A Rose Extract Protects the Skin against Stress Mediators: A Potential Role of Olfactory Receptors

 , , and
, , and
Abstract

1. Introduction
2. Results
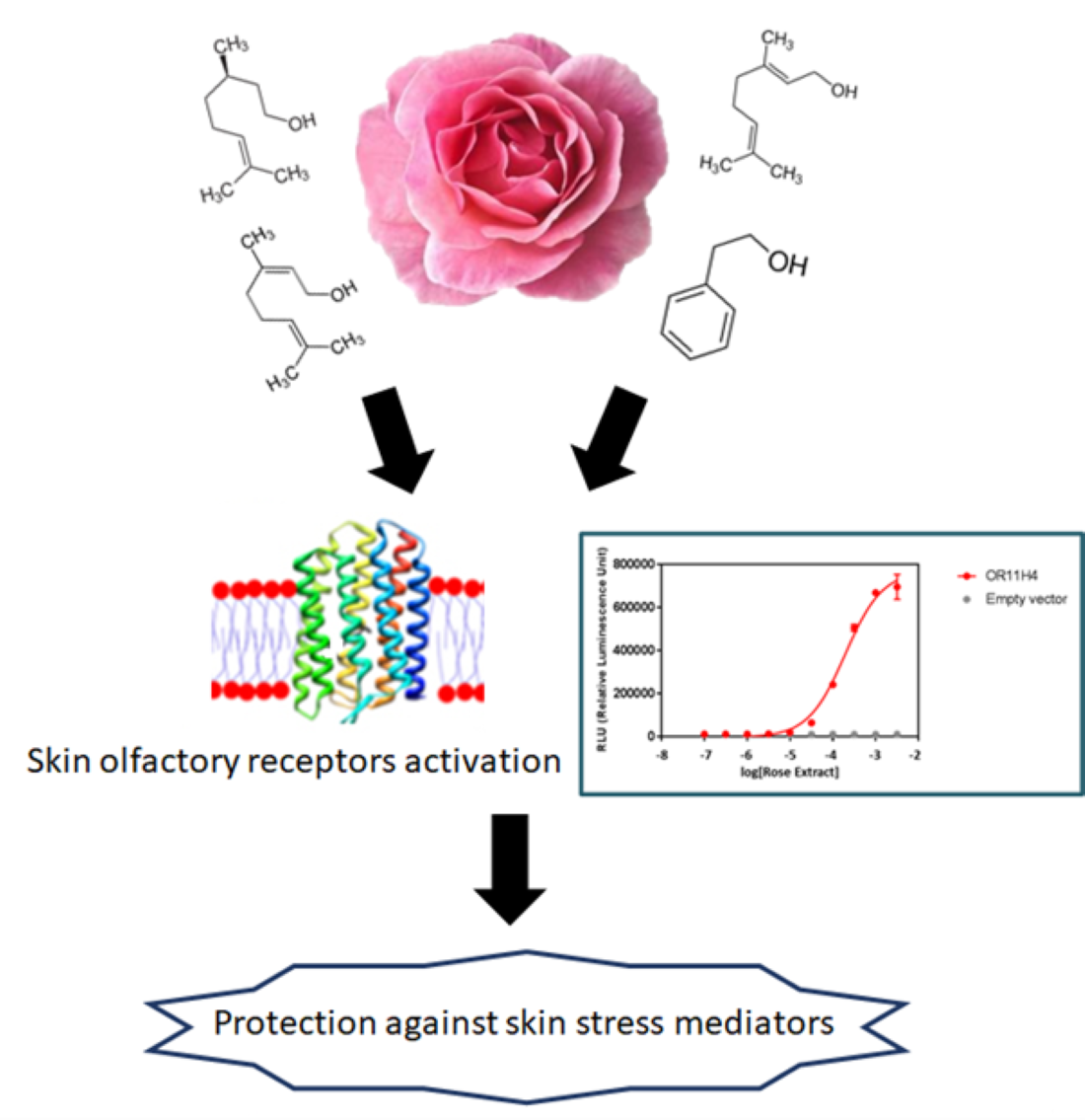
2.1. Chemical Composition of Rose Extract
2.2. ORs are Expressed on Human Keratinocytes and Skin Explants
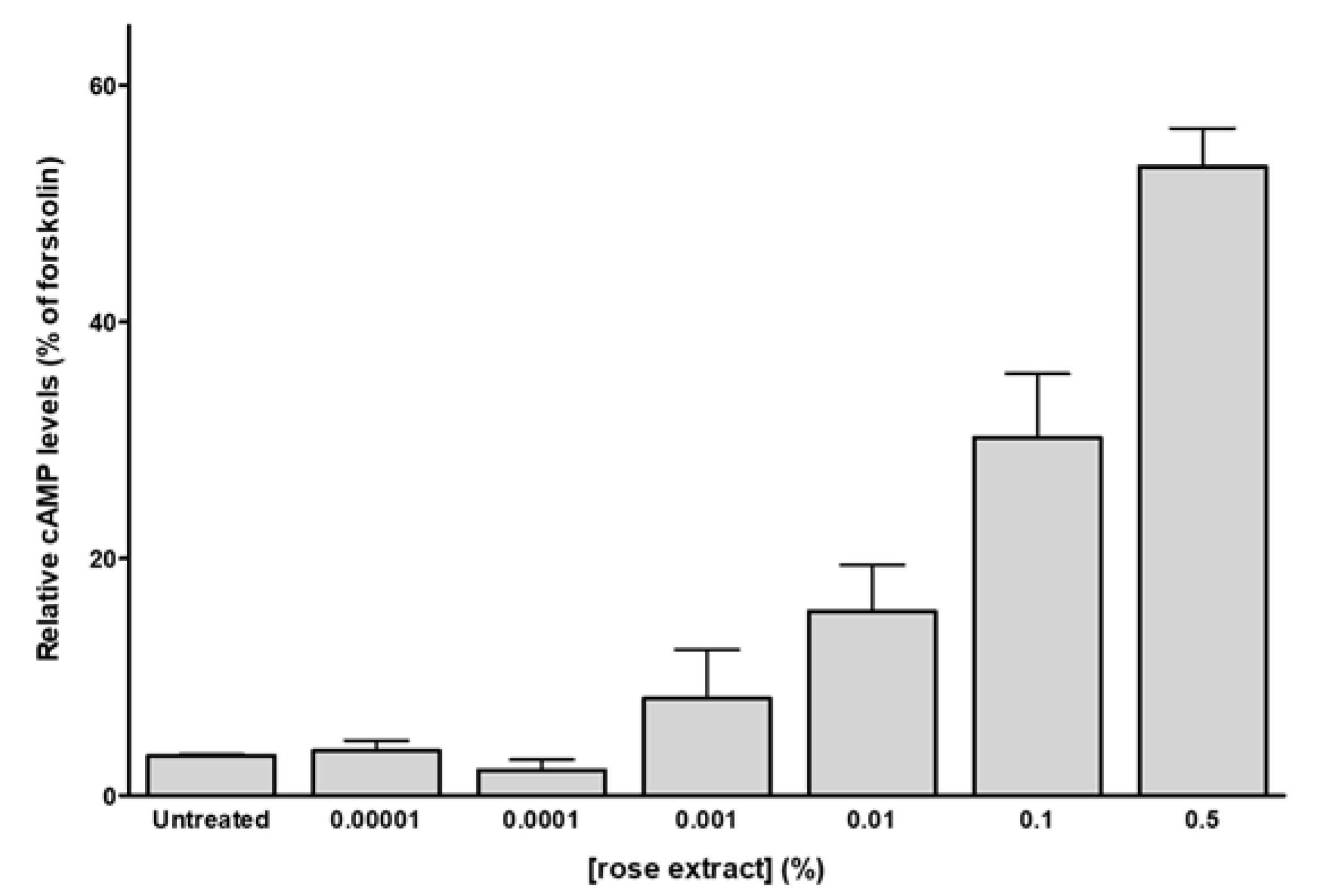
2.3. Rose Extract and Phenylethyl Alcohol Activate Skin Human Olfactory Receptors
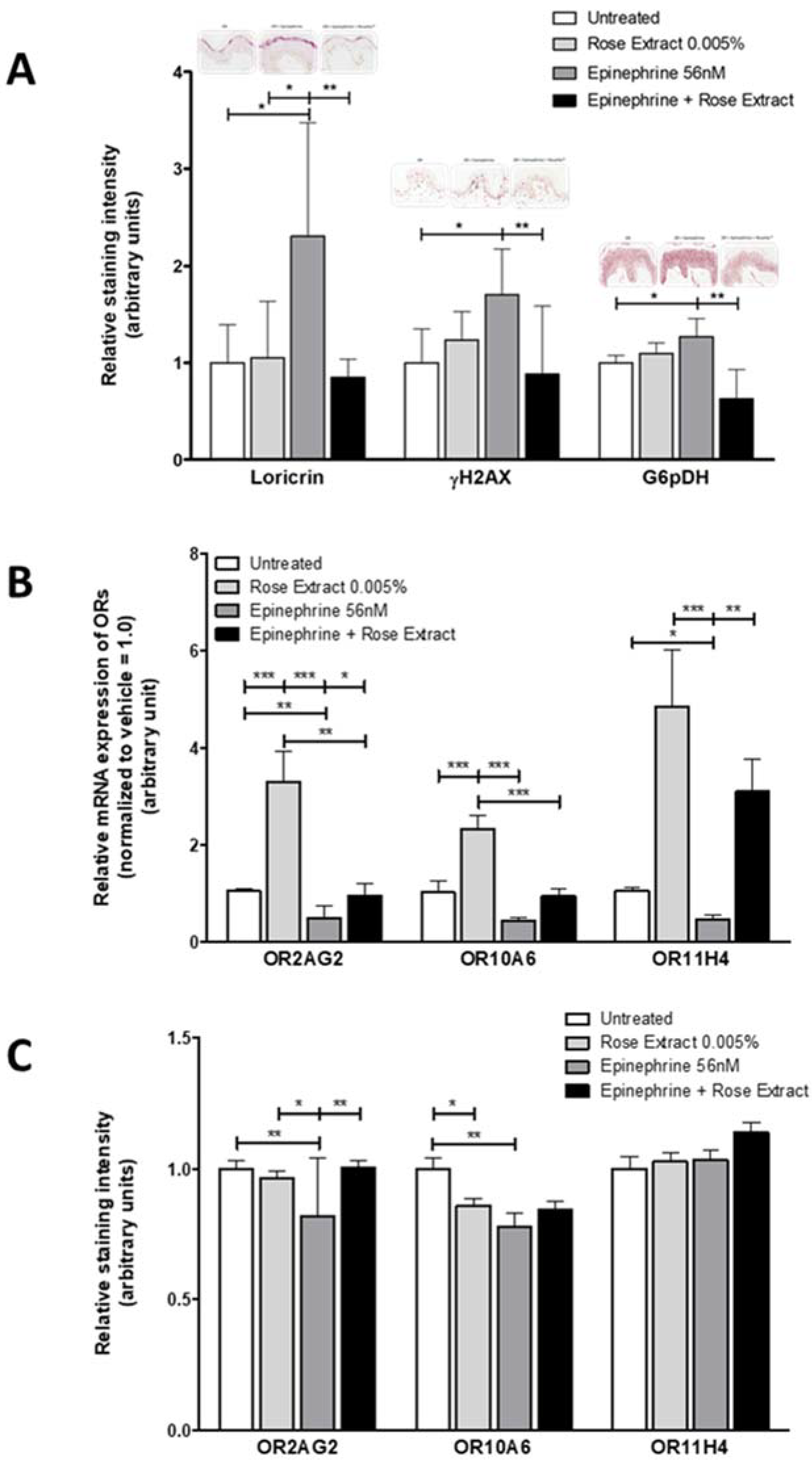
2.4. OR Agonists Restore Olfactory Receptor Expression Following Downregulation with Epinephrine and Protects the Skin against Stress
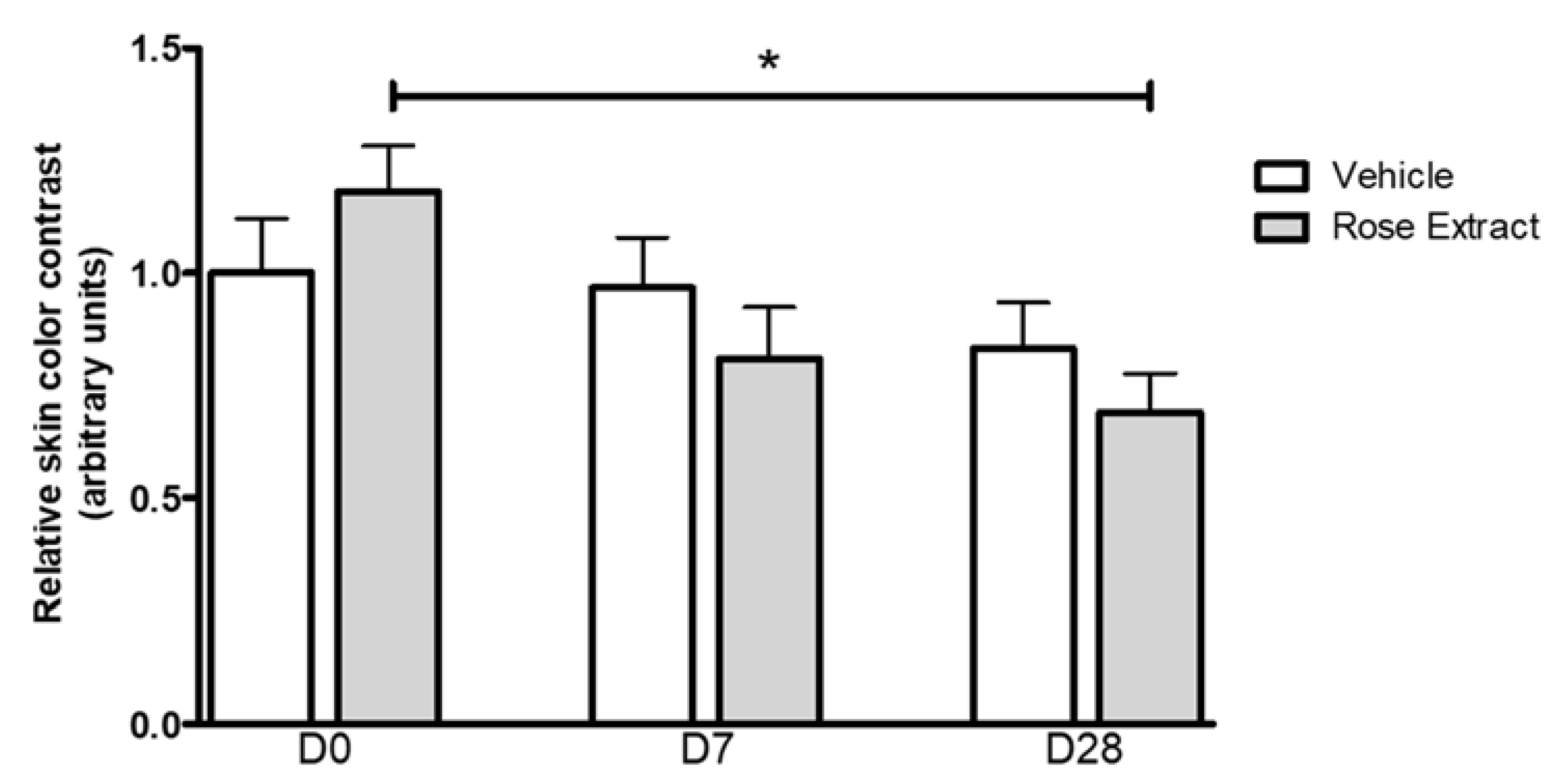
2.5. In Vivo Effects of Rose Extract Showing a Reduction of Under-Eye Dark Circle Appearance
3. Discussion
4. Materials and Methods
4.1. Rose Extract and Chemicals
4.2. Rose Extract Analysis
4.3. Keratinocytes Cell Culture and Skin Explants
4.4. Quantitative RT-PCR
4.5. Immunocytochemistry (ICC) and Immunohistochemistry (IHC)
4.6. CRE-Luciferase Assay
4.7. cAMP-GloTM Assay
4.8. Chemical Stimulation of Human Microdissected Skin
4.9. Clinical Evaluation of Rose Extract on Stress-Induced Skin Fatigue
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| RT-qPCR | Quantitative reverse transcription-polymerase chain reaction |
| G6PD | Glucose-6-phosphate dehydrogenase |
| γH2AX | H2A histone family member X |
| cAMP | Cyclic adenosine monophosphate |
| DNA | Deoxyribonucleic acid |
| RNA | Ribonucleic acid |
| DMEM | Dulbecco’s Modified Eagle Medium |
| KGM | Keratinocyte Growth Medium |
| ROS | Reactive Oxygen Species |
References
- Buck, L.; Axel, R. A novel multigene family may encode odorant receptors: A molecular basis for odor recognition. Cell 1991, 1, 175–187. [Google Scholar] [CrossRef]
- Teixeira, C.S.S.; Cerqueira, N.M.F.S.A.; Ferreira, A.C.S. Unravelling the Olfactory Sense: From the Gene to Odor Perception. Chem. Senses 2015, 41, 105–121. [Google Scholar]
