Molecular Identification, Characterization, and Expression Analysis of a Gonadotropin-Releasing Hormone Receptor (GnRH-R) in Pacific Abalone, Haliotis discus hannai



Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
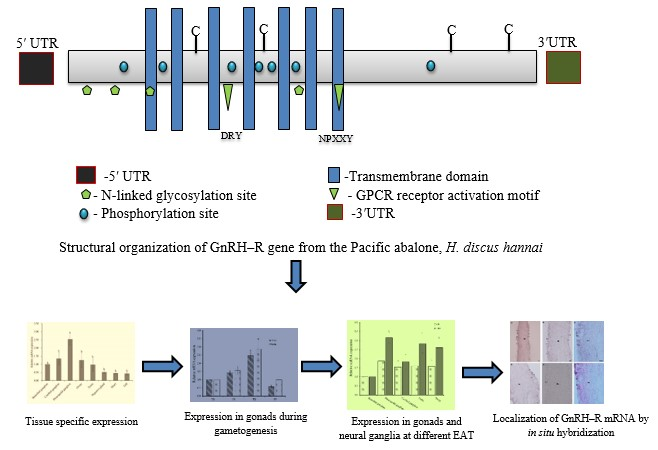
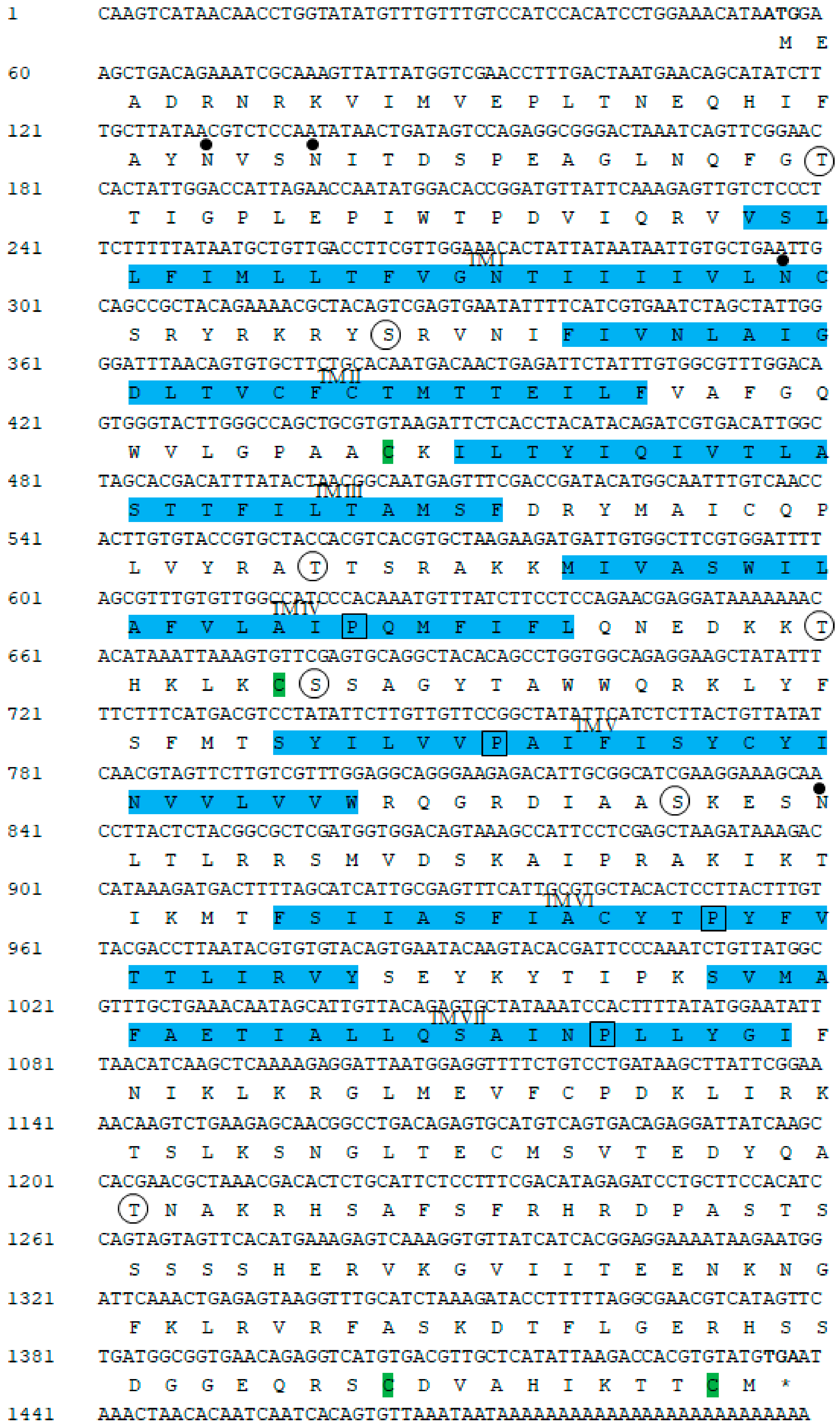
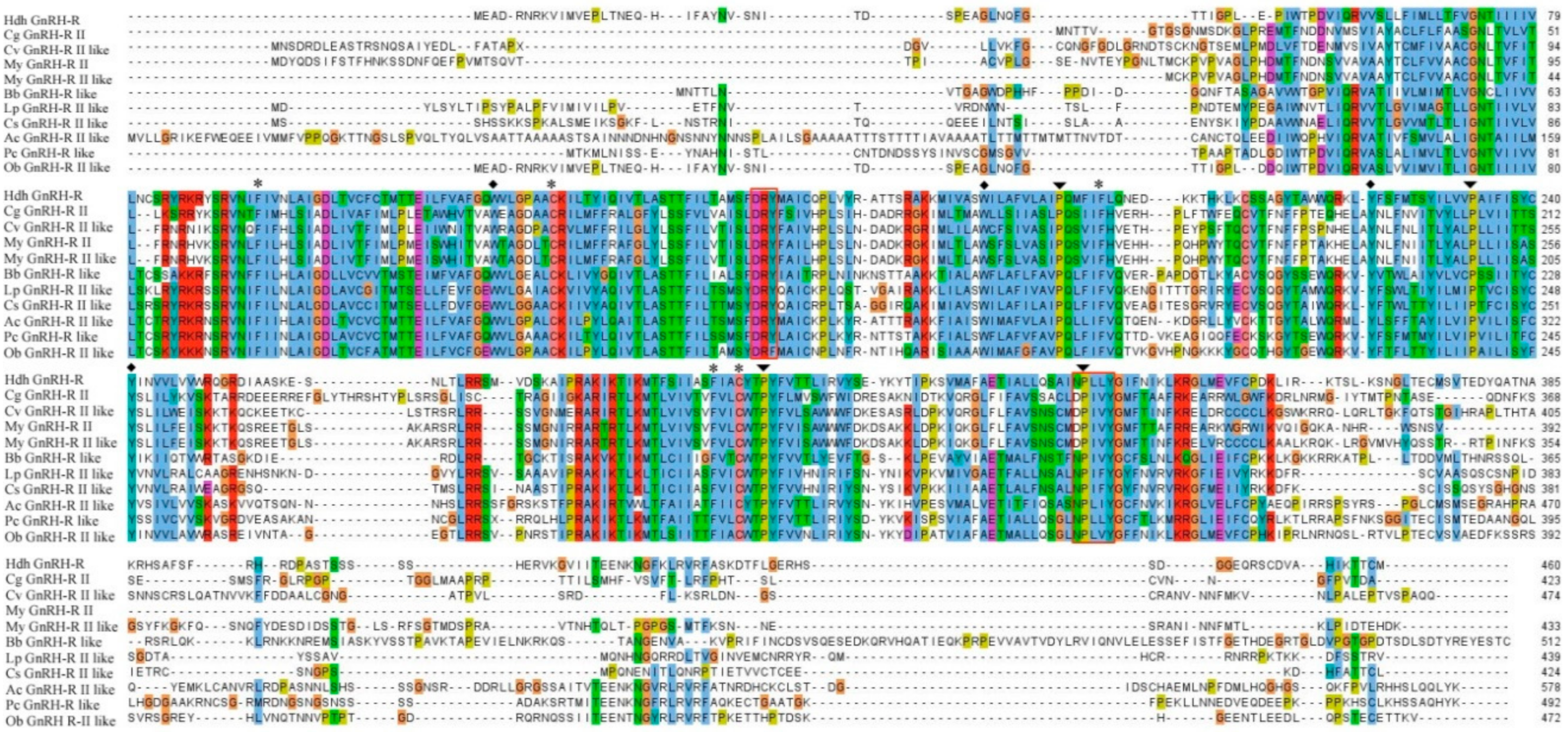
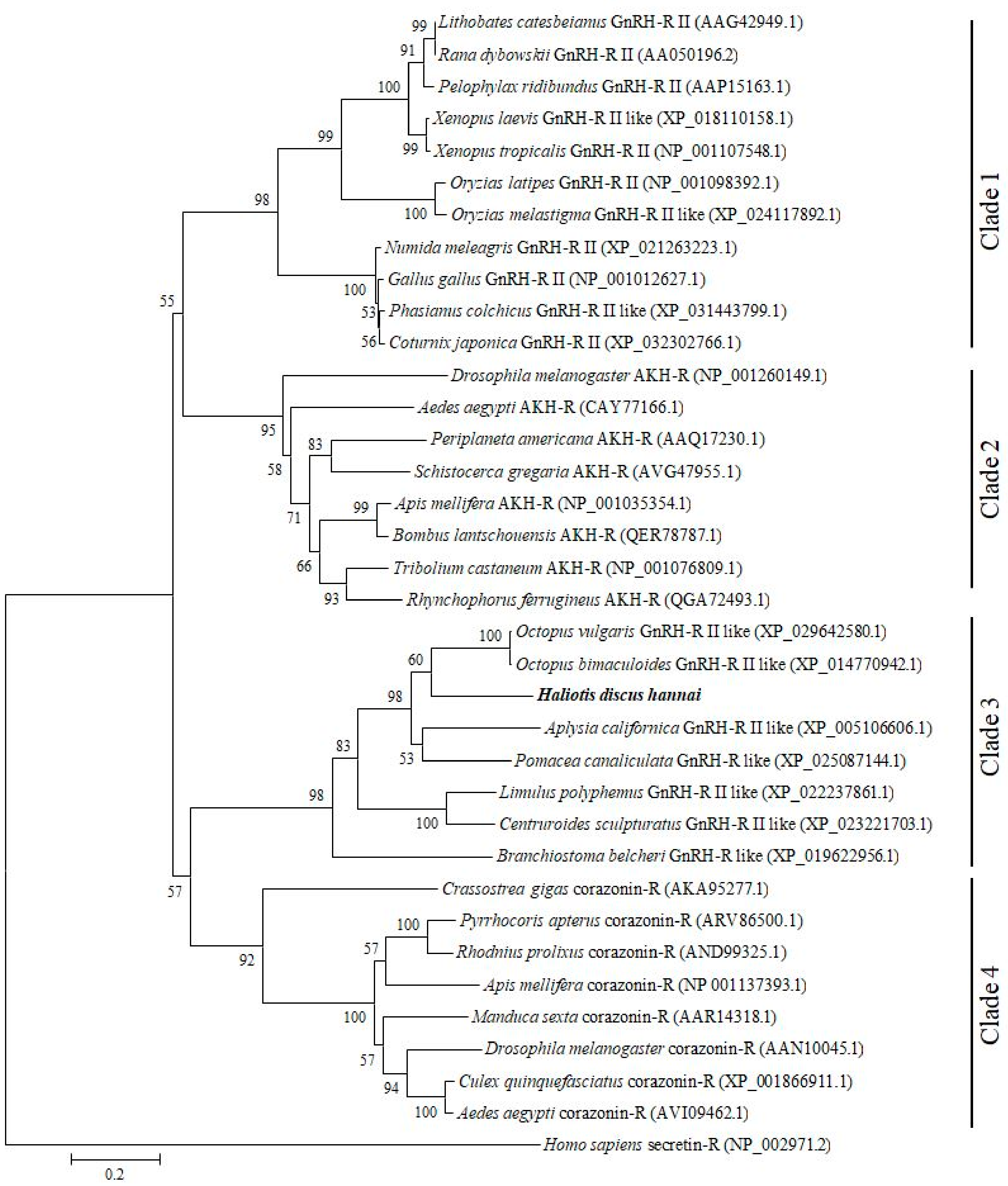
2.1. Cloning and Sequence Analysis of GnRH-R Gene
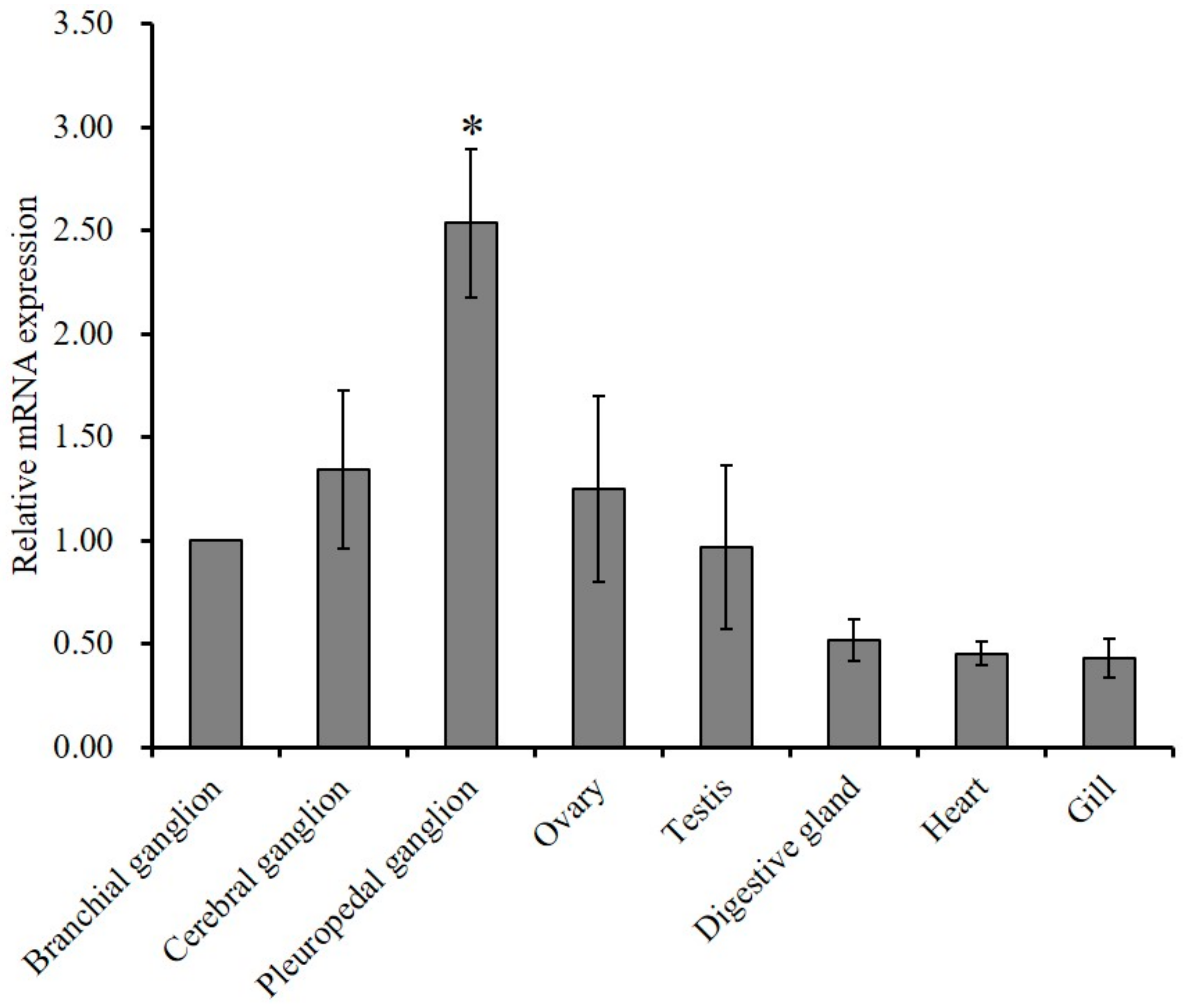
2.2. Tissue Expression Profile of GnRH-R mRNA
3. Discussion
4. Materials and Methods
4.1. Experimental Animals and Tissue Collection
4.2. RNA Isolation and cDNA Synthesis
4.3. cDNA Cloning of GnRH-R Gene
4.4. Bioinformatics Analysis
4.5. Quantitative PCR Expression Analysis
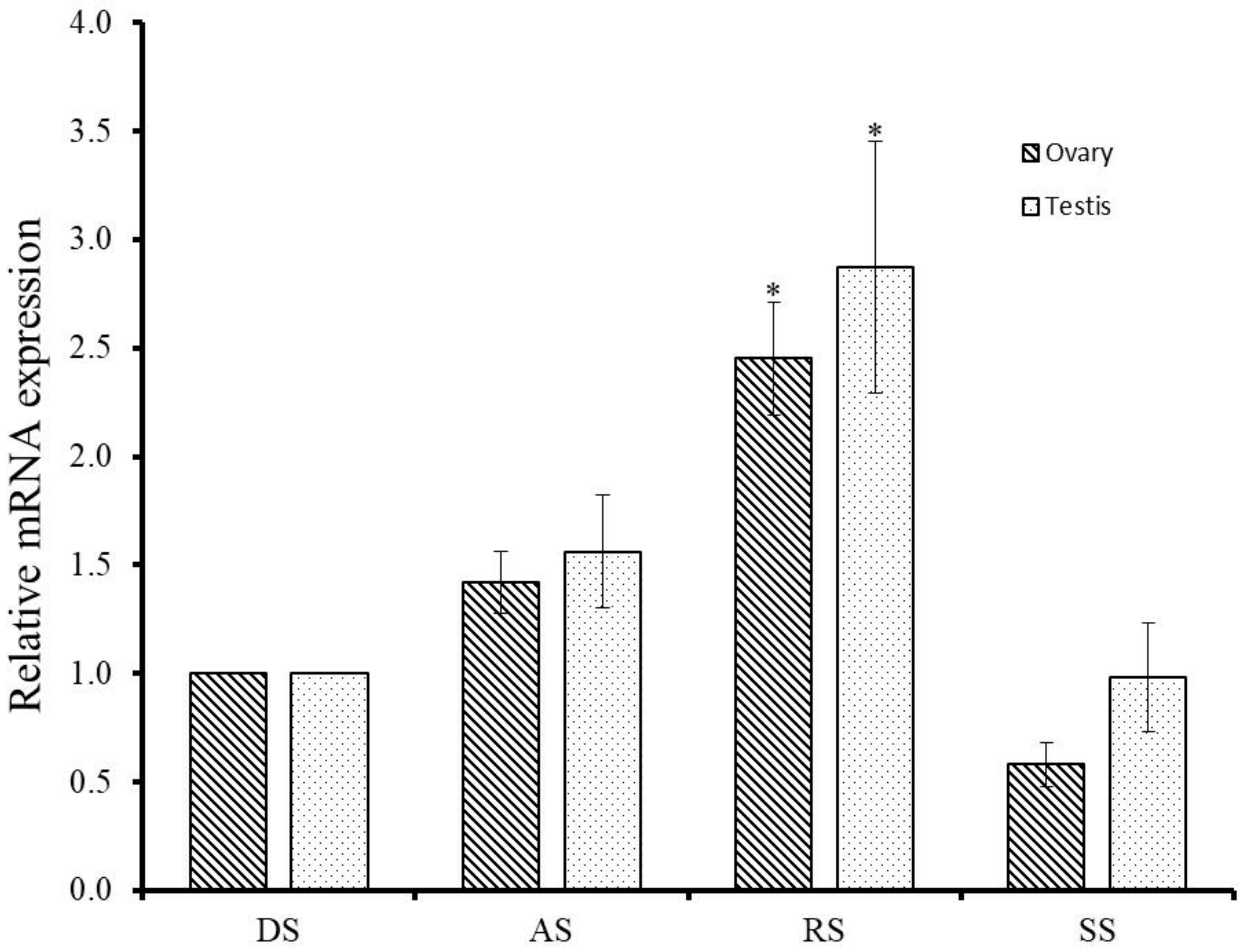
4.6. Gonadal Expression of GnRH-R mRNA during Gametogenesis
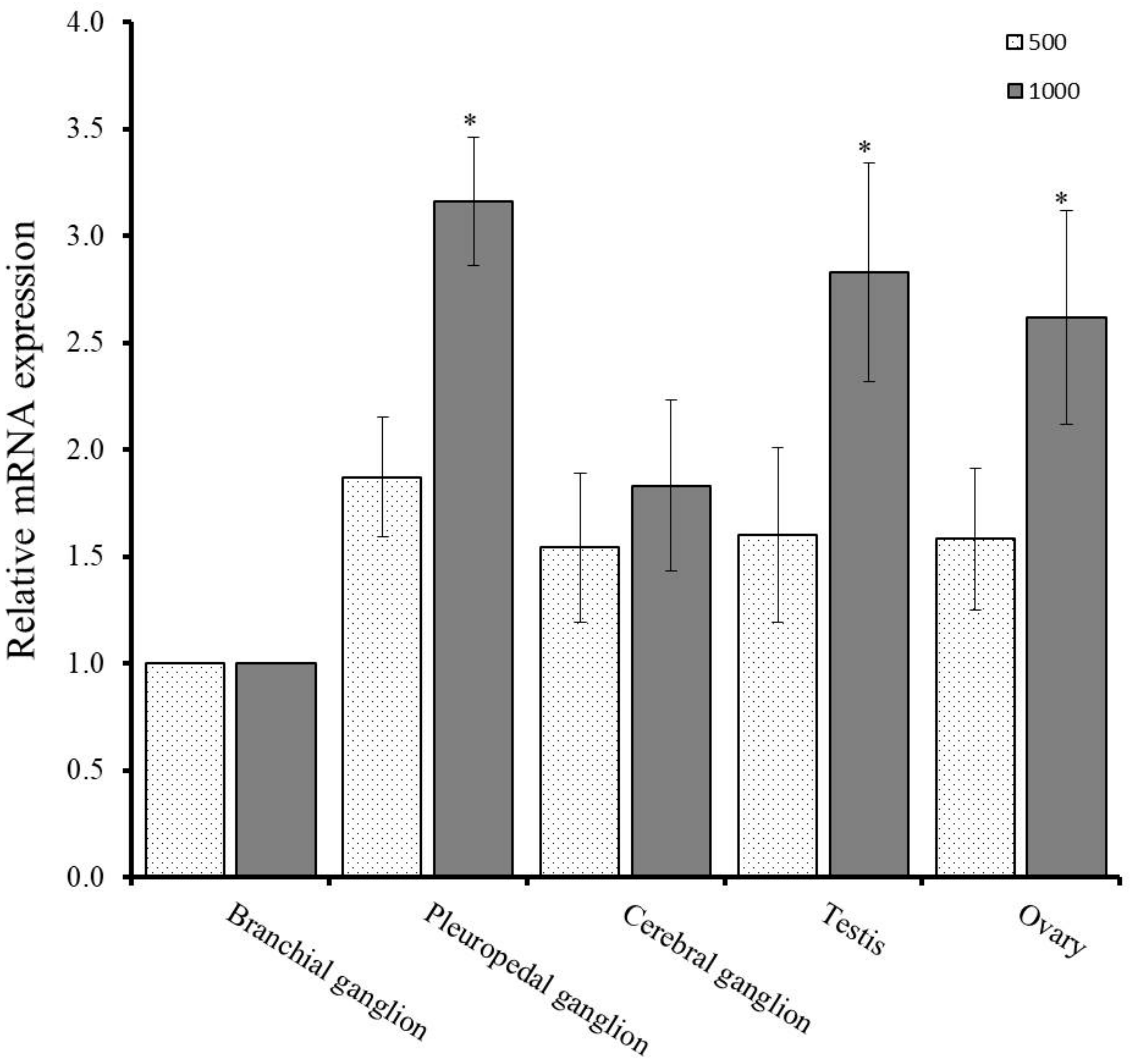
4.7. Effects of Effective Accumulated Temperature (EAT) on GnRH-R mRNA in Neural Ganglia and Gonad
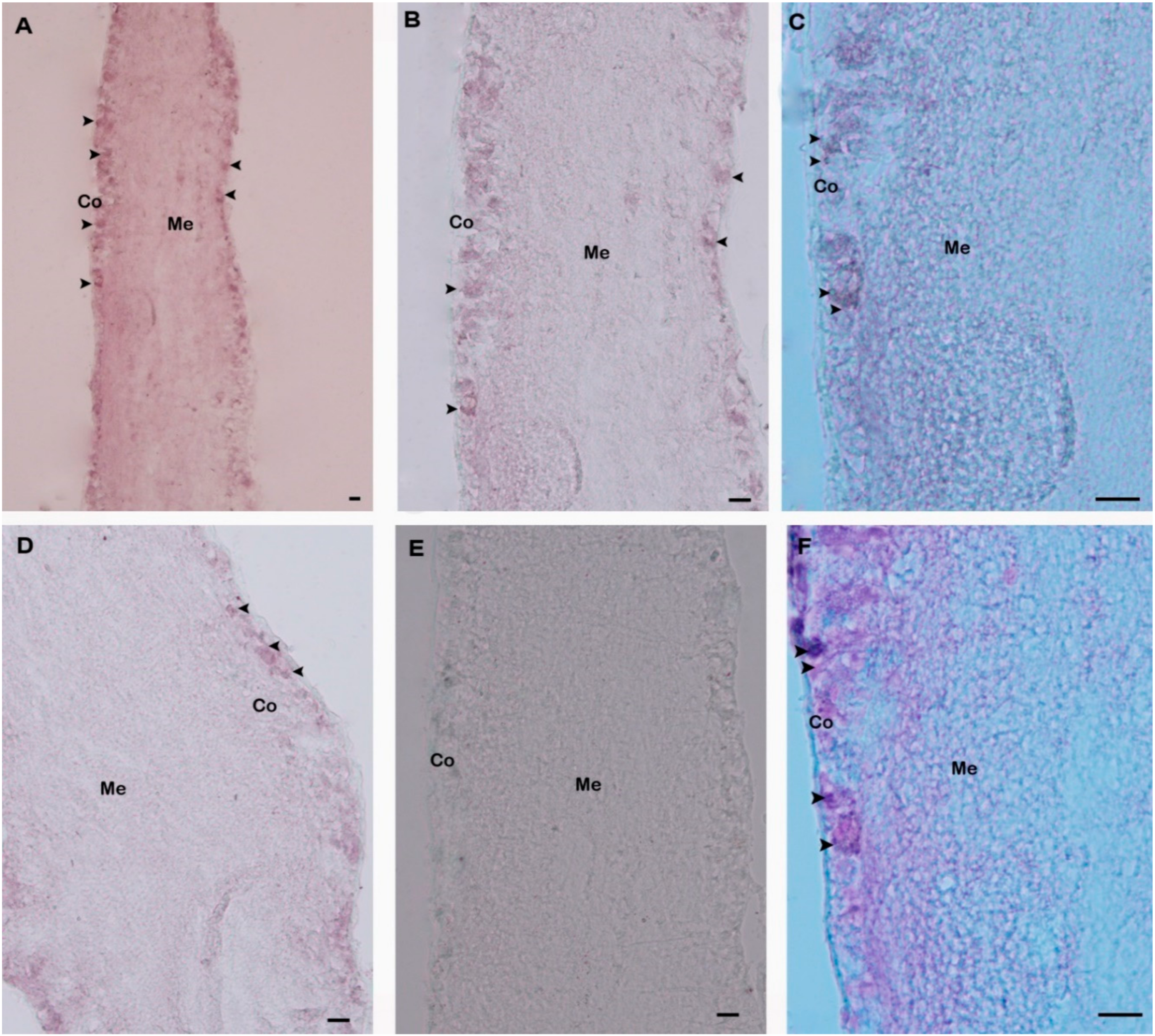
4.8. In Situ Hybridization (ISH)
4.9. Nuclear Fast Red Counterstain
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Flanagan, C.A.; Manilall, A. Gonadotropin-releasing hormone (GnRH) receptor structure and GnRH binding. Front. Endocrinol. 2017, 8, 1–14. [Google Scholar]
- Sealfon, S.C.; Millar, R.P. Functional domains of the gonadotropin-releasing hormone receptor. Cell. Mol. Neurobiol. 1995, 15, 25–42. [Google Scholar] [CrossRef] [PubMed]
