, L.; Marx, A.; Brenner, H.; Chang-Claude, J. Expression of oestrogen receptor β and prognosis of colorectal cancer. Br. J. Cancer 2012, 107, 831–839. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.-P.; Ho, J.-Y.; Huang, Y.-T.; Cha, T.-L.; Sun, G.-H.; Yu, D.-S.; Chang, F.-W.; Chen, S.-P.; Hsu, R.-J. Estrogen inhibits renal cell carcinoma cell progression through estrogen receptor-β activation. PLoS ONE 2013, 8, e56667. [Google Scholar] [CrossRef]
- Garcia, A.J.; Tom, C.; Guemes, M.; Polanco, G.; Mayorga, M.E.; Wend, K.; Miranda-Carboni, G.A.; Krum, S.A. ERα signaling regulates MMP3 expression to induce FasL cleavage and osteoclast apoptosis. J. Bone Miner. Res. 2013, 28, 283–290. [Google Scholar] [CrossRef]
- Khalid, A.B.; Krum, S.A. Estrogen Receptors Alpha and Beta in Bone. Bone 2016, 87, 130–135. [Google Scholar] [CrossRef]
- Berry, M.; Metzger, D.; Chambon, P. Role of the two activating domains of the oestrogen receptor in the cell-type and promoter-context dependent agonistic activity of the anti-oestrogen 4-hydroxytamoxifen. EMBO J. 1990, 9, 2811–2818. [Google Scholar] [CrossRef]
- Peng, J.; Sengupta, S.; Jordan, V.C. Potential of selective estrogen receptor modulators as treatments and preventives of breast cancer. Anticancer Agents Med. Chem. 2009, 9, 481–499. [Google Scholar] [CrossRef]
- Shang, Y.; Brown, M. Molecular determinants for the tissue specificity of SERMs. Science 2002, 295, 2465–2468. [Google Scholar] [CrossRef]
- Raffo, D.; Pontiggia, O.; Bal de Kier Joffé, E.; Simian, M. Non-genomic actions of estradiol and 4-OH-tamoxifen on murine breast cancer cells. Oncol. Rep. 2015, 33, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Bender, W. The mechanism of tamoxifen in breast cancer prevention. Breast Cancer Res. 2001, 3, A74. [Google Scholar] [CrossRef]
- Khan, F.; Peter, X.K.; Mackenzie, R.M.; Katsoulis, L.; Gehring, R.; Munro, O.Q.; van Heerden, F.R.; Drewes, S.E. Venusol from Gunnera perpensa: Structural and activity studies. Phytochemistry 2004, 65, 1117–1121. [Google Scholar] [CrossRef] [PubMed]
- De Mariotti, K.C.; Schmitt, G.C.; Barreto, F.; Fortunato, R.E.; Singer, R.B.; Dallegrave, E.; Leal, M.B.; Limberger, R.P. Evaluation of anti-estrogenic or estrogenic activities of aqueous root extracts of Gunnera manicata L. Braz. J. Pharm. Sci. 2011, 47, 601–604. [Google Scholar] [CrossRef]
- Dauvois, S.; White, R.; Parker, M.G. The antiestrogen ICI 182780 disrupts estrogen receptor nucleocytoplasmic shuttling. J. Cell Sci. 1993, 106, 1377–1388. [Google Scholar] [PubMed]
- Kim, H.R.; Lee, J.E.; Jeong, M.H.; Choi, S.J.; Lee, K.; Chung, K.H. Comparative evaluation of the mutagenicity and genotoxicity of smoke condensate derived from Korean cigarettes. Environ. Health Toxicol. 2015, 30. [Google Scholar] [CrossRef]
