Immobilization of Purified Pectin Lyase from Pseudomonas putida onto Magnetic Lily Flowers (Lilium candidum L.) Nanoparticles and Applicability in Industrial Processes


Abstract
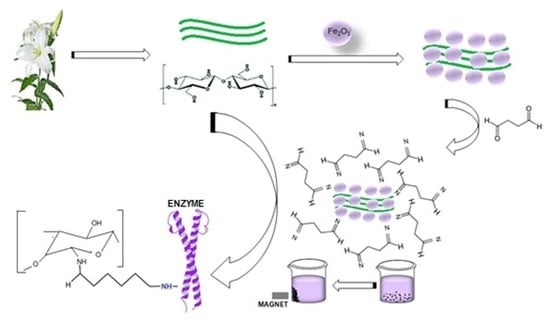
1. Introduction
2. Results and Discussion
2.1. Purification of Pectin Lyase from Pseudomonas putida
2.2. Characterization of Purified Pectin Lyase from Pseudomonas putida
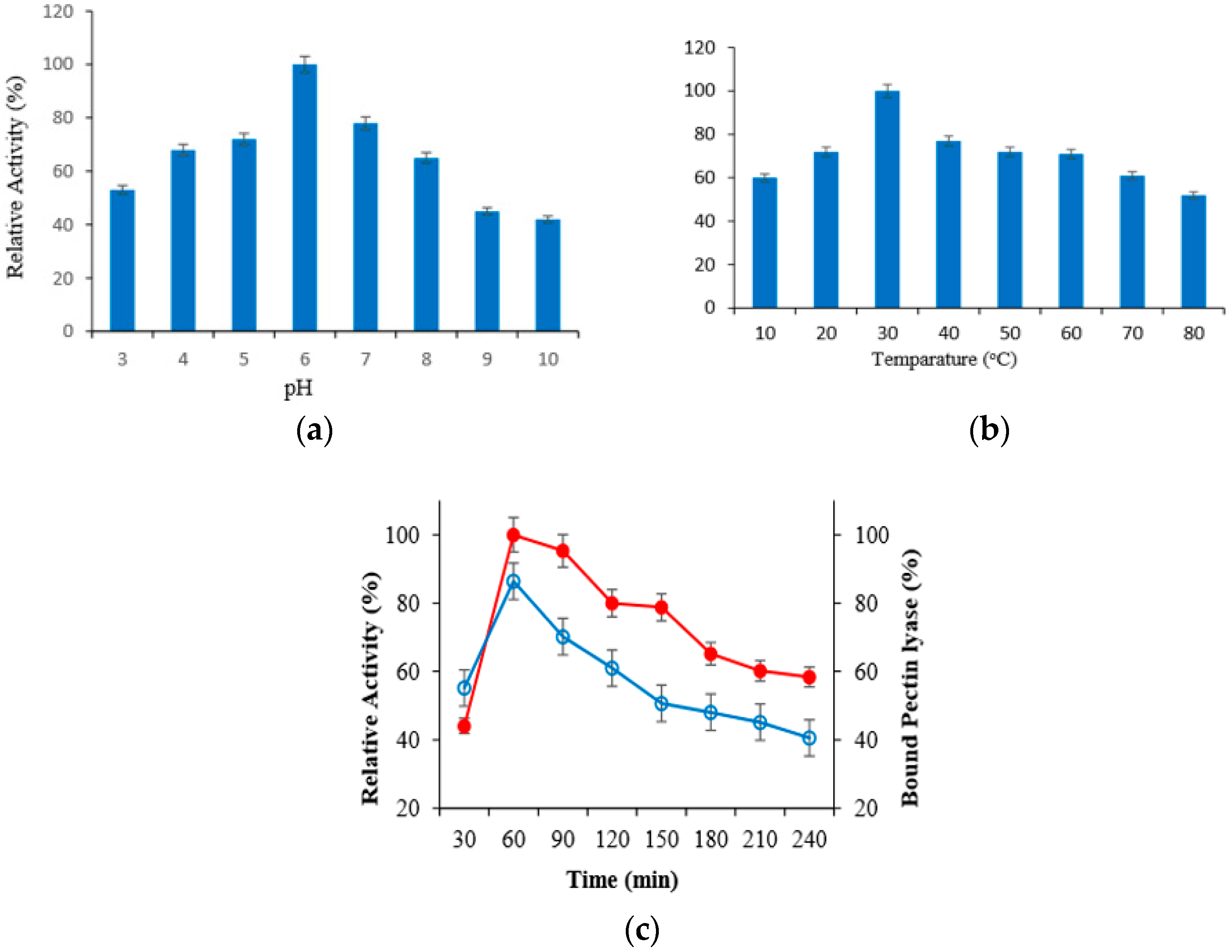
2.3. Effect of Incubation Time, pH and Temperature to Immobilization of PL Enzyme on MLf
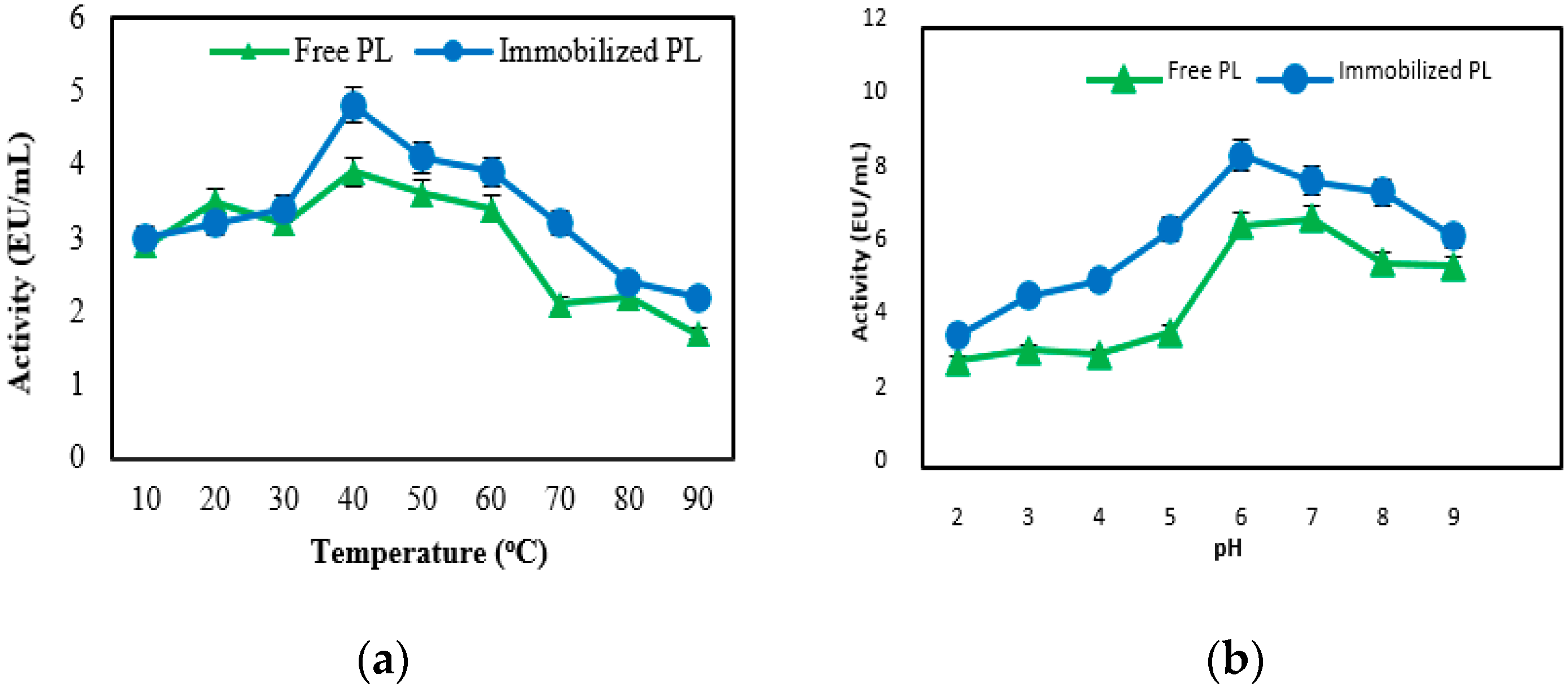
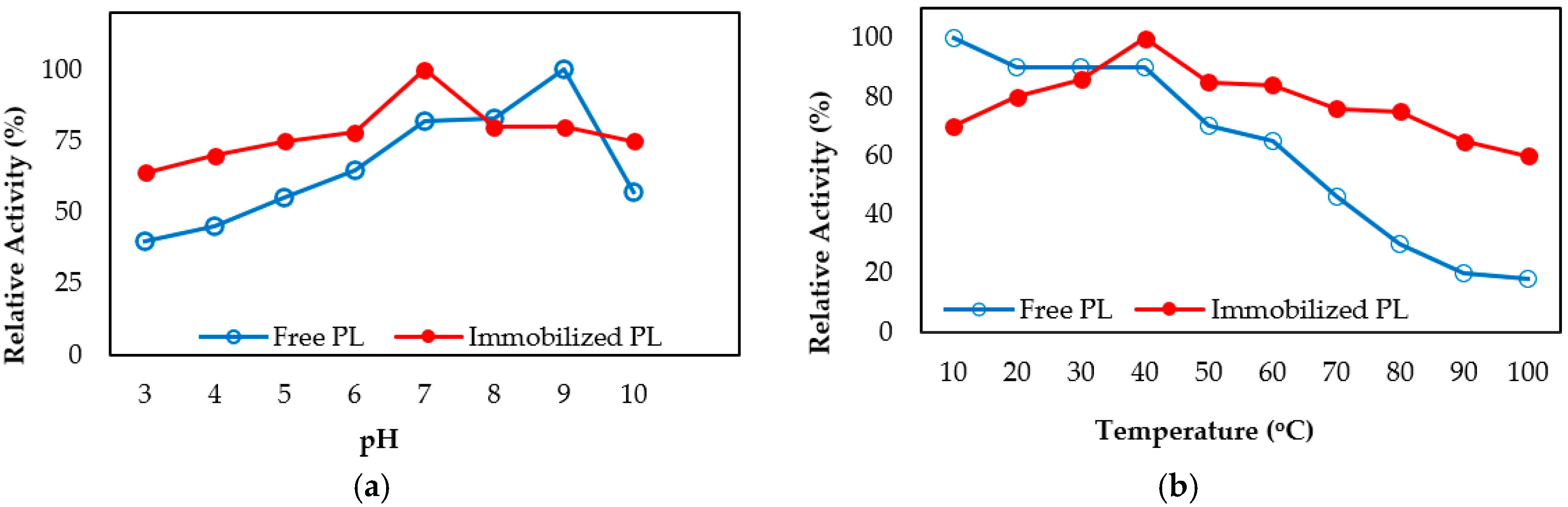
2.4. Biochemical Properties of Free and Immobilized Pectin Lyase
Effect of Temperature and pH
2.5. pH Stability and Thermal Stability
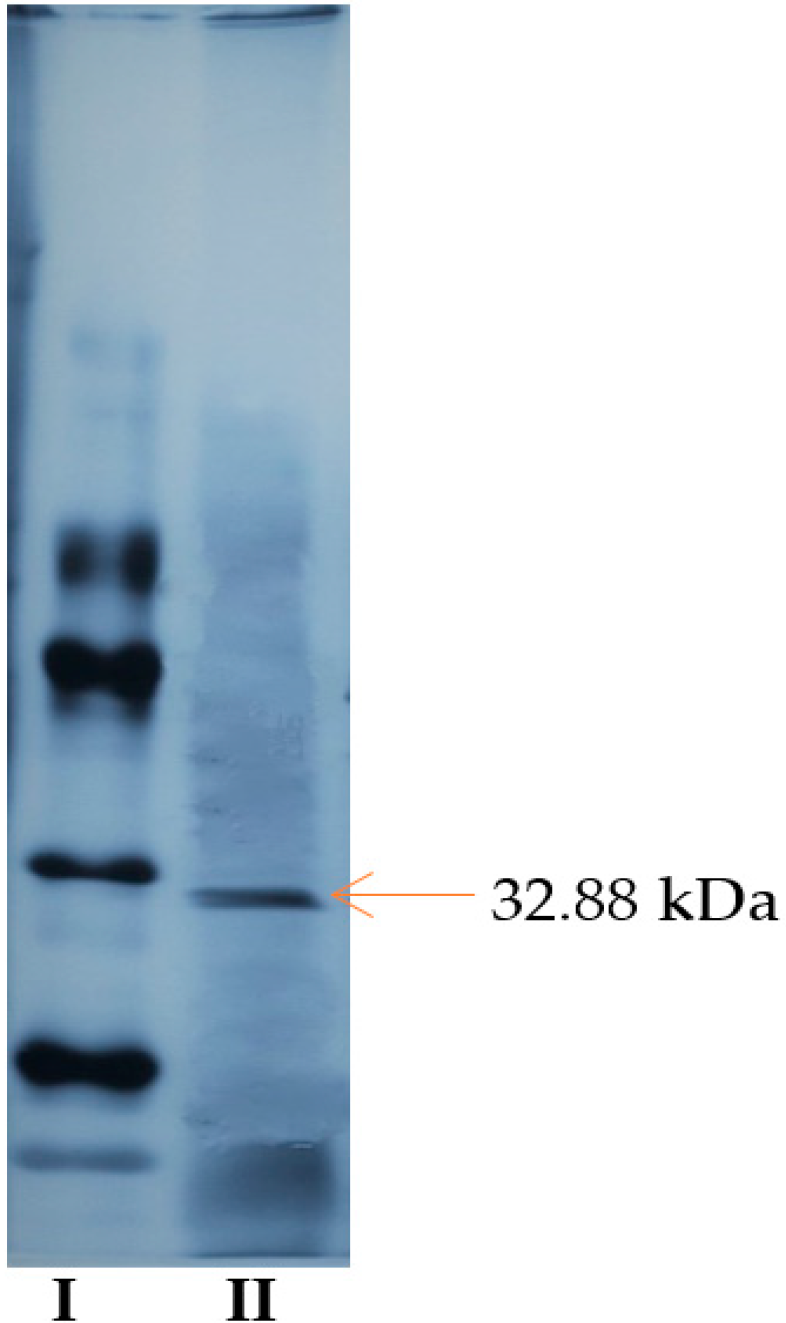
2.6. SDS-PAGE Electrophoresis
2.7. Structural Characterization of Support
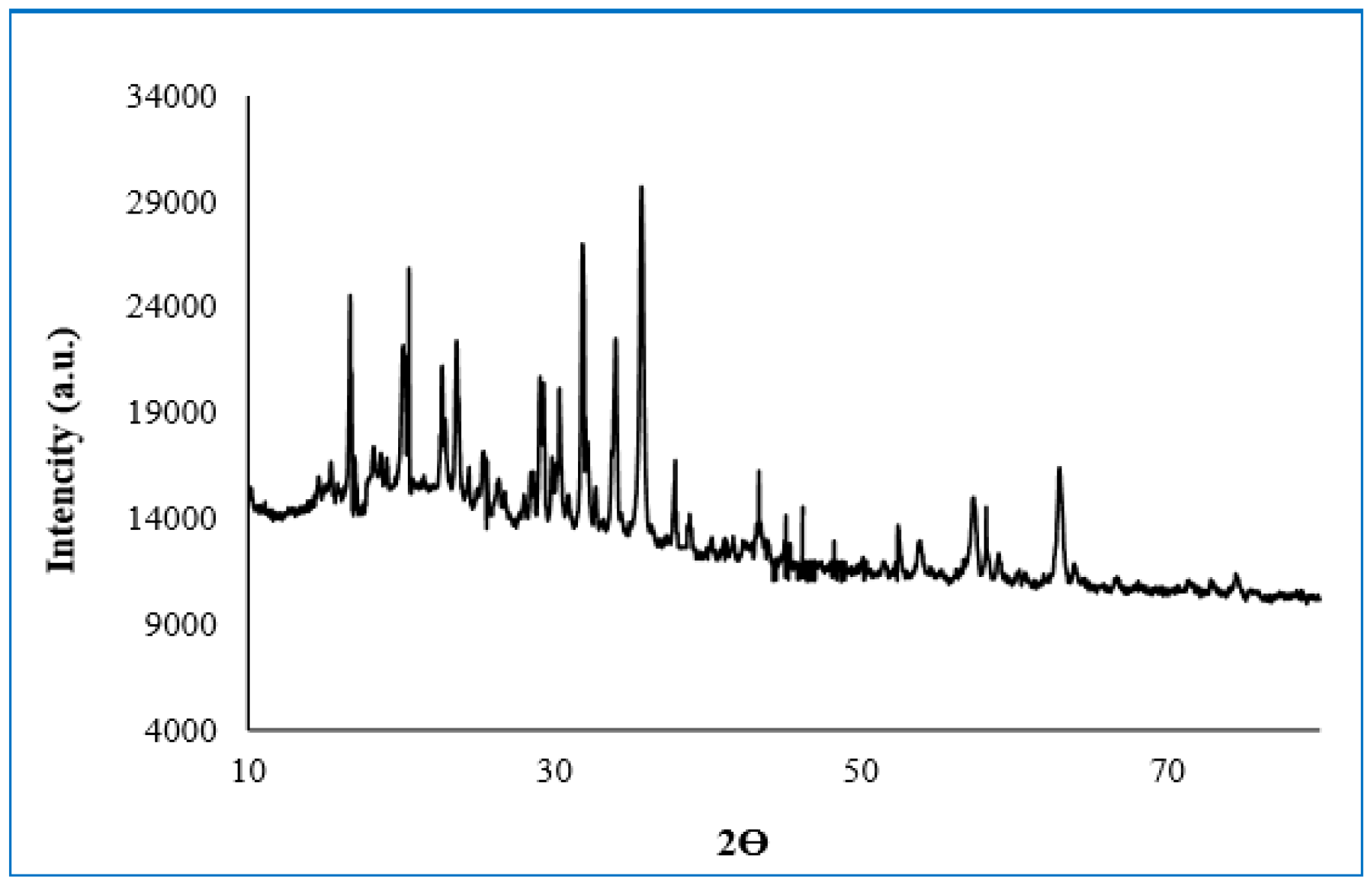
2.8. X-ray Diffraction (XRD) Analysis
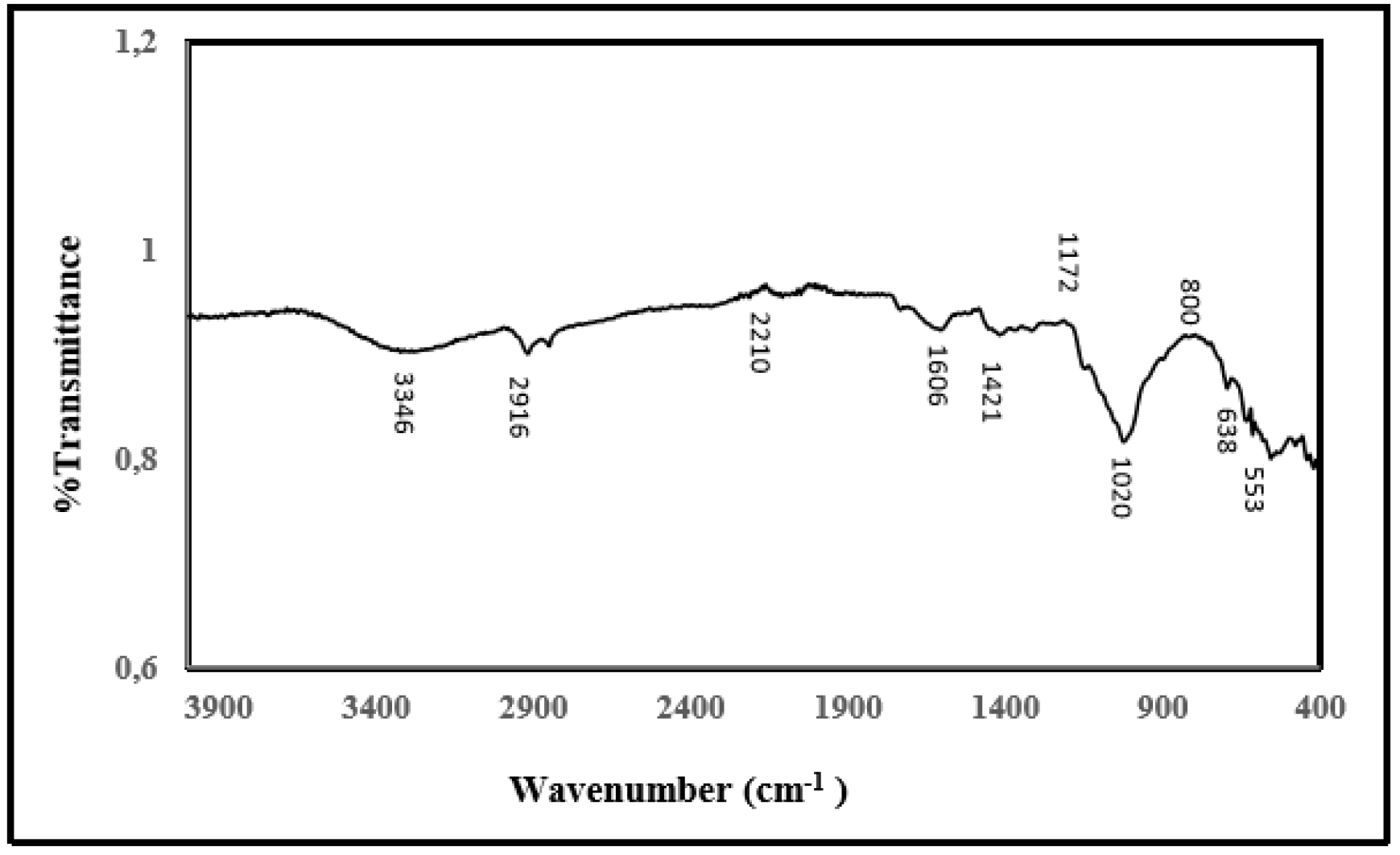
2.9. FT-IR Analysis
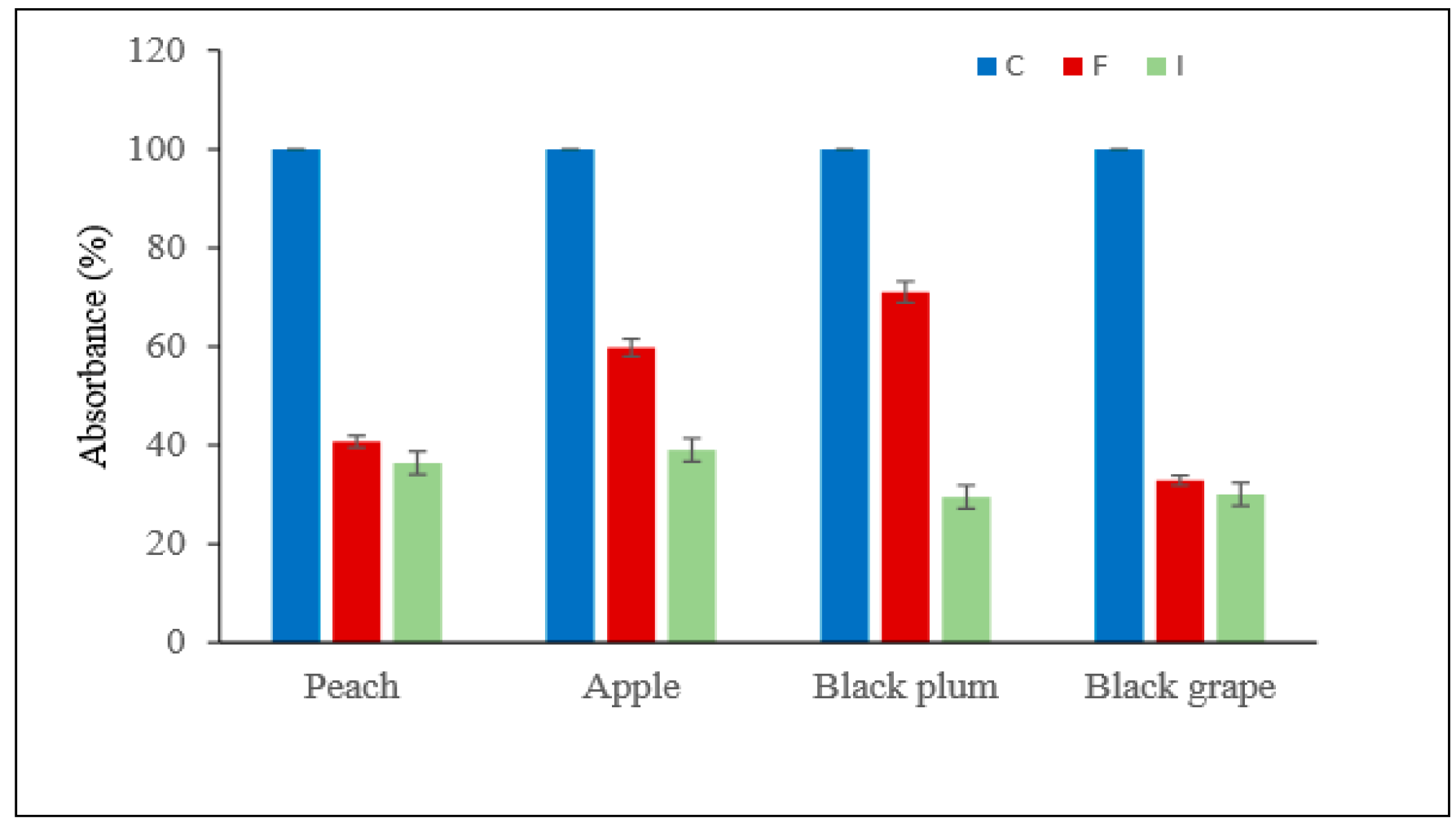
2.10. Application of Immobilised Pectin Lyase Enzyme in Juice Clarification
3. Conclusions
4. Materials and Methods
4.1. Bacterial Isolation
4.2. Identification Tests
4.3. Identification of Microorganism
4.4. Genotype Characterization of Bacteria
4.5. PL Enzyme Production with Solid Culture Fermentation
4.6. Purification of PL from Pseudomonas Putida Using TPP Method
4.7. Pectin Lyase Activity Assay
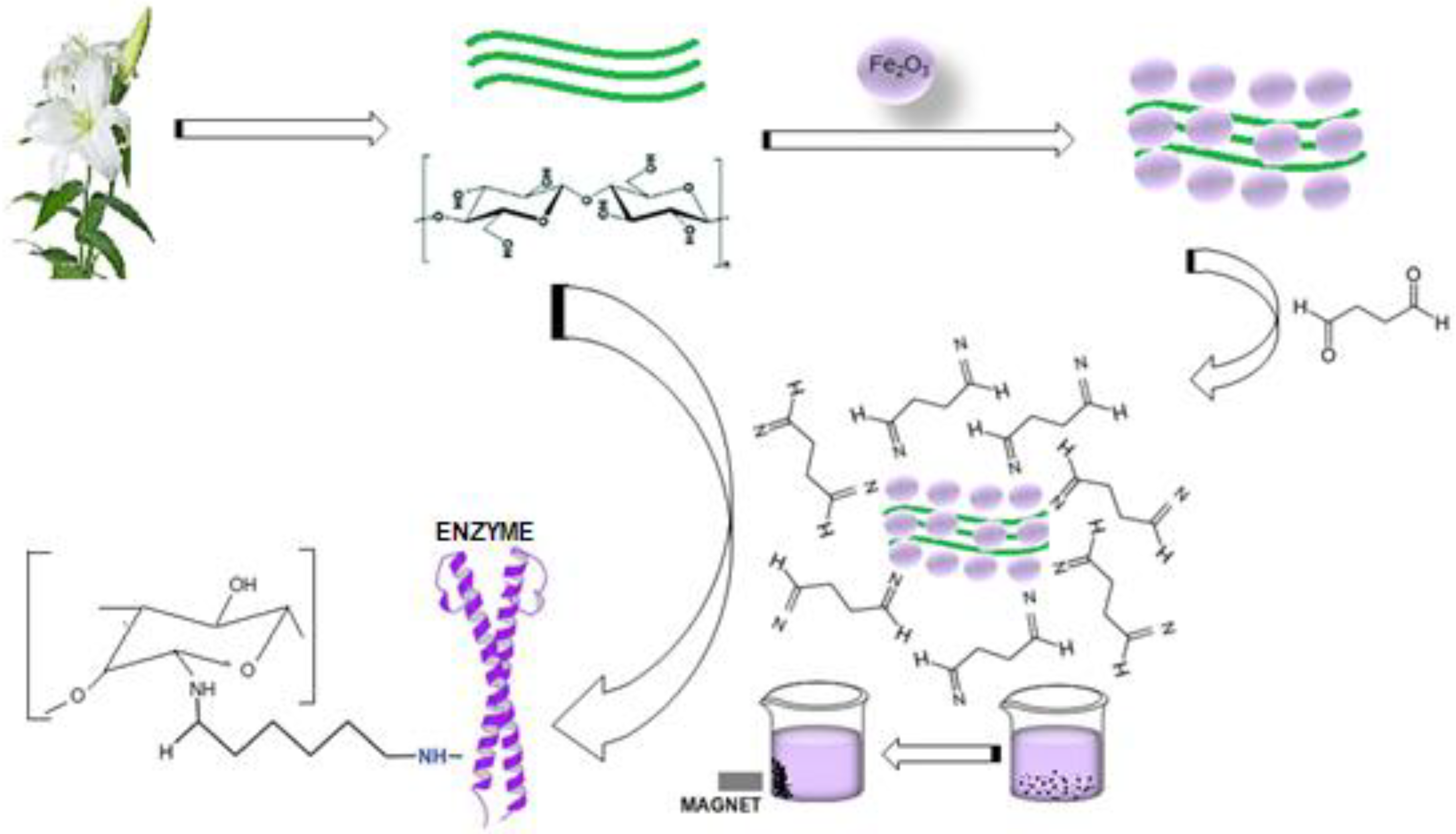
4.8. Preparation of Nano-Magnetic Lily Flowers (N-MLf) and Immobilization of Purified Pectin Lyase
4.9. Protein Determination
4.10. Preparation of Nanomagnetite Flowers of Lily (Lilium candidum L.)
4.11. Determination of Optimum Conditions for Immobilization of Purified PL Enzyme onto Modified Lily Flowers with γ-Fe3O4 NPs