- Munk, C.; Isberg, V.; Mordalski, S.; Harpsøe, K.; Rataj, K.; Hauser, A.S.; Kolb, P.; Bojarski, A.J.; Vriend, G.; Gloriam, D.E. GPCRdb: The G protein-coupled receptor database-an introduction. Br. J. Pharmacol. 2016, 173, 2195–2207. [Google Scholar] [CrossRef] [PubMed]
- Perrett, R.M.; McArdle, C.A. Molecular mechanisms of gonadotropin-releasing hormone signaling: Integrating cyclic nucleotides into the network. Front. Endocrinol. 2013, 4, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Naor, Z. Signaling by G-protein-coupled receptor (GPCR): Studies on the GnRH receptor. Front. Neuroendocrinol. 2009, 30, 10–29. [Google Scholar] [CrossRef]
- Roch, G.J.; Busby, E.R.; Sherwood, N.M. GnRH receptors and peptides: Skating backward. Gen. Comp. Endocrinol. 2014, 209, 118–134. [Google Scholar] [CrossRef] [PubMed]
- Grimmelikhuijzen, C.J.P.; Hauser, F. Mini-review: The evolution of neuropeptide signaling. Regul. Pept. 2012, 177, S6. [Google Scholar] [CrossRef]
- Roch, G.J.; Busby, E.R.; Sherwood, N.M. Evolution of GnRH: Diving deeper. Gen. Comp. Endocrinol. 2011, 171, 1–16. [Google Scholar] [CrossRef]
- Tsutsumi, M.; Zhou, W.; Millar, R.P.; Mellon, P.L.; Roberts, J.L.; Flanagan, C.A.; Dong, K.; Gillo, B.; Sealfon, S.C. Cloning and functional expression of a mouse gonadotropin-releasing hormone receptor. Mol. Endocrinol. 1992, 6, 1163–1169. [Google Scholar]
- Bogerd, J.; Diepenbroek, W.B.; Hund, E.; van Oosterhout, F.; Teves, A.C.C.; Leurs, R.; Blomenröhr, M. Two gonadotropin-releasing hormone receptors in the African catfish: No differences in ligand selectivity, but differences in tissue distribution. Endocrinology 2002, 143, 4673–4682. [Google Scholar] [CrossRef]
