A Limonoid, 7-Deacetoxy-7-Oxogedunin (CG-1) from Andiroba (Carapa guianensis, Meliaceae) Lowers the Accumulation of Intracellular Lipids in Adipocytes via Suppression of IRS-1/Akt-Mediated Glucose Uptake and a Decrease in GLUT4 Expression


Abstract
1. Introduction
2. Results
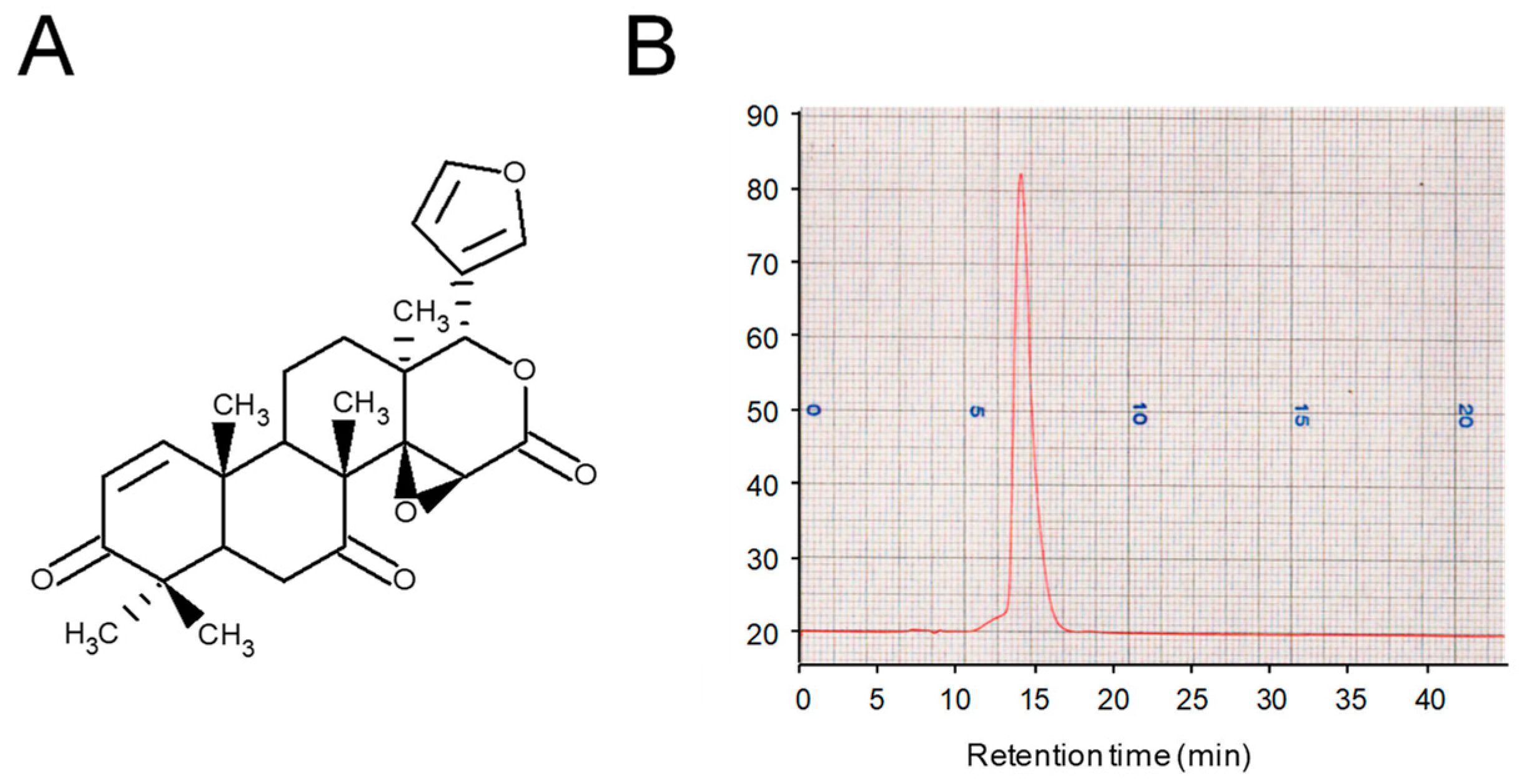
