The Effect of Chlorogenic Acid on Bacillus subtilis Based on Metabolomics

 and
and
Abstract
1. Introduction
2. Results and Discussion
2.1. MIC
2.2. Scanning Electron Microscope (SEM) and Membrane Permeability Assay
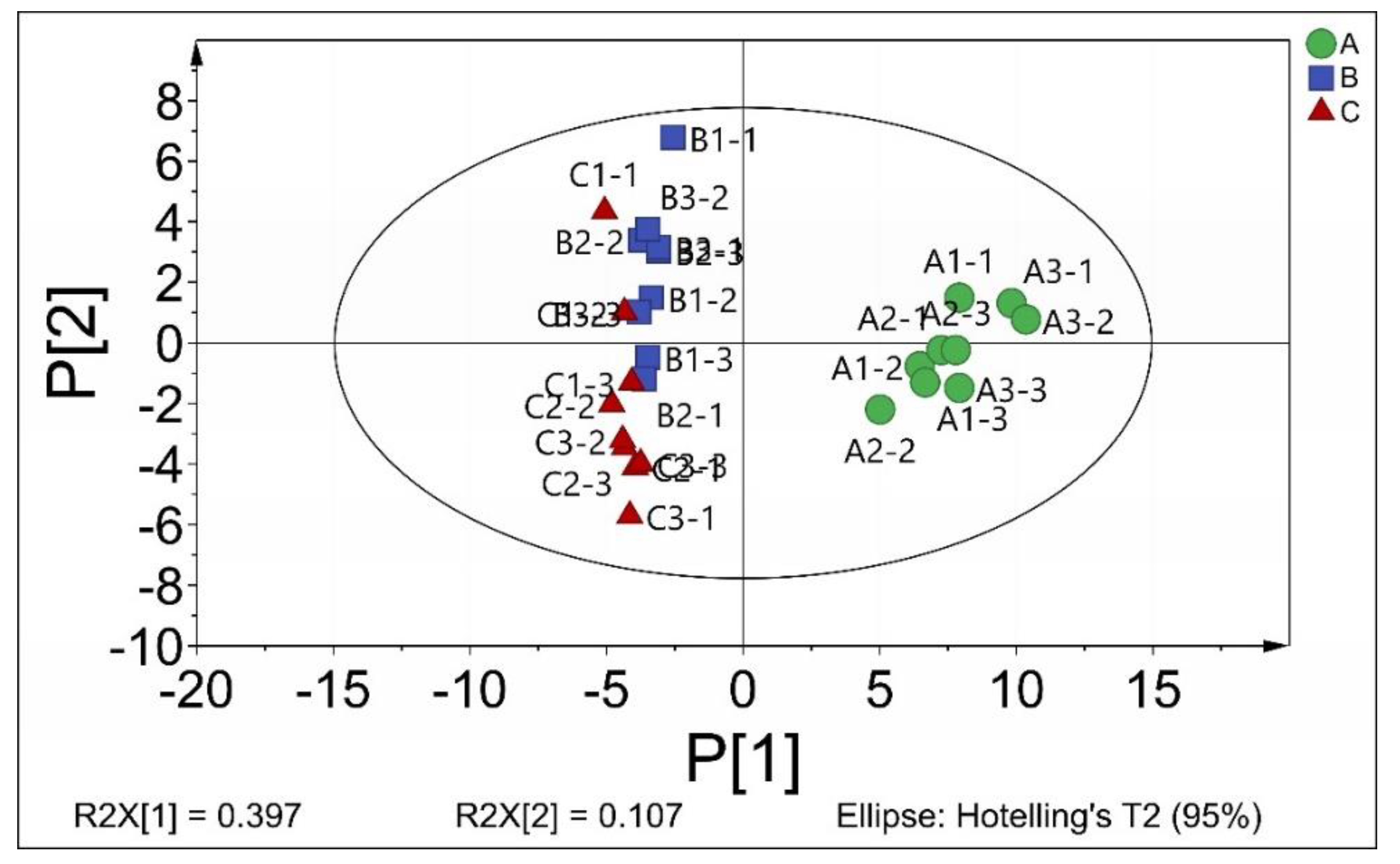
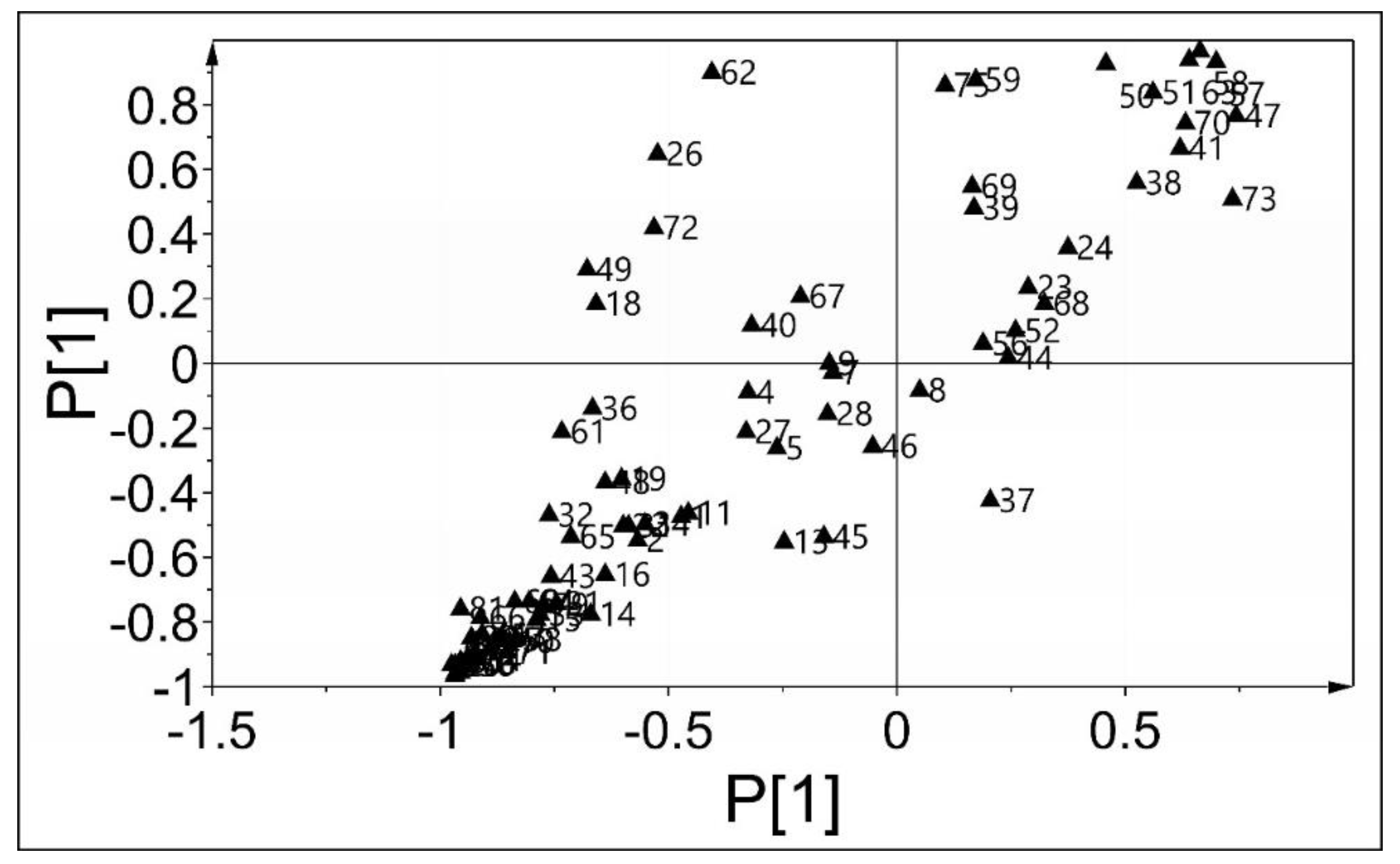
2.3. The Intracellular Metabolites of B. subtilis
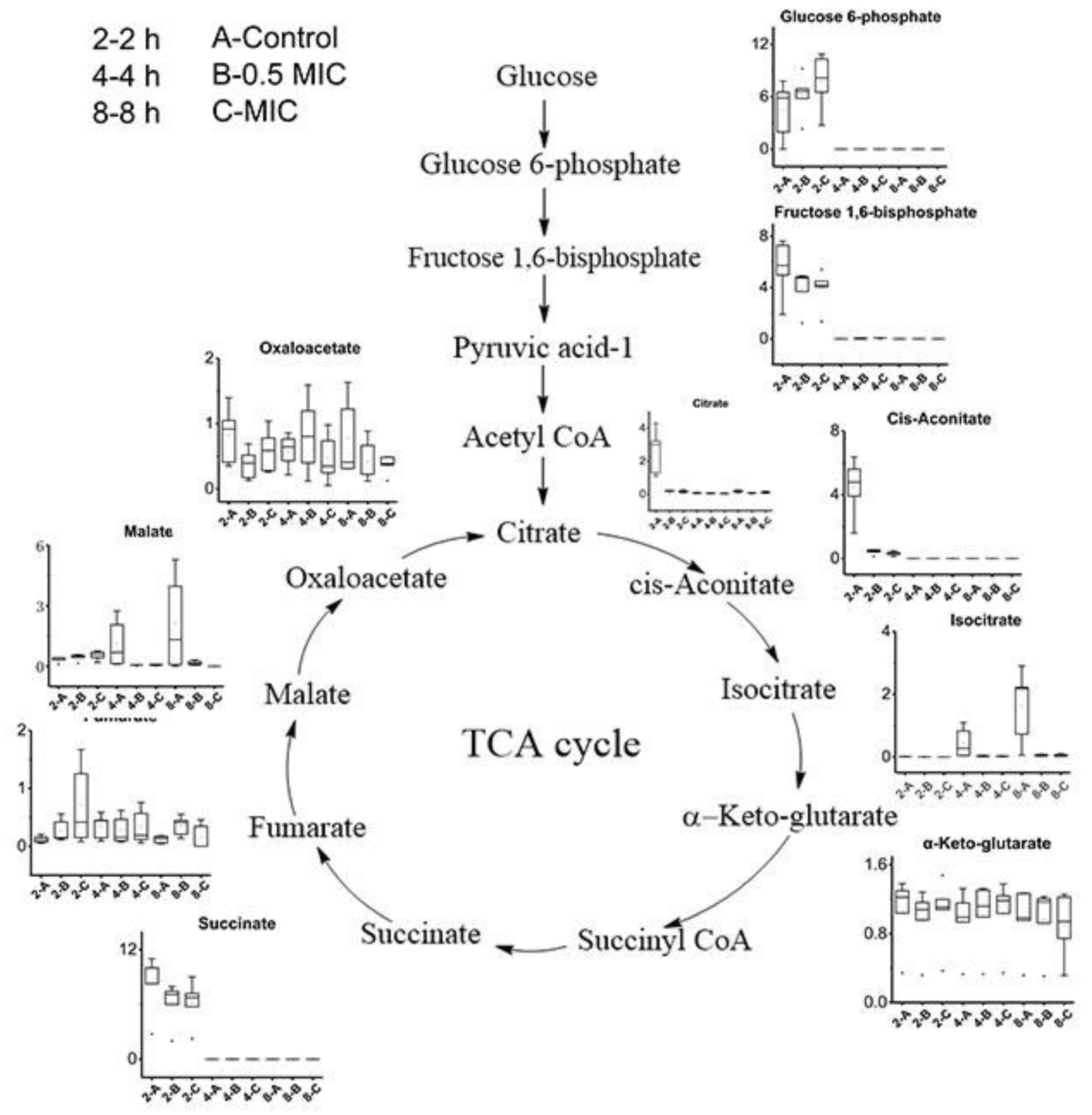
2.4. The Effect of CGA on the Primary Metabolism of B. subtilis
3. Materials and Methods
3.1. The Reagent and Bacterial Strains
3.2. Determination of MIC
3.3. The Detection of Extracellular Protein
3.4. SEM Assay
3.5. Membrane Permeability Assay
3.6. Measurement of the ATP Content
3.7. Extraction of the Intracellular Metabolites
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Santana-Gálvez, J.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Chlorogenic acid: Recent advances on its dual role as a food additive and a nutraceutical against metabolic syndrome. Molecules 2017, 22, 358. [Google Scholar] [CrossRef]
- Kabir, F.; Katayama, S.; Tanji, N.; Nakamura, S. Antimicrobial effects of chlorogenic acid and related compounds. J. Korean Soc. Appl. Biol. Chem. 2014, 57, 359–365. [Google Scholar] [CrossRef]