- Antunes, G.; De Souza, F.M.S. Olfactory receptor signaling. Methods Cell Biol. 2016, 132, 127–145. [Google Scholar] [PubMed]
- Dibattista, M.; Reisert, J. The odorant receptor-dependent role of olfactory marker protein in olfactory receptor neurons. J. Neurosci. 2016, 36, 2995–3006. [Google Scholar] [CrossRef] [PubMed]
- Persuy, M.A.; Sanz, G.; Tromelin, A.; Thomas-Danguin, T.; Gibrat, J.F.; Pajot-Augy, E. Mammalian olfactory receptors: Molecular mechanisms of odorant detection, 3D-modeling, and structure-activity relationships. Prog. Mol. Biol. Transl. Sci. 2015, 130, 1–36. [Google Scholar] [PubMed]
- Menashe, I.; Man, O.; Lancet, D.; Gilad, Y. Different noses for different people. Nat. Genet. 2003, 34, 143–144. [Google Scholar] [CrossRef]
- McRae, J.F.; Mainland, J.D.; Jaeger, S.R.; Adipietro, K.A.; Matsunami, H.; Newcomb, R.D. Genetic variation in the odorant receptor OR2J3 is associated with the ability to detect the “grassy” smelling odor, cis-3-hexen-1-ol. Chem. Senses 2012, 37, 585–593. [Google Scholar] [CrossRef]
- MaBberg, D.; Hatt, H. Human Olfactory Receptors: Novel Cellular Functions Outside of the Nose. Physiol. Rev. 2018, 98, 1739–1763. [Google Scholar] [CrossRef]
- Lee, S.J.; Depoortere, I.; Hatt, H. Therapeutic potential of ectopic olfactory and taste receptors. Nat. Rev. Drug Discov. 2019, 18, 116–138. [Google Scholar] [CrossRef]
- Xu, L.L.; Stackhouse, B.G.; Florence, K.; Zhang, W.; Shanmugam, N. PSGR, a novel prostate-specific gene with homology to a G proteincoupled receptor, is overexpressed in prostate cancer. Cancer Res. 2000, 60, 6568–6572. [Google Scholar]
- Gaudin, J.C.; Breuils, L.; Haertlé, T. Mouse orthologs of human olfactory-like receptors expressed in the tongue. Gene 2006, 381, 42–48. [Google Scholar] [CrossRef] [PubMed]
- De la Cruz, O.; Blekhman, R.; Zhang, X.; Nicolae, D.; Firestein, S. A signature of evolutionary constraint on a subset of ectopically expressed olfactory receptor genes. Mol. Biol. Evol. 2009, 26, 491–494. [Google Scholar] [CrossRef]