- Illing, N.; Troskie, B.E.; Nahorniak, C.S.; Hapgood, J.P.; Peter, R.E.; Millar, R.P. Two gonadotropin-releasing hormone receptor subtypes with distinct ligand selectivity and differential distribution in brain and pituitary in the goldfish (Carassius auratus). Proc. Natl. Acad. Sci. USA 1999, 96, 2526–2531. [Google Scholar] [CrossRef] [PubMed]
- Millar, R.; Lowe, S.; Conklin, D.; Pawson, A.; Maudsley, S.; Troskie, B.; Ott, T.; Millar, M.; Lincoln, G.; Sellar, R.; et al. A novel mammalian receptor for the evolutionarily conserved type II GnRH. Proc. Natl. Acad. Sci. USA 2001, 98, 9636–9641. [Google Scholar] [CrossRef] [PubMed]
- Neill, J.D.; Duck, L.W.; Sellers, J.C.; Musgrove, L.C. A gonadotropin-releasing hormone (GnRH) receptor specific for GnRH II in primates. Biochem. Biophys. Res. Commun. 2001, 282, 1012–1018. [Google Scholar] [CrossRef] [PubMed]
- Okubo, K.; Nagata, S.; Ko, R.; Kataoka, H.; Yoshiura, Y.; Mitani, H.; Kondo, M.; Naruse, K.; Shima, A.; Aida, K. Identification and characterization of two distinct GnRH receptor subtypes in a teleost, the medaka Oryzias latipes. Endocrinology 2001, 142, 4729–4739. [Google Scholar] [CrossRef] [PubMed]
- Seong, J.Y.; Wang, L.; Oh, D.Y.; Yun, O.; Maiti, K.; Li, J.H.; Soh, J.M.; Choi, H.S.; Kim, K.; Vaudry, H.; et al. Ala/Thr(201) in extracellular loop 2 and Leu/Phe(290) in transmembrane domain 6 of type 1 frog gonadotropin-releasing hormone receptor confer differential ligand sensitivity and signal transduction. Endocrinology 2003, 144, 454–466. [Google Scholar] [CrossRef] [PubMed]