4.12. Determination of the Optimum pH and Stable pH
4.13. Determination of Optimum and Stable Temperature Values
4.14. Characterization of Free and Immobilized Enzymes
4.15. Application of Immobilized Enzyme in Fruit Juice Clarity
4.16. Structural Characterization of Support Material
4.17. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Demir, N.; Nadaroğlu, H.; Tasgın, E.; Adıgüzel, A.; Gulluce, M. Purification and Characterization of a pectin lyase produced by Geobacillus stearothermophilus (Ah22) and fruit juice application. Ann. Microbiol. 2011, 61, 939–946. [Google Scholar] [CrossRef]
- Nadaroglu, H.; Taskin, E.; Adiguzel, A.; Gulluce, M.; Demir, N. Production of a novel pectin lyase from Bacillus pumilus (P9), purification and characterisation and fruit juice application. Rom. Biotechnol. Lett. 2010, 15, 5167–5176. [Google Scholar]
- Márquez, A.L.; Zavala-Páramo, M.G.; López-Romero, E.; Calderón-Cortés, N.; López-Gómez, R.; Conejo-Saucedo, U.; Cano-Camacho, H. Cloning and characterization of a pectin lyase gene from Colletotrichum lindemuthianum and comparative phylogenetic/structural analyses with genes from phytopathogenic and saprophytic/opportunistic microorganisms. BMC Microbiol. 2011, 11, 260. [Google Scholar]
- Fontana, R.C.; Silveira, M.M. Influence of pectin, glucose, and pH on the production of endo- and exo-polygalacturonase by Aspergillus oryzae in liquid medium. Braz. J. Chem. Eng. 2012, 29, 683–690. [Google Scholar] [CrossRef]
- Demir, N.; Nadaroglu, H.; Demir, Y.; Isık, C.; Taskin, E.; Adiguzel, A.; Gulluce, M. Purification and characterisation of an alkaline pectin lyase produced by a newly isolated Brevibacillus borstelensis (P35), and its applications in fruit juice and oil extraction. Eur. Food Res. Technol. 2014, 239, 127–135. [Google Scholar] [CrossRef]
