In Vitro Digestion and Fermentation by Human Fecal Microbiota of Polysaccharides from Flaxseed


Abstract

1. Introduction
2. Results and Discussion
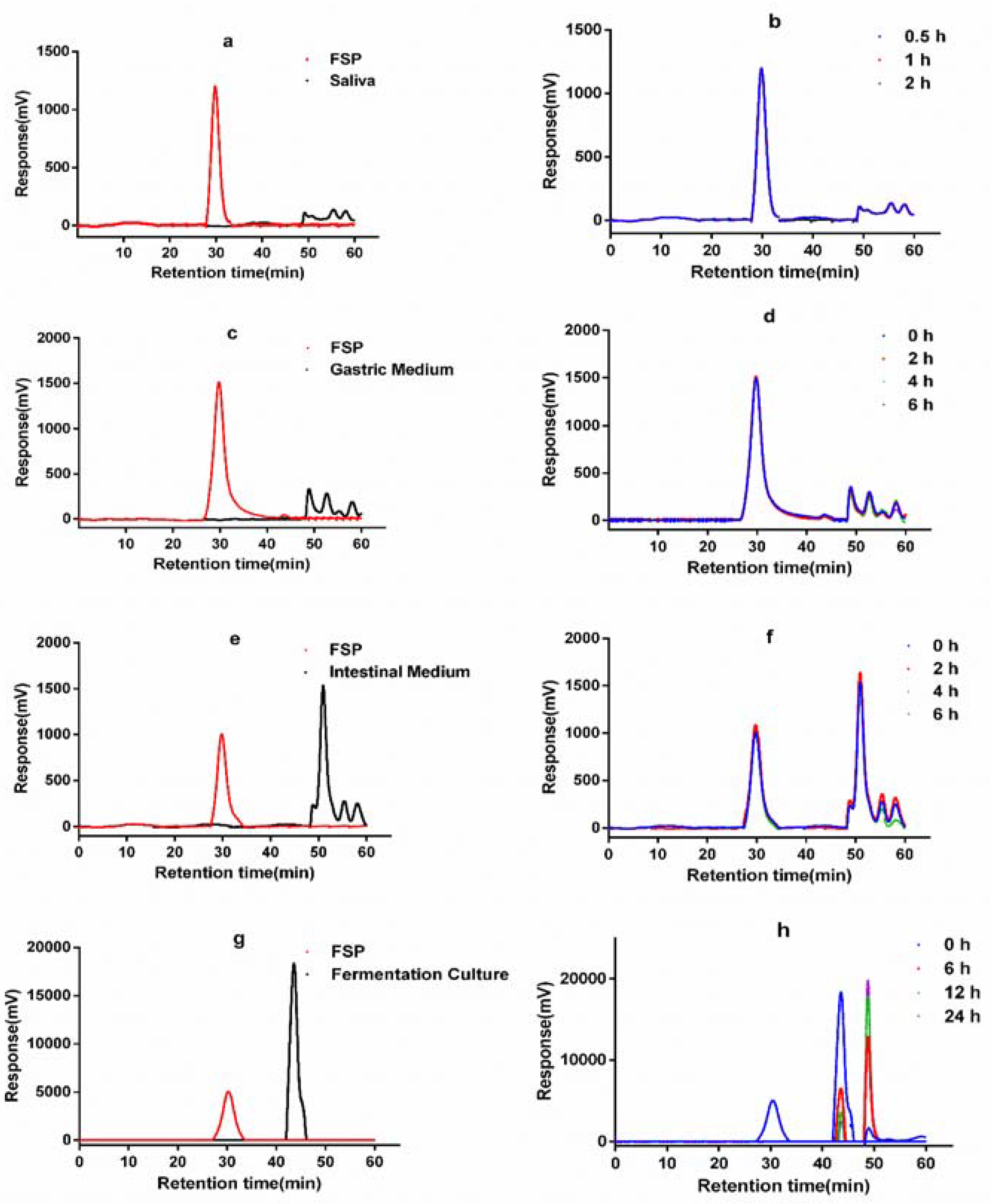
2.1. Change of FSP on Digestion in Simulated Saliva
2.2. Change of FSP on Simulated Gastric Digestion
2.3. Change of FSP in Simulated Intestinal Digestion
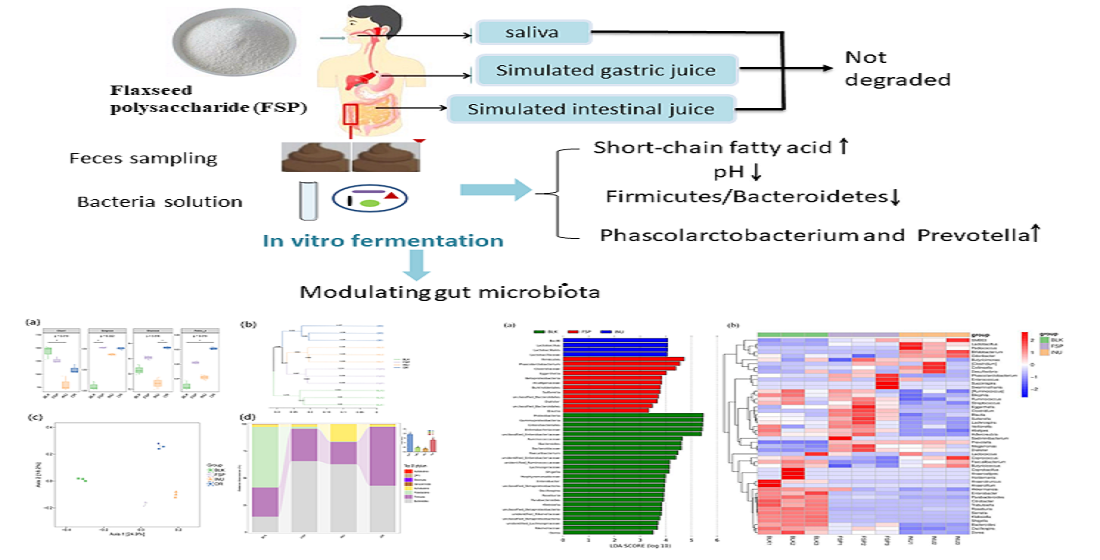
2.4. Change of FSP in In Vitro Fermentation by Human Gut Microbiota
2.5. Effects of FSP on SCFA Production
2.6. Effects of FSP on Gut Microbiota
3. Materials and Methods
3.1. Materials
3.2. Preparation of Polysaccharide
3.3. Simulated Saliva Digestion
3.4. Simulated Gastric Digestion
3.5. Simulated Small Intestinal Digestion
3.6. In Vitro Fermentation of FSP
3.6.1. Preparation of Fermentation Medium
3.6.2. Preparation of Fecal Slurry and Fermentation
3.7. Determinations of pH, Carbohydrate Content and Reducing Sugar
3.8. Determination of SCFA Content
3.9. DNA Extraction and Analysis
3.10. Molecular Weight Determination
3.11. Determination of Free Monosaccharide and Fourier Transform Infrared (FT-IR) Spectra
3.12. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gill, S.R.; Pop, M.; Deboy, R.T.; Eckburg, P.B.; Turnbaugh, P.J.; Samuel, B.S.; Gordon, J.I.; Relman, D.A.; Fraser-Liggett, C.M.; Nelson, K.E. Metagenomic analysis of the human distal gut microbiome. Science 2006, 312, 1355–1359. [Google Scholar] [CrossRef]
- Donaldson, G.P.; Lee, S.M.; Mazmanian, S.K. Gut biogeography of the bacterial microbiota. Nat. Rev. Microbiol. 2016, 14, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Marialetizia, R.; Cani, P.D.; Claude, K. The Gut Microbiome Influences Host Endocrine Functions. Endocr. Rev. 2018, 40, 1271–1284. [Google Scholar]
- Rao, T.P.; Quartarone, G. Role of guar fiber in improving digestive health and function. Nutrition 2019, 59, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef] [PubMed]
- Bindels, L.B.; Delzenne, N.M.; Cani, P.D.; Walter, J. Towards a more comprehensive concept for prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, A.; Liu, P.; Li, Z. Effects of Fuzhuan Brick-Tea Water Extract on Mice Infected with E. coli O157:H7. Nutrients 2015, 7, 5309–5326. [Google Scholar] [CrossRef]
- Di, T.; Chen, G.; Sun, Y.; Ou, S.; Zeng, X.; Ye, H. In vitro digestion by saliva, simulated gastric and small intestinal juices and fermentation by human fecal microbiota of sulfated polysaccharides from Gracilaria rubra. J. Funct. Foods 2018, 40, 18–27. [Google Scholar] [CrossRef]