- González-Martínez, D.; Madigou, T.; Mañanos, E.; Cerdá-Reverter, J.M.; Zanuy, S.; Kah, O.; Muñoz-Cueto, J.A. Cloning and expression of gonadotropin-releasing hormone receptor in the brain and pituitary of the European sea bass: An in situ hybridization study. Biol. Reprod. 2004, 70, 1380–1391. [Google Scholar] [CrossRef] [PubMed]
- King, J.A.; Fidler, A.; Lawrence, S.; Adam, T.; Millar, R.P.; Katz, A. Cloning and expression, pharmacological characterization, and internalization kinetics of the pituitary GnRH receptor in a metatherian species of mammal. Gen. Comp. Endocrinol. 2000, 117, 439–448. [Google Scholar] [CrossRef]
- Cui, J.; Smith, R.G.; Mount, G.R.; Lo, J.L.; Yu, J.; Walsh, T.F.; Singh, S.B.; DeVita, R.J.; Goulet, M.T.; Schaeffer, J.M.; et al. Identification of Phe313 of the gonadotropin-releasing hormone (GnRH) receptor as a site critical for the binding of nonpeptide GnRH antagonists. Mol. Endocrinol. 2000, 14, 671–681. [Google Scholar] [CrossRef][Green Version]
- Troskie, B.E.; Hapgood, J.P.; Millar, R.P.; Illing, N. Complementary deoxyribonucleic acid cloning, gene expression, and ligand selectivity of a novel gonadotropin-releasing hormone receptor expressed in the pituitary and midbrain of Xenopus laevis. Endocrinology 2000, 141, 1764–1771. [Google Scholar] [CrossRef]
- Kanda, A.; Takahashi, T.; Satake, H.; Minakata, H. Molecular and functional characterization of a novel gonadotropin-releasing-hormone receptor isolated from the common octopus (Octopus vulgaris). Biochem. J. 2006, 395, 125–135. [Google Scholar] [CrossRef]
- Nagasawa, K.; Muroi, K.; Thitiphuree, T.; Minegishi, Y.; Itoh, N.; Osada, M. Cloning of invertebrate gonadotropin-releasing hormone receptor (GnRHR)-like gene in Yesso scallop, Patinopecten yessoensis. Agri Gene 2017, 3, 46–56. [Google Scholar] [CrossRef]