- Mohamadi, A.S.; Shahbazi, S.; Behgar, M.; Fard, S.M.; Askari, H. A study of pectinase enzyme activity changes in gamma- irradiated Trichoderma reesei mutants. Int. J. Farm. Allied Sci. 2014, 3, 555–561. [Google Scholar]
- Pedrolli, D.B.; Monteiro, A.C.; Gomes, E.; Carmona, E.C. Pectin and Pectinases: Production, Characterization and Industrial Application of Microbial Pectinolytic Enzymes. Open Biotechnol. J. 2009, 3, 9–18. [Google Scholar] [CrossRef]
- Busto, M.D.; García-Tramontín, K.E.; Ortega, N.; Perez-Mateos, M. Preparation and properties of an immobilized pectinlyase for the treatment of fruit juices. Bioresour. Technol. 2006, 97, 1477–1483. [Google Scholar] [CrossRef]
- Sorrivas, V.; Genovese, D.B.; Lozano, J.E. Effect of pectinolytic and amylolytic enzymes on apple juice turbidity. J. Food Proc. Preserv. 2006, 30, 118–133. [Google Scholar] [CrossRef]
- Magro, L.D.; Silva de Moura, S.; Backes, B.E.; Weber de Menezes, E.; Valmir Benvenutti, E.; Nicolodi, S.; Kleind, M.P.; Fernandez-Lafuentee, R.; Rodriguesa, R.C. Immobilization of pectinase on chitosan-magnetic particles: Influence of particle preparation protocol on enzyme properties for fruit juice clarification. Biotechnol. Rep. 2019. [Google Scholar] [CrossRef]
- Sharma, A.K.; Gupta, M.N. Three phase partitioning of carbohydrate polymers: Separation and purification of alginates. Carbohydr. Polym. 2002, 48, 391–395. [Google Scholar] [CrossRef]
- Onem, H.; Nadaroglu, H. Immobilization of Purified Phytase Enzyme from Tirmit (Lactarius volemus) on Coated Chitosan with Iron Nanoparticles and Investigation of Its Usability in Cereal Industry. Iran. J. Sci. Technol. Trans. Sci. 2018, 42, 1063–1075. [Google Scholar] [CrossRef]
- Kohli, P.; Gupta, R. Application of calcium alginate immobilized and crude pectin lyase from Bacillus cereus in degumming of plant fibres. Biocatal. Biotransform. 2019. [Google Scholar] [CrossRef]