- Garcia-Esparcia, P.; Schluter, A.; Carmona, M.; Moreno, J.; Ansoleaga, B.; Torrejón-Escribano, B.; Gustincich, S.; Pujol, A.; Ferrer, I. Functional genomics reveals dysregulation of cortical olfactory receptors in Parkinson disease: Novel putative chemoreceptors in the human brain. J. Neuropathol. Exp. Neurol. 2013, 72, 524–539. [Google Scholar] [CrossRef] [PubMed]
- Braun, T.; Voland, P.; Kunz, L.; Prinz, C.; Gratzl, M. Enterochromaffin cells of the human gut: Sensors for spices and odorants. Gastroenterology 2007, 132, 1890–1901. [Google Scholar] [CrossRef]
- Chéret, J.; Bertolini, M.; Ponce, L.; Lehmann, J.; Tsai, T.; Alam, M.; Hatt, H.; Paus, R. Olfactory receptor OR2AT4 regulates human hair growth. Nat. Commun. 2018, 9, 3624. [Google Scholar] [CrossRef]
- Busse, D.; Kudella, P.; Grüning, N.M.; Gisselmann, G.; Staünder, S.; Luger, T.; Jacobsen, F.; Steinstraüer, L.; Paus, R.; Gkogkolou, P.; et al. A synthetic sandalwood odorant induces wound-healing processes in human keratinocytes via the olfactory receptor OR2AT4. J. Investig. Dermatol. 2014, 134, 2823–2832. [Google Scholar] [CrossRef] [PubMed]
- Tsai, T.; Veitinger, S.; Peek, I.; Busse, D.; Eckardt, J.; Vladimirova, D.; Jovancevic, N.; Wojcik, W.; Gisselmann, G.; Altmüller, J.; et al. Two olfactory receptors-OR2A4/7 and OR51B5-differentially affect epidermal proliferation and differentiation. Exp. Dermatol. 2016, 26, 58–65. [Google Scholar] [CrossRef]
- Gelis, L.; Jovancevic, N.; Veitinger, S.; Mandal, B.; Arndt, H.D.; Neuhaus, E.M.; Hatt, H. Functional Characterization of the Odorant Receptor 51E2 in Human Melanocytes. J. Biol. Chem. 2016, 291, 17772–17786. [Google Scholar] [CrossRef]
- Wojcik, S.; Weidinger, D.; Ständer, S.; Luger, T.; Hatt, H.; Jovancevic, N. Functional characterization of the extranasal OR2A4/7 expressed in human melanocytes. Exp. Dermatol. 2018, 11, 1216–1223. [Google Scholar] [CrossRef]