- Vandhana, S.; Deepa, P.R.; Aparna, G.; Jayanthi, U.; Krishnakumar, S. Evaluation of suitable solvents for testing the anti-proliferative activity of triclosan—A hydrophobic drug in cell culture. IJBB 2010, 47, 166–171. [Google Scholar]
- Jamalzadeh, L.; Ghafoori, H.; Sariri, R.; Rabuti, H.; Nasirzade, J.; Hasani, H.; Aghamaali, M.R. Cytotoxic Effects of Some Common Organic Solvents on MCF-7, RAW-264.7 and Human Umbilical Vein Endothelial Cells. Avicenna J. Med. Biochem. 2016, in press. [Google Scholar] [CrossRef]
- Oh, S.M.; Ryu, B.T.; Chung, K.H. Identification of estrogenic and antiestrogenic activities of respirable diesel exhaust particles by bioassay-directed fractionation. Arch. Pharm. Res. 2008, 31, 75–82. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carbon Position | DEPT | HSQC | COSY | NOESY | HMBC | |
|---|---|---|---|---|---|---|
| C/H | 13C | 1H | ||||
| 1 | CH | 28.5 | 4.89 (1H, d, 8.11 Hz) | H-2 | C-2, 3, 8, 9, 10, 1a, 2a, 6a | |
| 2 | CH2 | 29.9 | 2.42/2.66 (2H, m) | H-1 | C-3, 4, 1a | |
| 3 | CH | 123.7 | 5.68 (1H, m) | H-2 | H-4 | C-2, 10 |
| 4 | CH | 121.3 | 6.82 (1H, dd, 3.19, 9.75 Hz) | H-2, 3 | C-2 | |
| 5 | C | 151.7 | ||||
| 6 | CH | 96.0 | 6.44 (1H, s) | H-OMe | C-5, 7, 8, 10 | |
| 7 | C | 152.6 | ||||
| 8 | C | 140.4 | ||||
| 9 | C | 124.6 | ||||
| 10 | C | 117.1 | ||||
| 1a | C | 132.4 | ||||
| 2a | C | 150.8 | ||||
| 3a | CH | 98.0 | 6.30 (1H, s) | H-OMe | C-2a, 4a | |
| 4a | C | 148.0 | ||||
| 5a | C | 142.5 | ||||
| 6a | CH | 114.4 | 6.53 (1H, s) | C-1, 2a, 4a, 5a | ||
| 8-OMe | CH3 | 60.6 | 3.39 (3H, s) | C-8 | ||
| 5a-OMe | CH3 | 57.0 | 3.57 (3H, s) | H-6a | C-5a | |
| 4xOMe | CH3 | 56.9, 56.4, 56.2, 56.1 | 3.84, 3.86, 3.87, 3.91 (each 3H, s) | H-6, 3a | C-5, 7, 2a, 4a | |
| Group of Samples | Cell Viability (in%, Mean ± SD) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 0.1 µg/mL | 1 µg/mL | 10 µg/mL | |||||||
| Methanol Fraction | 97.15 | ± | 14.97 | 114.01 | ± | 48.88 | 118.99 | ± | 30.94 |
| Petroleum Ether Fraction | 67.67 | ± | 8.09 | 72.33 | ± | 8.78 | 76.60 | ± | 12.25 |
| Subfraction 1 | 101.75 | ± | 15.31 | 104.48 | ± | 18.09 | 106.23 | ± | 17.42 |
| Subfraction 2 | 103.15 | ± | 52.06 | 104.41 | ± | 12.63 | 108.90 | ± | 23.77 |
| Subfraction 3 | 118.42 | ± | 6.28 | 97.23 | ± | 10.37 | 81.20 | ± | 9.80 |
| Subfraction 4 | 118.66 | ± | 19.22 | 95.83 | ± | 12.79 | 87.17 | ± | 0.17 |
| Subfraction 5 | 80.47 | ± | 21.69 | 91.55 | ± | 20.19 | 97.91 | ± | 29.11 |
| Subfraction 6 | 124.44 | ± | 23.11 | 106.25 | ± | 0.67 | 101.85 | ± | 47.25 |