- Sheldon, R.A. Enzyme immobilization: The quest for optimum performance. Adv. Synth. Catal. 2007, 349, 1289–1307. [Google Scholar] [CrossRef]
- Mateo, C.; Palomo, J.M.; Fernandez-Lorente, G.; Guisan, J.M.; Fernandez-Lafuente, R. Improvement of enzyme activity, stability and selectivity via immobilization techniques. Enzyme Microb. Technol. 2007, 40, 1451–1463. [Google Scholar] [CrossRef]
- Zdarta, J.; Meyer, A.S.; Jesionowski, T.I.D.; Pinelo, M. A General Overview of Support Materials for Enzyme Immobilization: Characteristics, Properties, Practical Utility. Catalysts 2018, 8, 2–27. [Google Scholar] [CrossRef]
- Ramirez, H.L.; Brizuela, L.G.; Iranzo, J.U.; Arevalo-Villena, M.; Perez, A.I.B. Pectinase immobilization on a chitosan-coated chitin support. J. Food Process. Eng. 2016, 39, 97–107. [Google Scholar] [CrossRef]
- Irshad, M.; Murtza, A.; Zafar, M.; Bihatti, K.H.; Rehman, A.; Anwar, Z. Chitosan-immobilized pectinolytics with novel catalytic features and fruit juice clarification potentialities. Int. J. Biol. Macromol. 2017, 104, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Rehman, H.U.; Aman, A.; Silipo, A.; Qader, S.A.U.; Molinaro, A.; Ansari, A. Degradation of complex carbohydrate: Immobilization of pectinase from Bacillus licheniformis KIBGE-IB21 using calcium alginate as a support. Food Chem. 2013, 139, 1081–1086. [Google Scholar] [CrossRef] [PubMed]
- Horchani, H.; Aissa, I.; Ouertani, S.; Zarai, Z.; Gargouri, Y.; Sayari, A. Staphylococcal lipases: Biotechnological applications. J. Mol. Catal. B Enzym. 2012, 76, 125–132. [Google Scholar] [CrossRef]