- Flint, M.S.; Baum, A.; Chambers, W.H.; Jenkins, F.J. Induction of DNA damage, alteration of DNA repair and transcriptional activation by stress hormones. Psychoneuroendocrinology 2007, 32, 470–479. [Google Scholar] [CrossRef]
- Sivamani, R.K.; Pullar, C.E.; Manabat-Hidalgo, C.G.; Rocke, D.M.; Carlsen, R.C.; Greenhalgh, D.G.; Isseroff, R.R. Stress-mediated increases in systemic and local epinephrine impair skin wound healing: Potential new indication for beta blockers. PLoS ONE 2009, 6, e12. [Google Scholar] [CrossRef] [PubMed]
- Makoto, R.H.; Jeffrey, J.K.; Erin, J.; Ryan, T.; Grant, W.; Towers, A.J.; Williams, B.; Lam, C.M.; Xiao, K.; Shenoy, S.K.; et al. A stress response pathway regulates DNA damage through β2-adrenoreceptors and β-arrestin-1. Nature 2011, 477, 349–353. [Google Scholar]
- Kim, M.H.; Gorouhi, F.; Ramirez, J.L.; Granick, B.; Soulika, A.M.; Simon, S.; Rivkah, I. Catecholamine stress alters neutrophil trafficking and impairs wound healing by β2-adrenergic receptor-mediated upregulation of IL-6. J. Investig. Dermatol. 2014, 134, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Hongratanaworakit, T. Relaxing Effect of Rose Oil on Humans. Nat. Prod. Commun. 2009, 4, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Angelucci, F.L.; Silva, V.V.; Dal Pizzol, C.; Spir, L.G.; Praes, C.E.O.; Maibach, H. Physiological effect of olfactory stimuli inhalation in humans: An overview. Int. J. Cosm. Sci. 2014, 36, 117–123. [Google Scholar] [CrossRef]
- Stanton, R.C. Glucose-6-phosphate dehydrogenase, NADPH, and cell survival. IUBMB Life 2012, 64, 362–369. [Google Scholar] [CrossRef]
- Tiwari, M. Glucose 6 phosphatase dehydrogenase (G6PD) and neurodegenerative disorders: Mapping diagnostic and therapeutic opportunities. Genes Dis. 2017, 4, 196–203. [Google Scholar] [CrossRef]
- Esser, C.; Rannug, A. The aryl hydrocarbon receptor in barrier organ physiology, immunology, and toxicology. Pharmacol Rev. 2015, 67, 259–279. [Google Scholar] [CrossRef]