2.1. Extraction, Purification, and Structural Identification of the Limonoid CG-1
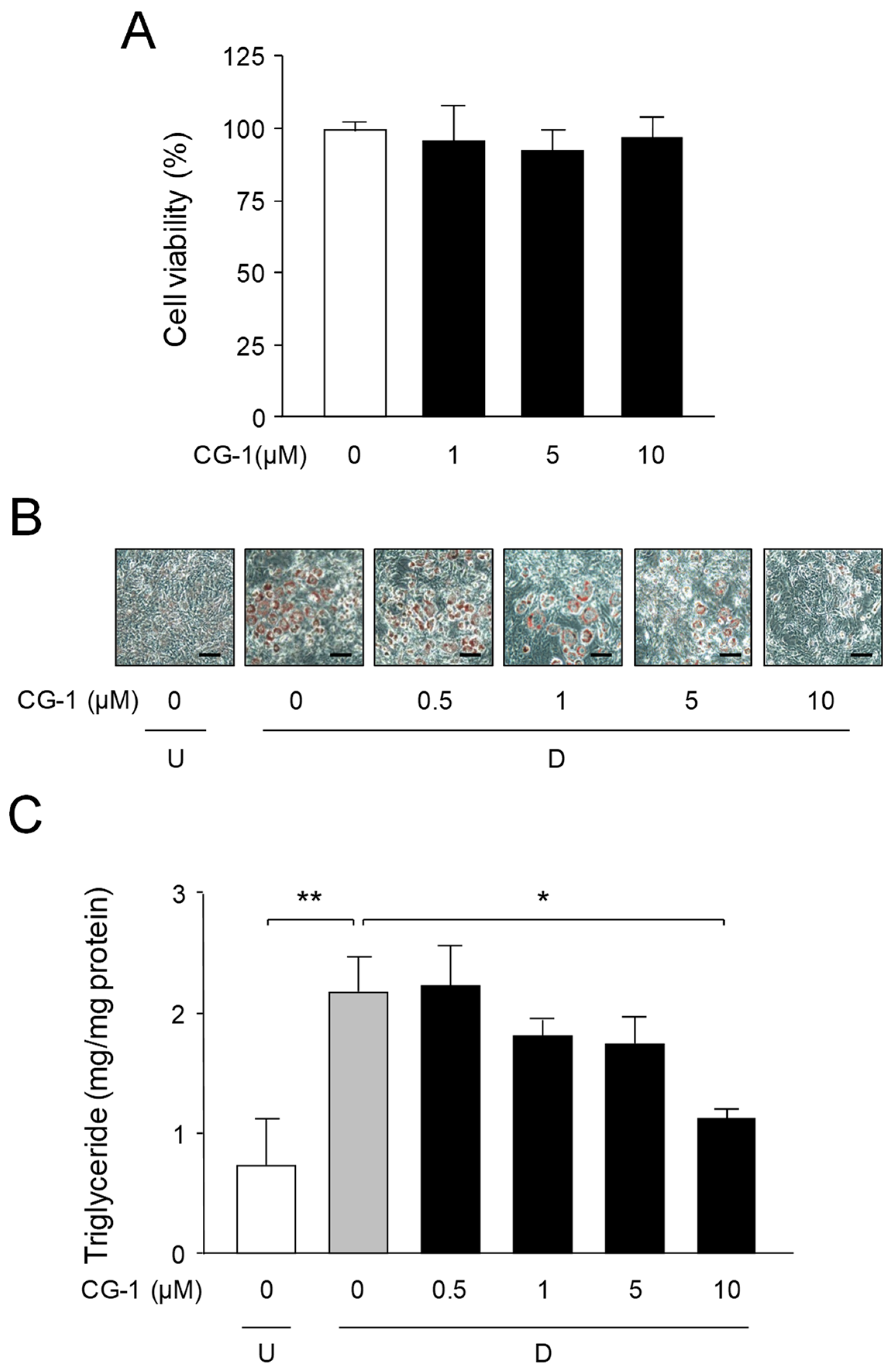
2.2. Decrease in Lipid Accumulation by CG-1