- Perrone, D.; Donangelo, R.; Donangelo, C.M.; Farah, A. Modeling weight loss and chlorogenic acids content in coffee during roasting. J. Agric. Food. Chem. 2010, 58, 12238–12243. [Google Scholar] [CrossRef]
- Awad, M.A.; de Jager, A.; van Westing, L.M. Flavonoid and chlorogenic acid levels in apple fruit: Characterisation of variation. Sci. Hortic. 2000, 83, 249–263. [Google Scholar] [CrossRef]
- Sung, W.S.; Lee, D.G. Antifungal action of chlorogenic acid against pathogenic fungi, mediated by membrane disruption. Pure Appl. Chem. 2010, 82, 219–226. [Google Scholar] [CrossRef]
- Pedro, M.d.M.; Isabelle, D.; Aurélie, H.; Aurélie, J.; Thomas, R.; Stéphanie, D.; Thérèse, S.; Schneider, Y.J. Anti-inflammatory effect and modulation of cytochrome P450 activities by Artemisia annua tea infusions in human intestinal Caco-2 cells. Food Chem. 2012, 134, 864–871. [Google Scholar]
- Wang, G.F.; Shi, L.P.; Ren, Y.D.; Liu, Q.F.; Liu, H.F.; Zhang, R.J.; Li, Z.; Zhu, F.H.; He, P.L.; Tang, W.; et al. Anti-hepatitis B virus activity of chlorogenic acid, quinic acid and caffeic acid in vivo and in vitro. Antiviral Res. 2009, 83, 186–190. [Google Scholar] [CrossRef]