- Furue, M.; Takahara, M.; Nakahara, T.; Uchi, H. Role of AhR/ARNT system in skin homeostasis. Arch. Dermatol. Res. 2014, 306, 769–779. [Google Scholar] [CrossRef]
- Mah, L.J.; El-Osta, A.; Karagiannis, T.C. GammaH2AX: A sensitive molecular marker of DNA damage and repair. Leukemia 2010, 24, 679–686. [Google Scholar] [CrossRef]
- Dickey, J.S.; Redon, C.E.; Nakamura, A.J.; Baird, B.J.; Sedelnikova, O.A.; Bonner, W.M. H2AX: Functional roles and potential applications. Chromosoma 2009, 118, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Jovancevic, N.; Wunderlich, K.A.; Hearing, C.; Flegel, C.; Maßberg, D.; Weinrich, M.; Weber, L.; Tebbe, L.; Kampik, A.; Gisselmann, G.; et al. Deep Sequencing of the Human Retinae Reveals the Expression of Odorant Receptors. Front. Cell. Neurosci. 2017, 11, 3. [Google Scholar] [CrossRef]
- Flegel, C.; Vogel, F.; Hofreuter, A.; Schreiner, B.S.P.; Osthold, S.; Veitinger, S.; Becker, C.; Brockmeyer, N.H.; Muschol, M.; Wennemuth, G.; et al. Characterization of the Olfactory Receptors Expressed in Human Spermatozoa. Front. Mol. Biosci. 2015, 2, 73. [Google Scholar] [CrossRef]
- Mainland, J.D.; Li, Y.R.; Zhou, T.; Liu, W.L.; Matsunami, H. Human olfactory receptor responses to odorants. Sci. Data 2015, 2, 150002. [Google Scholar] [CrossRef] [PubMed]
- Pfister, P.; Matthew, E.R.; Parris, R.J. Methods of Identifying, Isolating and Using Odorant and Aroma Receptors. WO2014210585, 31 December 2014. [Google Scholar]
- Chatelain, P.; Wilkin, F.; Veithen, A.; Philippeau, M.; Noiset, O.; Quesnel, Y. Sensorial Chemistry: Functional characterization and structure-activity relationship understanding of OR5K1 and OR2AG1 allow to design and synthetize new selective compounds. Chem. Senses 2017, 42, e28. [Google Scholar]
- Mohebitabar, S.; Shirazi, M.; Bioos, S.; Rahimi, R.; Malekshahi, F.; Nejatbakhsh, F. Therapeutic efficacy of rose oil: A comprehensive review of clinical evidence. Avicenna. J. Phytomed. 2017, 7, 206–213. [Google Scholar]