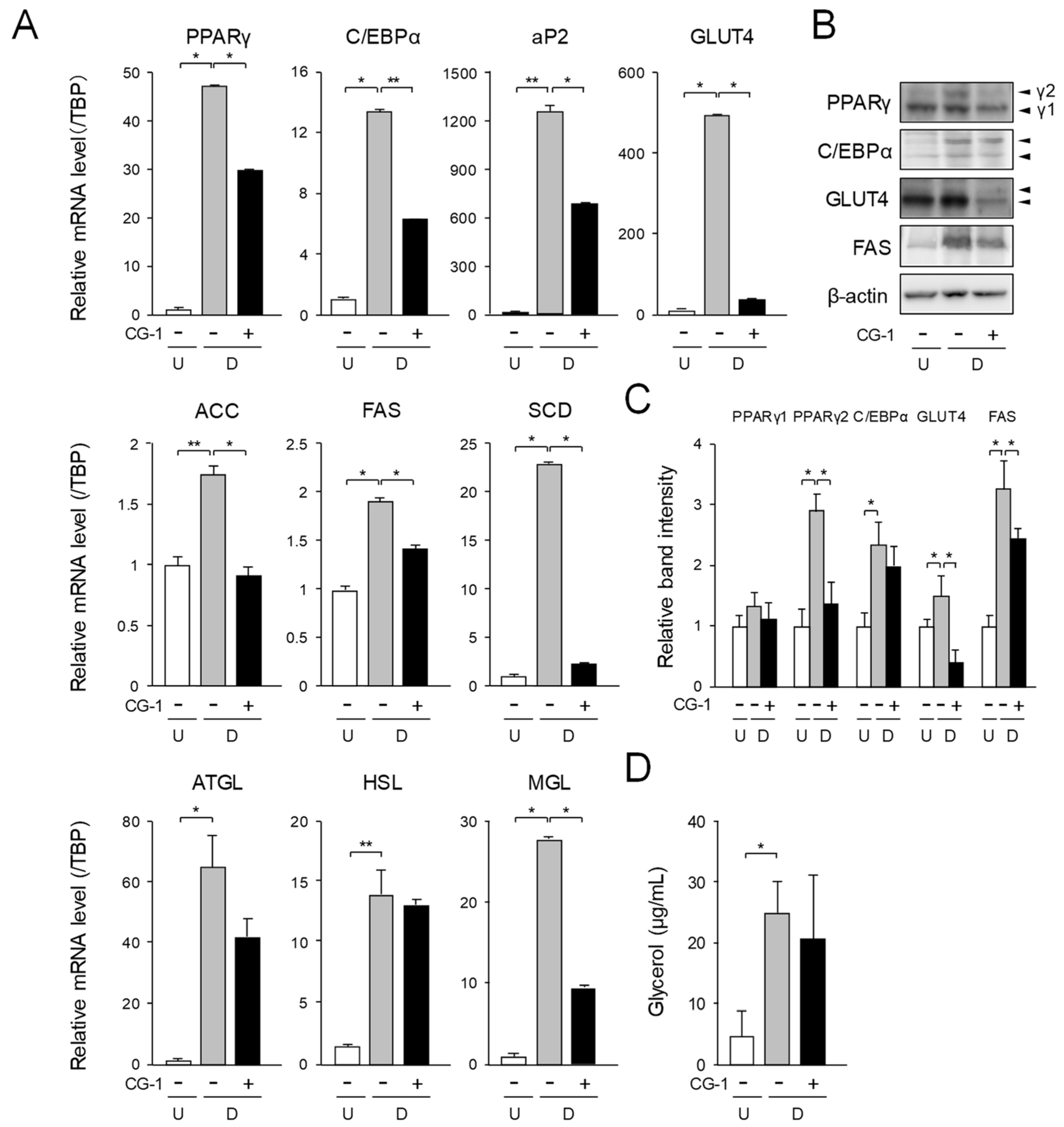
2.3. Effect of CG-1 on Expression of Adipogenic, Lipogenic, and Lipolytic Genes in Adipocytes
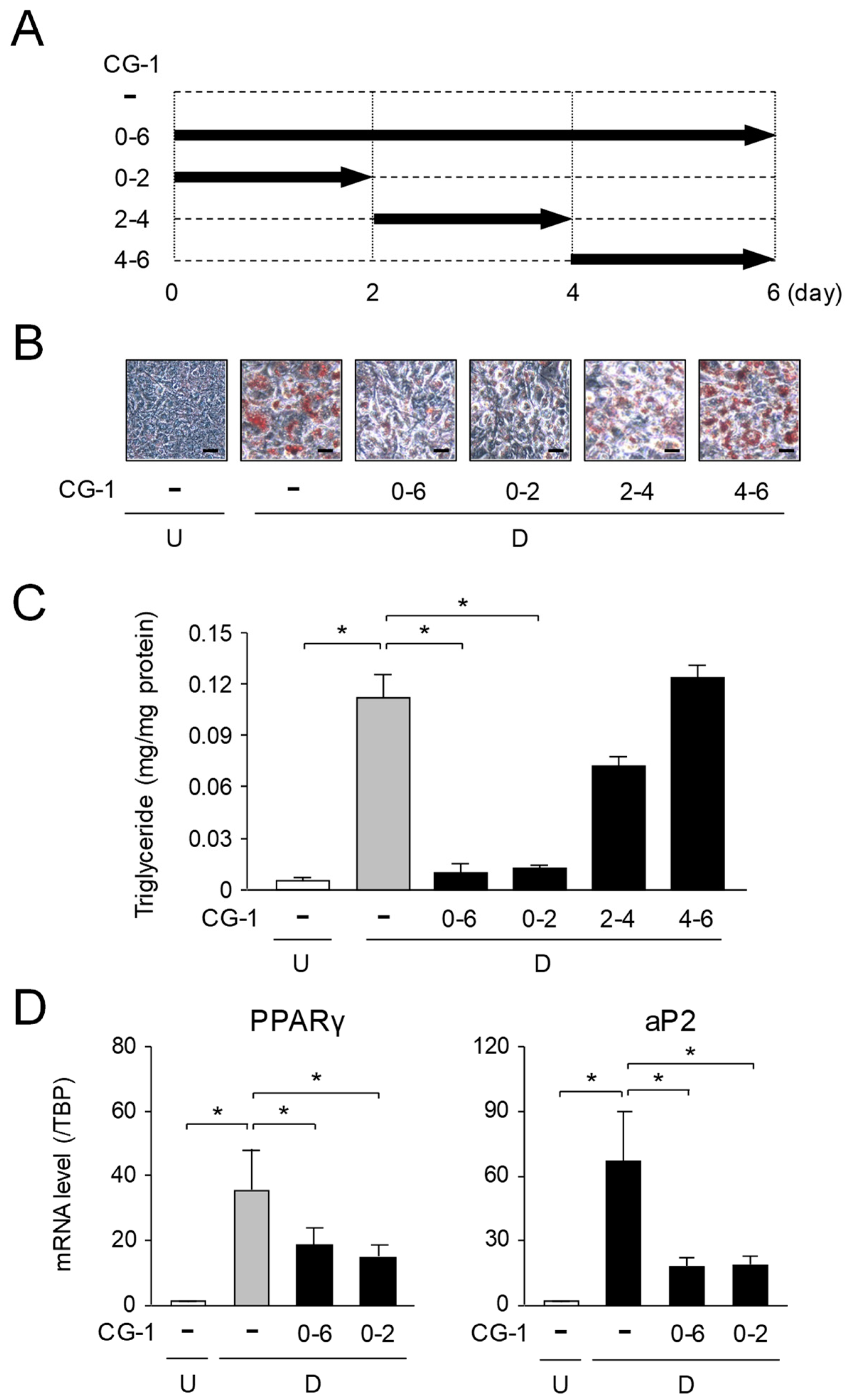