- Rodet, F.; Lelong, C.; Dubos, M.P.; Costil, K.; Favrel, P. Molecular cloning of a molluscan gonadotropin-releasing hormone receptor orthologue specifically expressed in the gonad. Biochim. Biophys. Acta Gene Struct. Expr. 2005, 1730, 187–195. [Google Scholar] [CrossRef]
- Suleria, H.A.R.; Masci, P.P.; Gobe, G.C.; Osborne, S.A. Therapeutic potential of abalone and status of bioactive molecules: A comprehensive review. Crit. Rev. Food Sci. Nutr. 2017, 57, 1742–1748. [Google Scholar] [CrossRef] [PubMed]
- Rodet, F.; Lelong, C.; Dubos, M.-P.; Favrel, P. Alternative splicing of a single precursor mRNA generates two subtypes of Gonadotropin-Releasing Hormone receptor orthologues and their variants in the bivalve mollusc Crassostrea gigas. Gene 2008, 414, 1–9. [Google Scholar] [CrossRef]
- Blom, N.; Gammeltoft, S.; Brunak, S. Sequence and structure-based prediction of eukaryotic protein phosphorylation sites. J. Mol. Biol. 1999, 294, 1351–1362. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, J.M. The probable arrangement of the helices in G protein-coupled receptors. EMBO J. 1993, 12, 1693–1703. [Google Scholar] [CrossRef]
- Bockaert, J.; Pin, J.P. Molecular tinkering of G protein-coupled receptors: An evolutionary success. EMBO J. 1999, 18, 1723–1729. [Google Scholar] [CrossRef]
- Ali, E.S.; Hua, J.; Wilson, C.H.; Tallis, G.A.; Zhou, F.H.; Rychkov, G.Y.; Barritt, G.J. The glucagon-like peptide-1 analogue exendin-4 reverses impaired intracellular Ca2 + signalling in steatotic hepatocytes. Biochim. Biophys. Acta Mol. Cell Res. 2016, 1863, 2135–2146. [Google Scholar] [CrossRef] [PubMed]