- Fukada, M.; Kano, E.; Miyoshi, M.; Komaki, R.; Watanabe, T. Effect of ‘‘Rose Essential Oil’’ Inhalation on Stress-Induced Skin-Barrier, Disruption in Rats and Humans. Chem. Senses 2012, 37, 347–356. [Google Scholar] [CrossRef]
- Igarashi, M.; Ikei, H.; Song, C.; Miyazaki, Y. Effects of olfactory stimulation with rose and orange oil on prefrontal cortex activity. Complement Ther. Med. 2014, 22, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Yang, H.; Kim, D.; Yang, M.; Park, T.J.; Hong, S. Human-like smelling of a rose scent using an olfactory receptor nanodisc-based bioelectronic nose. Sci. Rep. 2018, 8, 13945. [Google Scholar] [CrossRef]
- Saito, N.; Yamano, E.; Ishii, A.; Tanaka, M.; Nakamura, J.; Watanabe, Y. Involvement of the olfactory system in the induction of anti-fatigue effects by odorants. PLoS ONE 2018, 13, e0195263. [Google Scholar] [CrossRef]
- Chakraborty, S.; Dakle, P.; Sinha, A.; Vishweswaraiah, S.; Nagori, A.; Salimath, S.; Prakash, Y.S.; Lodha, R.; Kabra, S.K.; Ghosh, B.; et al. Genetic variations in olfactory receptor gene OR2AG2 in a large multigenerational family with asthma. Sci. Rep. 2019, 9, 19029. [Google Scholar] [CrossRef] [PubMed]
- Hou, G.; Arias-Carrion, O. Chronic Stress Impacts on Olfactory System. CNS Neurol. Disord.–Drug Targets 2015, 14, 486–491. [Google Scholar]
- Hoenen, M.; Wolf, O.T.; Pause, B.M. The Impact of Stress on Odor Perception. Perception 2017, 46, 366–376. [Google Scholar] [CrossRef] [PubMed]
- Greenbaum, D.; Colangelo, C.; Williams, K.; Gerstein, M. Comparing protein abundance and mRNA expression levels on a genomic scale. Genome Biol. 2003, 4, 117. [Google Scholar] [CrossRef]