- Vijayaraghavan, K.; Yamini, D.; Ambika, V.; Sravya Sowdamini, N. Trends in inulinase production—A review. Crit. Rev. Biotechnol. 2009, 29, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Homaei, A.A.; Sariri, R.; Vianello, F.; Stevanato, R. Enzyme immobilization: An update. J. Chem. Biol. 2013, 6, 185–205. [Google Scholar] [CrossRef] [PubMed]
- Krajewska, B. Application of chitin- and chitosan-based materials for enzyme immobilizations: A review. Enzyme Microb. Technol. 2004, 35, 126–139. [Google Scholar] [CrossRef]
- Kurita, K. Controlled functionalization of the polysaccharide chitin. Prog. Polym. Sci. 2001, 26, 1921–1971. [Google Scholar] [CrossRef]
- Peter, M. Applications and environmental aspects of chitin and chitosan. J. Macromol. Sci. 1995, 32, 629–640. [Google Scholar] [CrossRef]
- Mehmood, T.; Saman, T.; Irfan, M.; Anwar, F.; Ikram, M.S.; Tabassam, Q. Pectinase Production from Schizophyllum commune Through Central, Composite Design Using Citrus Waste and Its Immobilization for Industrial Exploitation. Waste Biomass Valorization 2018, 10, 2527–2536. [Google Scholar] [CrossRef]
- Yadav, S.; Yadav, P.K.; Yadav, D.; Yadav, K.D.S. Purification and characterization of an alkaline pectin lyase from Aspergillus flavus. Process Biochem. 2008, 43, 547–552. [Google Scholar] [CrossRef]
- Babagil, A. Purification and characterization of pectin lyase enzymes from microorganism, preparation of enzymic Ca2+-hybrid nanoflower structure and the investigation of use in fruit juice clarification. Ph.D. Thesis, Ataturk University, Erzurum, Turkey, 2018. [Google Scholar]
- Lei, Z.; Bi, S. The silica-coated chitosan particle from a layer-by-layer approach for pectinase immobilization. Enzym. Microb. Technol. 2007, 40, 1442–1447. [Google Scholar] [CrossRef]
- Banu, R.; Kalpana, A.; Devi, A.M.; Gnanaprabhal, G.R.; Pradeep, B.V.; Palaniswamy, M. Production and characterization of pectinase enzyme from Penicillium chrysogenum. Indian J. Sci. Technol. 2010, 3, 377–381. [Google Scholar]