- Chen, G.; Xie, M.; Dai, Z.; Wan, P.; Ye, H.; Zeng, X.; Sun, Y. Kudingcha and Fuzhuan Brick Tea Prevent Obesity and Modulate Gut Microbiota in High-Fat Diet Fed Mice. Mol. Nutr. Food Res. 2018, 62, 1700485. [Google Scholar] [CrossRef]
- Wang, X.; Wang, X.; Jiang, H.; Cai, C.; Li, G.; Hao, J.; Yu, G. Marine polysaccharides attenuate metabolic syndrome by fermentation products and altering gut microbiota: An overview. Carbohydr. Polym. 2018, 195, 601–612. [Google Scholar] [CrossRef]
- Ren, Y.; Geng, Y.; Du, Y.; Li, W.; Lu, Z.M.; Xu, H.Y.; Xu, G.H.; Shi, J.S.; Xu, Z.H. Polysaccharide of Hericium erinaceus attenuates colitis in C57BL/6 mice via regulation of oxidative stress, inflammation-related signaling pathways and modulating the composition of the gut microbiota. J. Nutr. Biochem. 2018, 57, 67–76. [Google Scholar] [CrossRef]
- Schonfeld, P.; Wojtczak, L. Short- and medium-chain fatty acids in energy metabolism: The cellular perspective. J. Lipid Res. 2016, 57, 943–954. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.T.; Davis-Richardson, A.G.; Giongo, A.; Gano, K.A.; Crabb, D.B.; Mukherjee, N.; Casella, G.; Drew, J.C.; Ilonen, J.; Knip, M.; et al. Gut microbiome metagenomics analysis suggests a functional model for the development of autoimmunity for type 1 diabetes. PLoS ONE 2011, 6, e25792. [Google Scholar] [CrossRef] [PubMed]
- Brooks, L.; Viardot, A.; Tsakmaki, A.; Stolarczyk, E.; Bewick, G.A. Fermentable carbohydrate stimulates FFAR2-dependent colonic PYY cell expansiontoincrease satiety. Mol. Metab. 2017, 6, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Prasad, K.; Dhar, A. Flaxseed and Diabetes. Curr. Pharm. Des. 2016, 22, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Biao, Y.; Nan, H.J.; Yao, L.C.; Jie, S.C.; Chun, D.H.; Mcclements, J.D.; Jiang, C.C. Identification and characterization of antioxidant and immune-stimulatory polysaccharides in flaxseed hull. Food Chem. 2020, 315, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Safdar, B.; Pang, Z.; Liu, X.; Ahmad Jatoi, M.; Mehmood, A.; Tayyab Rashid, M.; Ali, N.; Naveed, M. Flaxseed gum: Extraction, bioactive composition, structural characterization, and its potential antioxidant activity. J. Food Biochem. 2020, 44, e13134. [Google Scholar] [CrossRef]
- Li, D.; Wang, P.; Wang, P.; Hu, X.; Chen, F. The gut microbiota: A treasure for human health. Biotechnol. Adv. 2016, 34, 1210–1224. [Google Scholar] [CrossRef]