- Findlay, J.; Eliopoulos, E. Three-dimensional modelling of G protein-linked receptors. Trends Pharmacol. Sci. 1990, 11, 492–499. [Google Scholar] [CrossRef]
- O’Dowd, B.F.; Hnatowich, M.; Caron, M.G.; Lefkowitz, R.J.; Bouvier, M. Palmitoylation of the human β2-adrenergic receptor. J. Biol. Chem. 1989, 264, 7564–7569. [Google Scholar] [PubMed]
- Oliveira, L.; Paiva, A.C.; Sander, C.; Vriend, G. A common step for signal transduction in G protein-coupled receptors. Trends Pharmacol. Sci. 1994, 15, 170–172. [Google Scholar] [CrossRef]
- Scheer, A.; Fanelli, F.; Costa, T.; De Benedetti, P.G.; Cotecchia, S. Constitutively active mutants of the alpha 1B-adrenergic receptor: Role of highly conserved polar amino acids in receptor activation. EMBO J. 1996, 15, 3566–3578. [Google Scholar] [CrossRef] [PubMed]
- Probst, W.C.; Snyder, L.A.; Schuster, D.I.; Brosius, J.; Sealfon, S.C. Sequence Alignment of the G-Protein Coupled Receptor Superfamily. DNA Cell Biol. 1992, 11, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Gether, U. Uncovering molecular mechanisms involved in activation of G protein-coupled receptors. Endocr. Rev. 2000, 21, 90–113. [Google Scholar] [CrossRef] [PubMed]
- Hibert, M.F.; Trumpp-Kallmeyer, S.; Bruinvels, A.; Hoflack, J. Three-dimensional models of neurotransmitter G-binding protein-coupled receptors. Mol. Pharmacol. 1991, 40, 8–15. [Google Scholar]