- Ogawa, K.; Hirano, S.; Miyanishi, T.; Yui, T.; Watanabe, T. A new polymorph of chitosan. Macromolecules 1984, 17, 973–975. [Google Scholar] [CrossRef]
- Cerreti, M.; Markosova, K.; Esti, M.; Rosenberg, M.; Rebros, M. Immobilisation of pectinases into PVA gel for fruit juice application. Int. J. Food Sci. Technol. 2017, 52, 531–539. [Google Scholar] [CrossRef]
- Dey, T.B.; Banerjee, R. Application of decolourized and partially purified polygalacturonase and α-amylase. Braz. J. Microbiol. 2014, 45, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Urlaub, R. Advantages of enzymatic apple mash treatment and pomace liquefaction. Fruit Process. 1996, 6, 399–406. [Google Scholar]
- Yuan, P.; Meng, K.; Huang, H.; Shi, P.; Luo, H.; Yang, P.; Yao, B. A novel acidic and low-temperature-active endo-polygalacturonase from Penicillium sp. CGMCC 1669 with potential for application in apple juice clarification. Food Chem. 2011, 129, 1369–1375. [Google Scholar] [CrossRef]
- Singh, S.; Gupta, R. Apple juice clarification using fungal pectinolytic enzyme and gelatine. Indian J. Biotechnol. 2004, 3, 573–576. [Google Scholar]
- Xu, S.X.; Qin, X.; Liu, B.; Zhang, D.Q.; Zhang, W.; Wu, K.; Zhang, Y.H. An acidic pectin lyase from Aspergillus niger with favourable efficiency in fruit juice clarification. Lett. Appl. Microbiol. 2014, 60, 181–187. [Google Scholar] [CrossRef]
- Woo, P.C.Y.; Fung, A.M.Y.; Lau, S.K.P.; Yuen, K.Y. Identification by 16S rRNA gene sequencing of Lactobacillus salivarius bacteremic cholecystitis. J. Clin. Microbiol. 2002, 40, 265–267. [Google Scholar] [CrossRef]
- Forsman, P.; Tilsala-Timisjarvi, A.; Alatossava, T. Identification of staphylococcal and streptococcal causes of bovine mastisis using 16S-23S rRNA spacer regions. Microbiology 1997, 143, 3491–3500. [Google Scholar] [CrossRef]