- Vieira, J.M.; Andrade, C.C.P.; Santos, T.P.; Okuro, P.K.; Garcia, S.T.; Rodrigues, M.I.; Vicente, A.A.; Cunha, R.L. Flaxseed gum-biopolymers interactions driving rheological behaviour of oropharyngeal dysphagia-oriented products. Food Hydrocoll. 2020, 111, 132–144. [Google Scholar]
- Yang, C.; Xu, Z.; Deng, Q.; Huang, Q.; Wang, X.; Huang, F. Beneficial effects of flaxseed polysaccharides on metabolic syndrome via gut microbiota in high-fat diet fed mice. Food Res. Int. 2020, 131, 108994. [Google Scholar] [CrossRef]
- Pedersen, A.; Sorensen, C.E.; Proctor, G.B.; Carpenter, G.H. Salivary functions in mastication, taste and textural perception, swallowing and initial digestion. Oral Dis. 2018, 24, 1399–1416. [Google Scholar] [CrossRef] [PubMed]
- Stokes, J.R.; Davies, G.A. Viscoelasticity of human whole saliva collected after acid and mechanical stimulation. Biorheology 2007, 44, 141–160. [Google Scholar] [PubMed]
- Liang, S.; Li, X.; Ma, X.; Li, A.; Wang, Y.; Reaney, M.J.T.; Shim, Y.Y. A flaxseed heteropolysaccharide stimulates immune responses and inhibits hepatitis B virus. Int. J. Biol. Macromol. 2019, 136, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Xie, M.; Wan, P.; Chen, D.; Ye, H.; Chen, L.; Zeng, X.; Liu, Z. Digestion under saliva, simulated gastric and small intestinal conditions and fermentation in vitro by human intestinal microbiota of polysaccharides from Fuzhuan brick tea. Food Chem. 2018, 244, 331–339. [Google Scholar] [CrossRef]
- Hu, J.L.; Nie, S.P.; Min, F.F.; Xie, M.Y. Artificial simulated saliva, gastric and intestinal digestion of polysaccharide from the seeds of Plantago asiatica L. Carbohydr. Polym. 2013, 92, 1143–1150. [Google Scholar] [CrossRef]
- Kastl, A.J., Jr.; Terry, N.A.; Wu, G.D.; Albenberg, L.G. The Structure and Function of the Human Small Intestinal Microbiota: Current Understanding and Future Directions. Cell Mol. Gastroenterol. Hepatol. 2020, 9, 33–45. [Google Scholar] [CrossRef]
- Liu, H.; Gong, F.; Wei, F.; Wu, H. Artificial simulation of salivary and gastrointestinal digestion, and fermentation by human fecal microbiota, of polysaccharides from Dendrobium aphyllum. RSC Adv. 2018, 8, 13954–13963. [Google Scholar] [CrossRef]
- Gullón, B.; Gullón, P.; Tavaria, F.; Pintado, M.; Gomes, A.M.; Alonso, J.L.; Parajó, J.C. Structural features and assessment of prebiotic activity of refined arabinoxylooligosaccharides from wheat bran. J. Funct. Foods 2014, 6, 438–449. [Google Scholar] [CrossRef]