- Hall, J.A.; Decatur, W.A.; Daukss, D.M.; Hayes, M.K.; Marquis, T.J.; Morin, S.J.; Kelleher, T.F.; Sower, S.A. Expression of three GnRH receptors in specific tissues in male and female sea lampreys Petromyzon marinus at three distinct life stages. Front. Neurosci. 2013, 7, 1–9. [Google Scholar] [CrossRef]
- Uki, N.; Kikuchi, S. Regulation of maturation and spawning of an abalone, Haliotis (Gastropoda) by external environmental factors. Aquaculture 1984, 39, 247–261. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 1–6. [Google Scholar] [CrossRef]
- Alva, V.; Nam, S.-Z.; Söding, J.; Lupas, A.N. The MPI bioinformatics Toolkit as an integrative platform for advanced protein sequence and structure analysis. Nucleic Acids Res. 2016, 44, W410–W415. [Google Scholar] [CrossRef]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2-A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Wan, Q.; Whang, I.; Choi, C.Y.; Lee, J.S.; Lee, J. Validation of housekeeping genes as internal controls for studying biomarkers of endocrine-disrupting chemicals in disk abalone by real-time PCR. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2011, 153, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Sharker, M.R.; Sukhan, Z.P.; Kim, S.C.; Lee, W.K.; Kho, K.H. Identification, characterization, and expression analysis of a serotonin receptor involved in the reproductive process of the Pacific abalone, Haliotis discus hannai. Mol. Biol. Rep. 2020, 47, 555–567. [Google Scholar] [CrossRef]
- Sharker, M.R.; Kim, S.C.; Sumi, K.R.; Sukhan, Z.P.; Sohn, Y.C.; Lee, W.K.; Kho, K.H. Characterization and expression analysis of a GnRH-like peptide in the Pacific abalone, Haliotis discus hannai. Agri Gene 2020, 15, 1–10. [Google Scholar] [CrossRef]