- Garbers, I.; Britz, T.J.; Witthuhn, R.C. Ctypification and identification of the microbial consortium present in Kefir grains. World J. Microbiol. Biotech. 2004, 20, 687–693. [Google Scholar] [CrossRef]
- Nedjma, M.; Hoffmann, N.; Belarbi, A. Selective and sensitive detection of pectin lyase activity using a colorimetric test: Application to the screening of microorganisms possessing pectin lyase activity. Anal. Biochem. 2001, 291, 290–296. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Nadaroğlu, H.; Alayl Gungor, A.; Ince, S. Synthesis of nanoparticles by green synthesis method. Int. J. Innov. Res. Rev. 2017, 1, 6–9. [Google Scholar]
- Soares, M.M.C.N.; Sılva, R.; Gomes, E. Screening of bacterial strains for pectinolytic activity: Characterization of the polygalacturonase produced by Bacillus sp. Rev. Microbiol. 1999, 30, 299–303. [Google Scholar] [CrossRef]
Sample Availability: Samples of compound 1 are available from the authors. |










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme Fraction | Volume (mL) | Activity (EU/mL) | Total Activity (EU/mL) % | Specific Activity (EU/mL) | Protein (mg protein/mL) | Purification Fold |
|---|---|---|---|---|---|---|
| Crude extract | 60 | 263.2 ± 0.01 | 15792 100 | 0.17 | 1530.9 ± 0.011 | ----- |
| n-Butanol (1:0.5) | 20 | 224.6 ± 0.04 | 4492 85 | 1.16 | 192.4 ± 0.0171 | 6.82 |
| 1. (NH4)2SO4 (%60) | 20 | 208.5 ± 0.011 | 4170.8 79.2 | 5.49 | 37.97 ± 0.016 | 32.3 |
| 2. (NH4)2SO4 (%65) | 20 | 191.4 ± 0.012 | 3710 73 | 37.52 | 5.1 ± 0.014 | 220.7 |
| Pectin | Locust Bean Gum | Chitin | ||||
|---|---|---|---|---|---|---|
| Vmax | KM | Vmax | KM | Vmax | KM | |
| (µmol/Lmin) | (mg/mL) | (µmol/Lmin) | (mg/mL) | (µmol/Lmin) | (mg/mL) | |