- Ma, G.; Kimatu, B.M.; Zhao, L.; Yang, W.; Pei, F.; Hu, Q. In vivo fermentation of a Pleurotus eryngii polysaccharide and its effects on fecal microbiota composition and immune response. Food Funct. 2017, 8, 1810–1821. [Google Scholar] [CrossRef]
- Wu, J.; Zheng, J.; Xia, X.; Kan, J. Purification and Structural Identification of Polysaccharides from Bamboo Shoots (Dendrocalamus latiflorus). Int. J. Mol. Sci. 2015, 16, 15560–15577. [Google Scholar] [CrossRef]
- Correa, V.G.; Goncalves, G.A.; de Sa-Nakanishi, A.B.; Ferreira, I.; Barros, L.; Dias, M.I.; Koehnlein, E.A.; de Souza, C.G.M.; Bracht, A.; Peralta, R.M. Effects of in vitro digestion and in vitro colonic fermentation on stability and functional properties of yerba mate (Ilex paraguariensis A. St. Hil.) beverages. Food Chem. 2017, 237, 453–460. [Google Scholar] [CrossRef]
- Theodoridou, K.; Vail, S.; Yu, P. Explore protein molecular structure in endosperm tissues in newly developed black and yellow type canola seeds by using synchrotron-based Fourier transform infrared microspectroscopy. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2014, 120, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Goormaghtigh, E.; Ruysschaert, J.M.; Raussens, V. Evaluation of the information content in infrared spectra for protein secondary structure determination. Biophys. J. 2006, 90, 2946–2957. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Wu, J.; Zhang, Y.; Chen, H.; Wang, Y. Physicochemical characterization of puerh tea polysaccharides and their antioxidant and α-glycosidase inhibition. J. Funct. Foods 2014, 6, 545–554. [Google Scholar] [CrossRef]
- Al-Lahham, S.H.; Peppelenbosch, M.P.; Roelofsen, H.; Vonk, R.J.; Venema, K. Biological effects of propionic acid in humans; metabolism, potential applications and underlying mechanisms. Biochim. Biophys. Acta 2010, 1801, 1175–1183. [Google Scholar] [CrossRef] [PubMed]
- Canani, R.B.; Costanzo, M.D.; Leone, L.; Pedata, M.; Meli, R.; Calignano, A. Potential beneficial effects of butyrate in intestinal and extraintestinal diseases. World J. Gastroenterol. 2011, 17, 1519–1528. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Hamady, M.; Yatsunenko, T.; Cantarel, B.L.; Duncan, A.; Ley, R.E.; Sogin, M.L.; Jones, W.J.; Roe, B.A.; Affourtit, J.P.; et al. A core gut microbiome in obese and lean twins. Nature 2009, 457, 480–484. [Google Scholar] [CrossRef]