| Subfraction 7 | 104.56 | ± | 12.10 | 107.05 | ± | 30.85 | 115.53 | ± | 18.68 |
| Subfraction 8 | 108.36 | ± | 0.87 | 109.53 | ± | 17.14 | 112.90 | ± | 33.80 |
| Subfraction 9 | 111.34 | ± | 6.68 | 110.83 | ± | 6.07 | 98.07 | ± | 27.05 |
| Derivate of Dillapiole | 113.95 | ± | 18.36 | 107.08 | ± | 19.05 | 101.65 | ± | 19.26 |
| Pachypostaudin B | 105.43 | ± | 12.92 | 86.94 | ± | 8.93 | 82.06 | ± | 14.77 |
| Pellucidin A | 88.82 | ± | 2.65 | 107.92 | ± | 11.05 | 125.56 | ± | 23.10 |
| Dillapiole | 89.88 | ± | 19.04 | 105.00 | ± | 11.97 | 104.96 | ± | 4.49 |
| Apiol | 116.88 | ± | 5.39 | 110.04 | ± | 33.57 | 104.68 | ± | 23.58 |
| Compounds | Hydrogen Bonding (ER α) | Hydrogen Bonding (ER β) | ||||
|---|---|---|---|---|---|---|
| ΔG Bind (kcal/mol) | Amino Acid Residues | H-Bond Distance (Å) | ΔG Bind (kcal/mol) | Amino Acid Residues | H-Bond Distance (Å) | |
| 17β-Estradiol | −9.55 | ARG 394 | 2.26 | −9.55 | ARG 346 | 2.05 |
| HIS 524 | 2.24 | HIS 475 | 2.16 | |||
| Derivate of Dillapiole | −5.45 | - | - | −5.29 | LEU 298 | 2.09 |
| Pachypostaudin B | −8.17 | HIS-524 | 3.02 | −7.03 | HIS-475 | 1.91 |
| LEU-476 | 2.74 | |||||
| Pellucidin A | −8.20 | HIS-524 | 3.15 | −7.65 | - | - |
| Dillapiole | −5.14 | - | - | −5.26 | LEU 476 | 3.09 |
| Apiol | −5.58 | ARG-394 | 3.01 | −5.44 | ARG 346 | 2.11 |
Sample Availability: Samples of the compounds are available from the authors. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kartika, I.G.A.A.; Bang, I.J.; Riani, C.; Insanu, M.; Kwak, J.H.; Chung, K.H.; Adnyana, I.K. Isolation and Characterization of Phenylpropanoid and Lignan Compounds from Peperomia pellucida [L.] Kunth with Estrogenic Activities. Molecules 2020, 25, 4914. https://doi.org/10.3390/molecules25214914
Kartika IGAA, Bang IJ, Riani C, Insanu M, Kwak JH, Chung KH, Adnyana IK. Isolation and Characterization of Phenylpropanoid and Lignan Compounds from Peperomia pellucida [L.] Kunth with Estrogenic Activities. Molecules. 2020; 25(21):4914. https://doi.org/10.3390/molecules25214914
Chicago/Turabian StyleKartika, I Gusti Agung Ayu, In Jae Bang, Catur Riani, Muhamad Insanu, Jong Hwan Kwak, Kyu Hyuck Chung, and I Ketut Adnyana. 2020. "Isolation and Characterization of Phenylpropanoid and Lignan Compounds from Peperomia pellucida [L.] Kunth with Estrogenic Activities" Molecules 25, no. 21: 4914. https://doi.org/10.3390/molecules25214914
APA StyleKartika, I. G. A. A., Bang, I. J., Riani, C., Insanu, M., Kwak, J. H., Chung, K. H., & Adnyana, I. K. (2020). Isolation and Characterization of Phenylpropanoid and Lignan Compounds from Peperomia pellucida [L.] Kunth with Estrogenic Activities. Molecules, 25(21), 4914. https://doi.org/10.3390/molecules25214914