- Bagdas, D.; Etoz, B.C.; Gul, Z.; Ziyanok, S.; Inan, S.; Turacozen, O.; Gul, N.Y.; Topal, A.; Cinkilic, N.; Tas, S.; et al. In vivo systemic chlorogenic acid therapy under diabetic conditions: Wound healing effects and cytotoxicity/genotoxicity profile. Food Chem. Toxicol. 2015, 81, 54–61. [Google Scholar] [CrossRef]
- Lou, Z.; Wang, H.; Lv, W.; Ma, C.; Wang, Z.; Chen, S. Assessment of antibacterial activity of fractions from burdock leaf against food-related bacteria. Food Control 2010, 21, 1272–1278. [Google Scholar] [CrossRef]
- Fattouch, S.; Caboni, P.; Coroneo, V.; Tuberoso, C.I.G.; Angioni, A.; Dessi, S.; Marzouki, N.; Cabras, P. Antimicrobial activity of tunisian quince (Cydonia oblonga Miller) pulp and peel polyphenolic extracts. J. Agric. Food. Chem. 2007, 55, 963–969. [Google Scholar] [CrossRef]
- Kim, B.G.; Jung, W.D.; Mok, H.; Ahn, J.H. Production of hydroxycinnamoyl-shikimates and chlorogenic acid in Escherichia coli: Production of hydroxycinnamic acid conjugates. Microb. Cell Fact. 2013, 12, 15. [Google Scholar] [CrossRef]