- Fan, W.; Huo, G.; Li, X.; Yang, L.; Duan, C. Impact of diet in shaping gut microbiota revealed by a comparative study in infants during the six months of life. J. Microbiol. Biotechnol. 2014, 24, 133–143. [Google Scholar] [CrossRef]
- Luo, J.; Li, Y.; Mai, Y.; Gao, L.; Ou, S.; Wang, Y.; Liu, L.; Peng, X. Flaxseed gum reduces body weight by regulating gut microbiota. J. Funct. Foods 2018, 47, 136–142. [Google Scholar] [CrossRef]
- Wojciechowicz, M.; Heinrichova, K.; Ziolecki, A. A polygalacturonate lyase produced by Lachnospira multiparus isolated from the bovine rumen. J. Gen. Microbiol. 1980, 117, 193–199. [Google Scholar] [CrossRef]
- Jones, M.L.; Ganopolsky, J.G.; Martoni, C.J.; Labbe, A.; Prakash, S. Emerging science of the human microbiome. Gut Microbes 2014, 5, 446–457. [Google Scholar] [CrossRef] [PubMed]
- Hiippala, K.; Kainulainen, V.; Kalliomaki, M.; Arkkila, P.; Satokari, R. Mucosal Prevalence and Interactions with the Epithelium Indicate Commensalism of Sutterella spp. Front. Microbiol. 2016, 7, 1706. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhao, Y.; Xu, J.; Xue, Z.; Zhang, M.; Pang, X.; Zhang, X.; Zhao, L. Modulation of gut microbiota by berberine and metformin during the treatment of high-fat diet-induced obesity in rats. Sci. Rep. 2015, 5, 14405. [Google Scholar] [CrossRef]
- Li, T.; Long, M.; Gatesoupe, F.J.; Zhang, Q.; Li, A.; Gong, X. Comparative analysis of the intestinal bacterial communities in different species of carp by pyrosequencing. Microb. Ecol. 2015, 69, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Roulard, R.; Petit, E.; Mesnard, F.; Rhazi, L. Molecular investigations of flaxseed mucilage polysaccharides. Int. J. Biol. Macromol. 2016, 86, 840–847. [Google Scholar] [CrossRef]
- Fu, X.; Cao, C.; Ren, B.; Zhang, B.; Huang, Q.; Li, C. Structural characterization and in vitro fermentation of a novel polysaccharide from Sargassum thunbergii and its impact on gut microbiota. Carbohydr. Polym. 2018, 183, 230–239. [Google Scholar] [CrossRef]
- Tedeschi, C.; Clement, V.; Rouvet, M.; Valles-Pamies, B. Dissolution tests as a tool for predicting bioaccessibility of nutrients during digestion. Food Hydrocoll. 2009, 23, 1228–1235. [Google Scholar] [CrossRef]
- Ding, Y.; Yan, Y.; Peng, Y.; Chen, D.; Mi, J.; Lu, L.; Luo, Q.; Li, X.; Zeng, X.; Cao, Y. In vitro digestion under simulated saliva, gastric and small intestinal conditions and fermentation by human gut microbiota of polysaccharides from the fruits of Lycium barbarum. Int. J. Biol Macromol 2019, 125, 751–760. [Google Scholar] [CrossRef]