- Hague, C.; Uberti, M.A.; Chen, Z.; Bush, C.F.; Jones, S.V.; Ressler, K.J.; Hall, R.A.; Minneman, K.P. Olfactory receptor surface expression is driven by association with the β2-adrenergic receptor. Proc. Natl. Acad. Sci. USA 2004, 101, 13672–13676. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Pauer, S.H.; Yong, H.M.; An, S.S.; Liggett, S.B. β2-Adrenergic Receptors Chaperone Trapped Bitter Taste Receptor 14 to the Cell Surface as a Heterodimer and Exert Unidirectional Desensitization of Taste Receptor Function. J. Biol. Chem. 2016, 291, 17616–17628. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Romana-Souza, B.; Silva-Xavier, W.; Monte-Alto-Costa, A. Topical retinol attenuates stress-induced ageing signs in human skin ex vivo, through EGFR activation via EGF, but not ERK and AP-1 activation. Exp. Dermatol. 2018, 28, 906–913. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substance | Rt (min) | Compound | Content (%) |
|---|---|---|---|
| 1 | 7.0 | Hexanol | 0.12 |
| 2 | 8.5 | Heptanal | 0.04 |
| 3 | 11.6 | α-Pinene | 0.15 |
| 4 | 13.5 | Sabinene | 0.03 |
| 5 | 14.3 | Beta pinene | 0.04 |
| 6 | 15.5 | Myrcene | 0.07 |
| 7 | 17.1 | Benzyl Alcohol | 0.55 |
| 8 | 56.7 | Phenylethyl alcohol | 56.70 |
| 9 | 24.0 | Cis-Rose oxide | 0.12 |
| 10 | 25.1 | Trans-Rose oxide | 0.05 |
| 11 | 28.2 | Terpinen-4-ol | 0.23 |
| 12 | 29.0 | α-Terpineol | 0.10 |
| 13 | 32.4 | Nerol | 4.63 |
| 14 | 32.4 | Citronellol | 16.70 |
| 15 | 32.9 | Phenethyl acetate | 0.50 |
| 16 | 33.9 | Geraniol | 8.90 |
| 17 | 34.1 | Geranial | 0.25 |
| 18 | 35.1 | Citronellyl Formate | 0.02 |
| 19 | 36.7 | Geranyl Formate | 0.01 |
| 20 | 39.4 | Eugenol | 0.15 |
| 21 | 40.1 | Citronellyl Acetate | 0.40 |
| 22 | 41.6 | Neryl Acetate | 0.08 |
| 23 | 41.7 | Geranyl Acetate | 1.06 |
| 24 | 42.4 | β-Bourbonen | 0.05 |
| 25 | 42.9 | β-Elemen | 0.08 |
| 26 | 44.4 | β-Caryophyllene | 0.25 |
| 27 | 45.7 | α-Guaiene | 0.23 |
| 28 | 46.3 | α-Humulene | 0.16 |
| 29 | 47.8 | Germacrene D | 0.25 |
| 30 | 48.9 | (−)-α-Selinene | 0.06 |
| 31 | 49.3 | α-Bulnesene | 0.17 |
| 32 | 50.0 | Pentadecane | 0.11 |
| 33 | 69.1 | Nonadecene | 0.80 |
| 34 | 70.7 | Nonadecane | 3.80 |