- Li, G.; Wang, X.; Xu, Y.; Zhang, B.; Xia, X. Antimicrobial effect and mode of action of chlorogenic acid on Staphylococcus aureus. Eur. Food Res. Technol. 2014, 238, 589–596. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, S.; Zhao, M.; Li, T. Study on extraction, purification and antimicrobial activity of tobacco chlorogenic acid. Modern Food Sci. Technol. 2008, 3, 233–236. [Google Scholar]
- Lou, Z.; Wang, H.; Zhu, S.; Ma, C.; Wang, Z. Antibacterial activity and mechanism of action of chlorogenic acid. J. Food Sci. 2011, 76, M398–M403. [Google Scholar] [CrossRef]
- Su, M.; Sun, Z.; Liu, F.; Wu, H.; Zhang, X.; Zhu, Y.; Daoying, W.; Xu, W.; Luo, Z. Antimicrobial mechanism of chlorogenic acid against chicken spoilage bacteria. Jiangsu J. Agric. Sci. 2018, 34, 1386–1391. [Google Scholar]
- Kono, Y.; Kobayashi, K.; Tagawa, S.; Adachi, K.; Ueda, A.; Sawa, Y.; Shibata, H. Antioxidant activity of polyphenolics in diets: Rate constants of reactions of chlorogenic acid and caffeic acid with reactive species of oxygen and nitrogen. BBA Gen. Subjects 1997, 1335, 335–342. [Google Scholar] [CrossRef]