- Li, X.; Klaus, B. A novel carboxylesterase from Aspergillus niger and its hydrolysis of succinimide esters. Carlsberg. Res. Commun. 1989, 54, 241–249. [Google Scholar] [CrossRef]
- Toma, R.B.; Leung, H.K. Determination of Reducing Sugars in French Fried Potatoes by 3,5-Dinitrosalicylic Acid. Food Chem. 1987, 23, 29–33. [Google Scholar] [CrossRef]
- Zhao, G.; Nyman, M.; Jonsson, J.A. Rapid determination of short-chain fatty acids in colonic contents and faeces of humans and rats by acidified water-extraction and direct-injection gas chromatography. Biomed. Chromatogr. 2006, 20, 674–682. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Deng, Q.; Xu, J.; Wang, X.; Hu, C.; Tang, H.; Huang, F. Sinapic acid and resveratrol alleviate oxidative stress with modulation of gut microbiota in high-fat diet-fed rats. Food Res. Int. 2019, 116, 1202–1211. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Li, W.; Chen, Z.; Gao, X.; Yuan, G.; Pan, Y.; Chen, H. Effects of simulated gastrointestinal digestion in vitro on the chemical properties, antioxidant activity, alpha-amylase and alpha-glucosidase inhibitory activity of polysaccharides from Inonotus obliquus. Food Res. Int. 2018, 103, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Zhang, R.; Liu, Y.; Xiao, J.; Su, D.; Yi, Y.; Wang, G.; Wei, Z.; Zhang, M. Characterization and mesenteric lymph node cells-mediated immunomodulatory activity of litchi pulp polysaccharide fractions. Carbohydr. Polym. 2016, 152, 496–503. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Process | Time (h) | Content of Reducing Sugars (mg/mL) |
|---|---|---|
| Saliva digestion | 0 | 0.236 ± 0.009 a |
| 0.5 | 0.227 ± 0.005 a | |
| 1 | 0.234 ± 0.006 a | |
| 1.5 | 0.235 ± 0.008 a | |
| 2 | 0.233 ± 0.006 a | |
| Gastric juice digestion | 0 | 0.116 ± 0.011 a |
| 1 | 0.117 ± 0.012 a | |
| 2 | 0.115 ± 0.012 a | |
| 4 | 0.132 ± 0.014 a | |
| 6 | 0.118 ± 0.003 a | |
| Intestinal juice digestion | 0 | 0.532 ± 0.301 a |
| 1 | 0.545 ± 0.129 a | |
| 2 | 0.565 ± 0.203 a | |
| 4 | 0.54 ± 0.178 a | |
| 6 | 0.549 ± 0.219 a |
| Fermentation Time (h) | Monosaccharide (%) | ||||
|---|---|---|---|---|---|
| Rha | Fuc | Ara | Xyl | Gal | |
| 0 | 32.1 ± 0.018 a | 5.6 ± 0.004 a | 12.4 ± 0.007 a | 39.4 ± 0.015 a | 10.4 ± 0.001 a |
| 6 | 20.0 ± 0.009 b | 3.4 ± 0.002 b | 15.9 ± 0.005 b | 53.0 ± 0.029 b | 7.6 ± 0.003 b |