| Entry | Molecules | EC50 (µM) | ||
|---|---|---|---|---|
| OR10A6 | OR2AG2 | OR11H4 | ||
| 1 | Citronellol | 2.0 | 13.5 | NA |
| 2 | Nerol | 6.0 | 51.0 | NA |
| 3 | Cis-3-Hexenol | NA | 98.8 | NA |
| 4 | Benzyl alcohol | NA | NA | NA |
| 5 | Linalool | 5.6 | 56.2 | NA |
| 6 | Geraniol | 3.3 | 18.0 | NA |
| 7 | α-Cinnamyl alcohol | 1.31 | 8.43 | NA |
| 8 | Cyclamen aldehyde | 9.62 | NA | NA |
| 9 | Lyral | 25.47 | NA | NA |
| 10 | α-Damascone | NA | NA | NA |
| 11 | α-Ionone | 7.48 | NA | NA |
| 12 | Phenyl ethyl alcohol | 114.5 | 151.7 | 20.3 |
| 13 | Phenyl propyl alcohol | 49.32 | 103.04 | 52.84 |
| 14 | β-Ionone | NA | NA | NA |
| 15 | Lavandulol | NA | NA | NA |
| 16 | Benzyl acetone | 19.10 | 5.79 | NA |
| 17 | Cyclemone A | 18.66 | NA | NA |
| 18 | Nonadecane | 119.8 | NA | NA |
| 19 | Rose extract | 1.51 × 10−4 * | 8.12 × 10−4 * | 1.65 × 10−4 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duroux, R.; Mandeau, A.; Guiraudie-Capraz, G.; Quesnel, Y.; Loing, E. A Rose Extract Protects the Skin against Stress Mediators: A Potential Role of Olfactory Receptors. Molecules 2020, 25, 4743. https://doi.org/10.3390/molecules25204743
Duroux R, Mandeau A, Guiraudie-Capraz G, Quesnel Y, Loing E. A Rose Extract Protects the Skin against Stress Mediators: A Potential Role of Olfactory Receptors. Molecules. 2020; 25(20):4743. https://doi.org/10.3390/molecules25204743
Chicago/Turabian StyleDuroux, Romain, Anne Mandeau, Gaelle Guiraudie-Capraz, Yannick Quesnel, and Estelle Loing. 2020. "A Rose Extract Protects the Skin against Stress Mediators: A Potential Role of Olfactory Receptors" Molecules 25, no. 20: 4743. https://doi.org/10.3390/molecules25204743
APA StyleDuroux, R., Mandeau, A., Guiraudie-Capraz, G., Quesnel, Y., & Loing, E. (2020). A Rose Extract Protects the Skin against Stress Mediators: A Potential Role of Olfactory Receptors. Molecules, 25(20), 4743. https://doi.org/10.3390/molecules25204743