- Francisco, V.; Costa, G.; Figueirinha, A.; Marques, C.; Pereira, P.; Miguel Neves, B.; Celeste Lopes, M.; García-Rodríguez, C.; Teresa Cruz, M.; Teresa Batista, M. Anti-inflammatory activity of cymbopogon citratus leaves infusion via proteasome and nuclear factor-κB pathway inhibition: Contribution of chlorogenic acid. J. Ethnopharmacol. 2013, 148, 126–134. [Google Scholar] [CrossRef]
- Ren, S.; Wu, M.; Guo, J.; Zhang, W.; Liu, X.; Sun, L.; Holyst, R.; Hou, S.; Fang, Y.; Feng, X. Sterilization of polydimethylsiloxane surface with Chinese herb extract: A new antibiotic mechanism of chlorogenic acid. Sci. Rep. 2015, 5, 10464. [Google Scholar] [CrossRef]
- Craig, H. Food-related illness and death in the United States. Emerg. Infect. Dis. 1999, 5, 840. [Google Scholar]
- Batt, C.A. Microbial food spoilage. In Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- André, S.; Vallaeys, T.; Planchon, S. Spore-forming bacteria responsible for food spoilage. Res. Microbiol. 2017, 168, 379–387. [Google Scholar] [CrossRef]
- Batt, C.A. Microbial Food Spoilage. 2016. Available online: https://doi.org/10.1016/B978-0-08-100596-5.03440-5 (accessed on 1 July 2016).