| 12 | 3.0 ± 0.002 c | 0.6 ± 0.001 c | 21.5 ± 0.010 c | 68.1 ± 0.031 c | 6.8 ± 0.005 c |
| 24 | 0 ± 0.002 d | 0 ± 0.031 d | 22.7 ± 0.009 d | 71.8 ± 0.042 d | 5.6 ± 0.004 d |
| Group | Time (h) | SCFAs (mmol/L) | ||||||
|---|---|---|---|---|---|---|---|---|
| Acetic Acid | Propionic Acid | i-Butyric Acid | n-Butyric Acid | i-Valeric Acid | n-Valeric Acid | Total | ||
| BLK | 0 | 0.275 ± 0.012 b | 1.578 ± 0.015 a | 0.00 ± 0.00 a | 1.503 ± 0.050 b | 0.210 ± 0.011 a | 0.00 ± 0.00 a | 3.568 ± 0.062 a |
| 6 | 0.904 ± 0.050 d | 2.054 ± 0.105 a | 0.164 ± 0.009 b | 1.689 ± 0.075 b | 0.254 ± 0.020 a | 0.297 ± 0.018 b | 5.362 ± 0.067 b | |
| 12 | 0.704 ± 0.055 c | 3.412 ± 0.086 b | 0.338 ± 0.062 c | 1.116 ± 0.075 a b | 0.592 ± 0.054 c | 0.886 ± 0.091 c | 7.051 ± 0.176 c | |
| 24 | 0.00 ± 0.00 a | 3.389 ± 0.093 b | 0.433 ± 0.030 d | 0.587 ± 0.041 a | 0.936 ± 0.112 d | 0.865 ± 0.111 c | 6.211 ± 0.103 bc | |
| INU | 0 | 0.275 ± 0.012 b | 1.578 ± 0.015 a | 0.00 ± 0.00 a | 1.503 ± 0.050 b | 0.210 ± 0.011 a | 0.00 ± 0.00 a | 3.568 ± 0.062 a |
| 6 | 2.027 ± 0.073 e | 10.093 ± 0.200 d | 0.00 ± 0.00 a | 7.051 ± 0.236 d | 0.208 ± 0.004 a | 0.323 ± 0.013 b | 19.702 ± 0.470 e | |
| 12 | 2.008 ± 0.040 e | 15.408 ± 0.510 f | 0.00 ± 0.00 a | 9.213 ± 0.352 e | 0.426 ± 0.011 b | 0.311 ± 0.018 b | 27.366 ± 0.895 f | |
| 24 | 4.150 ± 0.049 g | 15.029 ± 0.200 f | 0.00 ± 0.00 a | 12.271 ± 0.742 g | 0.631 ± 0.040 c | 0.386 ± 0.022 b | 32.467 ± 0.901 h | |
| FSP | 0 | 0.275 ± 0.012 b | 1.578 ± 0.015 a | 0.00 ± 0.00 a | 1.503 ± 0.050 b | 0.210 ± 0.011 a | 0.00 ± 0.00 a | 3.568 ± 0.062 a |
| 6 | 0.00 ± 0.00 a | 7.495 ± 0.526 c | 0.00 ± 0.00 a | 5.296 ± 0.318 c | 0.574 ± 0.061 c | 0.901 ± 0.074 c | 14.266 ± 0.881 d | |
| 12 | 2.569 ± 0.105 f | 14.053 ± 0.254 e | 0.00 ± 0.00 a | 10.484 ± 0.454 f | 1.508 ± 0.011 e | 2.111 ± 0.139 d | 30.723 ± 0.081 g | |
| 24 | 2.079 ± 0.088 e | 13.918 ± 0.475 e | 0.00 ± 0.00 a | 9.121 ± 0.236 e | 1.383 ± 0.033 e | 2.023 ± 0.177 d | 28.52 4 ± 0.081 f | |
| Taxon | Mean (%) | SEM (%) | ||||||
|---|---|---|---|---|---|---|---|---|
| Phylum | Family | Genus | BLK | FSP | INU | BLK | FSP | INU |
| Bacteroidetes | Prevotellaceae | Prevotella | 3.12 b | 60.39 a | 61.38 b | 0.39 | 1.44 | 3.89 |
| Porphyromonadaceae | Parabacteroides | 1.68 a | 0.46 b | 0.14 c | 0.09 | 0.05 | 0.03 | |
| Bacteroidaceae | Bactericides | 9.20 a | 4.46 b | 1.10 c | 0.67 | 1.27 | 0.23 | |