- Sharker, M.R.; Nou, I.-S.; Kho, K.H. Molecular characterization and spatiotemporal expression of prohormone convertase 2 in the Pacific abalone, Haliotis discus hannai. PLoS ONE 2020, 15, e0231353. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds described in the paper are available from the authors. |







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharker, M.R.; Sukhan, Z.P.; Kim, S.C.; Lee, W.K.; Kho, K.H. Molecular Identification, Characterization, and Expression Analysis of a Gonadotropin-Releasing Hormone Receptor (GnRH-R) in Pacific Abalone, Haliotis discus hannai. Molecules 2020, 25, 2733. https://doi.org/10.3390/molecules25122733
Sharker MR, Sukhan ZP, Kim SC, Lee WK, Kho KH. Molecular Identification, Characterization, and Expression Analysis of a Gonadotropin-Releasing Hormone Receptor (GnRH-R) in Pacific Abalone, Haliotis discus hannai. Molecules. 2020; 25(12):2733. https://doi.org/10.3390/molecules25122733
Chicago/Turabian StyleSharker, Md. Rajib, Zahid Parvez Sukhan, Soo Cheol Kim, Won Kyo Lee, and Kang Hee Kho. 2020. "Molecular Identification, Characterization, and Expression Analysis of a Gonadotropin-Releasing Hormone Receptor (GnRH-R) in Pacific Abalone, Haliotis discus hannai" Molecules 25, no. 12: 2733. https://doi.org/10.3390/molecules25122733
APA StyleSharker, M. R., Sukhan, Z. P., Kim, S. C., Lee, W. K., & Kho, K. H. (2020). Molecular Identification, Characterization, and Expression Analysis of a Gonadotropin-Releasing Hormone Receptor (GnRH-R) in Pacific Abalone, Haliotis discus hannai. Molecules, 25(12), 2733. https://doi.org/10.3390/molecules25122733