- Xu, Y.; Xu, X.; Xin, S. Separation and identification of spoilage microorganisms in instant wet noodles. China Brew 2014, 33, 68–71. [Google Scholar]
- Wang, X.M.; Chen, J.; Ying-Guo, L.V.; Liu, Z.L. Research progress of preservation technology for fresh noodles. Cereals Oils 2013, 26, 12–15. [Google Scholar]
- Fiehn, O. Metabolomics–the link between genotypes and phenotypes. Plant Mol. Biol. 2002, 48, 155–171. [Google Scholar] [CrossRef]
- Saghatelian, A.; Cravatt, B.F. Global strategies to integrate the proteome and metabolome. Curr. Opin. Chem. Biol. 2005, 9, 62–68. [Google Scholar] [CrossRef]
- Ding, M.Z.; Wang, X.; Yang, Y.; Yuan, Y.J. Metabolomic study of interactive effects of phenol, furfural, and acetic acid on saccharomyces cerevisiae. OMICS J. Integrative Biol. 2011, 15, 647–653. [Google Scholar] [CrossRef]
- Ravenzwaay, B.; Cunha, G.C.-P.; Leibold, E.; Looser, R.; Mellert, W.; Prokoudine, A.; Walk, T.; Wiemer, J. The use of metabolomics for the discovery of new biomarkers of effect. Toxicol. Lett. 2007, 172, 21–28. [Google Scholar] [CrossRef]
- Omelon, S.; Georgiou, J.; Habraken, W. A cautionary (spectral) tail: Red-shifted fluorescence by DAPI–DAPI interactions. Biochem. Soc. Trans. 2016, 44, 46–49. [Google Scholar] [CrossRef]
- Bo, T.; Liu, M.; Zhong, C.; Zhang, Q.; Su, Q.Z.; Tan, Z.L.; Han, P.P.; Jia, S.R. Metabolomic analysis of antimicrobial mechanisms of ε-Poly-l-lysine on saccharomyces cerevisiae. J. Agric. Food. Chem. 2014, 62, 4454–4465. [Google Scholar] [CrossRef]
- Bylesjö, M.; Rantalainen, M.; Cloarec, O.; Nicholson, J.K.; Holmes, E.; Trygg, J. OPLS discriminant analysis: Combining the strengths of PLS-DA and SIMCA classification. J. Chemom. 2006, 20, 341–351. [Google Scholar] [CrossRef]
- Halouska, S.; Fenton, R.J.; Barletta, R.G.; Powers, R. Predicting the in vivo mechanism of action for drug leads using NMR metabolomics. ACS Chem. Biol. 2012, 7, 166–171. [Google Scholar] [CrossRef]
- Zhu, Z.; Wang, H.; Shang, Q.; Jiang, Y.; Cao, Y.; Chai, Y. Time course analysis of Candida albicans metabolites during biofilm development. J. Proteome Res. 2013, 12, 2375–2385. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Huang, L.; Yang, Y.; Zhu, M.; Wen, Y.; Liu, Y.; Zhang, R.; Liu, J. Study on the mechanism of chlorogenic acid inhibiting Staphylococcu saureus. J. Southwest Univ. 2016, 38, 15–19. [Google Scholar]
- Barsch, A.; Carvalho, H.G.; Cullimore, J.V.; Niehaus, K. GC–MS based metabolite profiling implies three interdependent ways of ammonium assimilation in Medicago truncatula root nodules. J. Biotechnol. 2006, 127, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ma, M.; Luo, S.; Zhang, R.; Han, P.; Hu, W. Metabolic responses to ethanol in Saccharomyces cerevisiae using a gas chromatography tandem mass spectrometry-based metabolomics approach. Int. J. Biochem. Cell Biol. 2012, 44, 1087–1096. [Google Scholar] [CrossRef]
- Govaris, A.; Solomakos, N.; Pexara, A.; Chatzopoulou, P.S. The antimicrobial effect of oregano essential oil, nisin and their combination against Salmonella Enteritidis in minced sheep meat during refrigerated storage. Int. J. Food Microbiol. 2010, 137, 175–180. [Google Scholar] [CrossRef]