| Actinobacteria | Bifidobacteriaceae | Bifidobacterium | 2.24 b | 2.12 b | 15.83 a | 0.37 | 0.47 | 2.77 |
| Proteobacteria | Alcaligenaceae | Sutterella | 0.93 b | 1.69 a | 0.41 b | 0.11 | 0.41 | 0.04 |
| Firmicutes | Veillonellaceae | Phascolarctobacterium | 3.90 b | 8.76 a | 1.61 b | 0.67 | 2.29 | 0.36 |
| Clostridiaceae | Clostridium | 0.16 b | 4.86 a | 0.15 b | 0.01 | 1.23 | 0.03 | |
| Veillonellaceae | Megamonas | 2.13 b | 4.73 a | 3.42 b | 0.28 | 1.10 | 0.53 | |
| Ruminococcaceae | Faecalibacterium | 7.83 a | 1.76 c | 5.63 b | 0.65 | 0.31 | 1.25 | |
| Ruminococcaceae | Oscillospira | 2.04 a | 1.35 b | 0.44 c | 0.22 | 0.19 | 0.13 | |
| Lachnospiraceae | Ui_Lachnospiraceae | 1.59 a | 1.26 a | 0.59 b | 0.16 | 0.16 | 0.12 | |
| Ruminococcaceae | Ui_Ruminococcaceae | 3.91 a | 1.28 c | 2.89 b | 0.17 | 0.08 | 0.57 | |
| Veillonellaceae | Dialister | 0.15 c | 0.97 a | 0.55 b | 0.02 | 0.23 | 0.07 | |
| Lachnospiraceae | Blautia | 0.48 b | 0.77 a | 0.35 b | 0.06 | 0.10 | 0.08 | |
| Lactobacillaceae | Lactobacillus | 0.01 b | 0.78 b | 2.27 a | 0.00 | 0.10 | 0.85 | |
| Ui_Clostridiales | Ui_Clostridiales | 0.42 a | 0.45 a | 0.17 b | 0.06 | 0.08 | 0.07 | |
| Ruminococcaceae | Uc_Ruminococcaceae | 0.44 a | 0.35 a | 0.40 a | 0.01 | 0.05 | 0.08 | |
| Lachnospiraceae | Uc_Lachnospiraceae | 0.25 a | 0.41 a | 0.37 a | 0.03 | 0.10 | 0.06 | |
| Ruminococcaceae | Ruminococcus | 0.27 a | 0.31 a | 0.16 b | 0.04 | 0.03 | 0.03 | |
| Uc_Clostridiales | Uc_Clostridiales | 0.31 a | 0.32 a | 0.16 b | 0.03 | 0.07 | 0.01 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, X.; Zhang, Z.; Huang, F.; Yang, C.; Huang, Q. In Vitro Digestion and Fermentation by Human Fecal Microbiota of Polysaccharides from Flaxseed. Molecules 2020, 25, 4354. https://doi.org/10.3390/molecules25194354
Zhou X, Zhang Z, Huang F, Yang C, Huang Q. In Vitro Digestion and Fermentation by Human Fecal Microbiota of Polysaccharides from Flaxseed. Molecules. 2020; 25(19):4354. https://doi.org/10.3390/molecules25194354
Chicago/Turabian StyleZhou, Xin, Zhao Zhang, Fenghong Huang, Chen Yang, and Qingde Huang. 2020. "In Vitro Digestion and Fermentation by Human Fecal Microbiota of Polysaccharides from Flaxseed" Molecules 25, no. 19: 4354. https://doi.org/10.3390/molecules25194354
APA StyleZhou, X., Zhang, Z., Huang, F., Yang, C., & Huang, Q. (2020). In Vitro Digestion and Fermentation by Human Fecal Microbiota of Polysaccharides from Flaxseed. Molecules, 25(19), 4354. https://doi.org/10.3390/molecules25194354