| Pure PL | 18.62 | 1 | 1.45 | 0.65 | 1.26 | 0.5 |
| NMF-PL | 23.20 | 0.86 | 1.98 | 0.61 | 1.34 | 0.42 |
| Fruit (10 g) | Dry Weight | Increasing Volume (mL) | ||||
|---|---|---|---|---|---|---|
| Control * | Free PL | Immobilized PL | Control | Free PL | Immobilized PL | |
| DW(g) %D | DW(g) %D | DW(g) %D | ||||
| Peach | 0.244 2.4 | 0.168 1.7 | 0.202 0.2 | 7.0 | 7.0 | 8.5 |
| Apple | 0.161 1.6 | 0.152 1.5 | 0.144 1.4 | 8.0 | 8.5 | 9.2 |
| Black Plum | 0.233 2.3 | 0.19 1.9 | 0.102 1 | 7.5 | 7.0 | 9.2 |
| Black Grape | 0.121 1.2 | 0.114 1.1 | 0.065 0.6 | 7.0 | 7.0 | 8.0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tasgin, E.; Nadaroglu, H.; Babagil, A.; Demir, N. Immobilization of Purified Pectin Lyase from Pseudomonas putida onto Magnetic Lily Flowers (Lilium candidum L.) Nanoparticles and Applicability in Industrial Processes. Molecules 2020, 25, 2671. https://doi.org/10.3390/molecules25112671
Tasgin E, Nadaroglu H, Babagil A, Demir N. Immobilization of Purified Pectin Lyase from Pseudomonas putida onto Magnetic Lily Flowers (Lilium candidum L.) Nanoparticles and Applicability in Industrial Processes. Molecules. 2020; 25(11):2671. https://doi.org/10.3390/molecules25112671
Chicago/Turabian StyleTasgin, Esen, Hayrunnisa Nadaroglu, Aynur Babagil, and Nazan Demir. 2020. "Immobilization of Purified Pectin Lyase from Pseudomonas putida onto Magnetic Lily Flowers (Lilium candidum L.) Nanoparticles and Applicability in Industrial Processes" Molecules 25, no. 11: 2671. https://doi.org/10.3390/molecules25112671
APA StyleTasgin, E., Nadaroglu, H., Babagil, A., & Demir, N. (2020). Immobilization of Purified Pectin Lyase from Pseudomonas putida onto Magnetic Lily Flowers (Lilium candidum L.) Nanoparticles and Applicability in Industrial Processes. Molecules, 25(11), 2671. https://doi.org/10.3390/molecules25112671