- Turgis, M.; Han, J.; Caillet, S.; Lacroix, M. Antimicrobial activity of mustard essential oil against Escherichia coli O157:H7 and Salmonella typhi. Food Control 2009, 20, 1073–1079. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NO. | Compound Name | NO. | Compound Name | NO. | Compound Name |
|---|---|---|---|---|---|
| 1 | Citric acid | 28 | L-Arginine | 55 | ADP |
| 2 | Cis-Aconitic acid/suberic acid | 29 | L-Aspartic acid | 56 | CDP |
| 3 | Isocitric acid | 30 | L-Glutamic acid | 57 | Cyclic AMP |
| 4 | Oxoglutaric acid | 31 | L-Glutamine | 58 | Cyclic GMP |
| 5 | Succinyl-CoA | 32 | L-Histidine | 59 | Cytidine triphosphate-1 |
| 6 | Succinic acid | 33 | L-Isoleucine | 60 | Guanine |
| 7 | Fumaric acid | 34 | Azelaic acid | 61 | Guanosine |
| 8 | Malic acid | 35 | D-2-Hydroxyglutarate | 62 | Guanosine diphosphate |
| 9 | Oxalacetic acid | 36 | Malonic acid | 63 | Guanosine monophosphate-2 |
| 10 | Glucose 6-phosphate | 37 | Glutaric acid | 64 | Guanosine triphosphate |
| 11 | Fructose 6-phosphate | 38 | Salicylic acid | 65 | Hypoxanthine |
| 12 | Fructose 1,6-bisphosphate | 39 | Phenylacetic acid | 66 | IDP |
| 13 | Dihydroxyacetone phosphate | 40 | Glucose 1-phosphate | 67 | Inosine |
| 14 | 3-Phosphoglycerate | 41 | Deoxyuridine triphosphate | 68 | Inosinic acid(IMP) |
| 15 | Phosphoenolpyruvic acid | 42 | Glycerol 3-phosphate | 69 | Pimelic acid/2-oxoadipate |
| 16 | Pyruvic acid-1 | 43 | Sedoheptulose 1,7-bisphosphate | 70 | Putrescine |
| 17 | L-Lactic acid | 44 | Sedoheptulose 7-phosphate | 71 | Pyridoxal phosphate |
| 18 | L-Leucine | 45 | Ribose-5-phosphate | 72 | Tryptamine |
| 19 | L-Lysine | 46 | Fructose 1-phosphate | 73 | Tyramine |
| 20 | L-Methionine | 47 | Orotic acid | 74 | Uracil |
| 21 | L-Phenylalanine | 48 | Oxalic acid | 75 | Uridine 5′-diphosphate |
| 22 | L-Proline | 49 | Pantothenic Acid | 76 | Uridine diphosphate glucose |
| 23 | L-Serine | 50 | Nicotinic acid-1 | 77 | Uridine diphosphate glucuronic acid |
| 24 | L-Threonine | 51 | 4-Hydroxybenzaldehyde | 78 | Uridine triphosphate |
| 25 | L-Tyrosine | 52 | 4-Hydroxybenzoic acid | 79 | Xanthine |
| 26 | L-Valine | 53 | Adenosine diphosphate ribose | 80 | Xanthosine |
| 27 | L-Alanine | 54 | Adenosine triphosphate | 81 | Xanthylic acid |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Liang, S.; Zhang, M.; Wang, Z.; Wang, Z.; Ren, X. The Effect of Chlorogenic Acid on Bacillus subtilis Based on Metabolomics. Molecules 2020, 25, 4038. https://doi.org/10.3390/molecules25184038
Wu Y, Liang S, Zhang M, Wang Z, Wang Z, Ren X. The Effect of Chlorogenic Acid on Bacillus subtilis Based on Metabolomics. Molecules. 2020; 25(18):4038. https://doi.org/10.3390/molecules25184038
Chicago/Turabian StyleWu, Yan, Shan Liang, Min Zhang, Zhenhua Wang, Ziyuan Wang, and Xin Ren. 2020. "The Effect of Chlorogenic Acid on Bacillus subtilis Based on Metabolomics" Molecules 25, no. 18: 4038. https://doi.org/10.3390/molecules25184038
APA StyleWu, Y., Liang, S., Zhang, M., Wang, Z., Wang, Z., & Ren, X. (2020). The Effect of Chlorogenic Acid on Bacillus subtilis Based on Metabolomics. Molecules, 25(18), 4038. https://doi.org/10.3390/molecules25184038