2.4. Suppression of Early Phase of Adipogenesis by CG-1
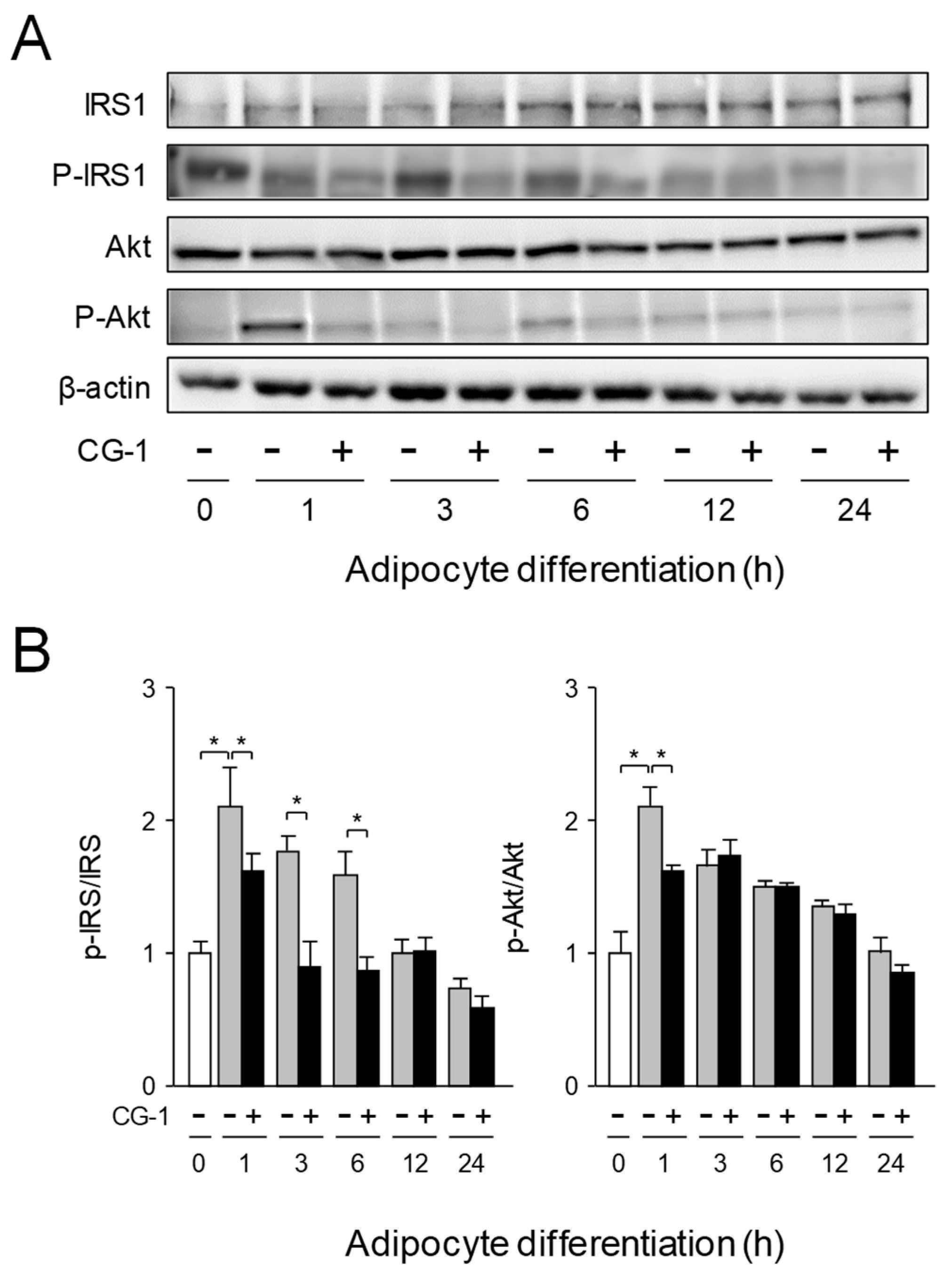
2.5. Inhibition of Activation of IRS-1/Akt Axis by CG-1
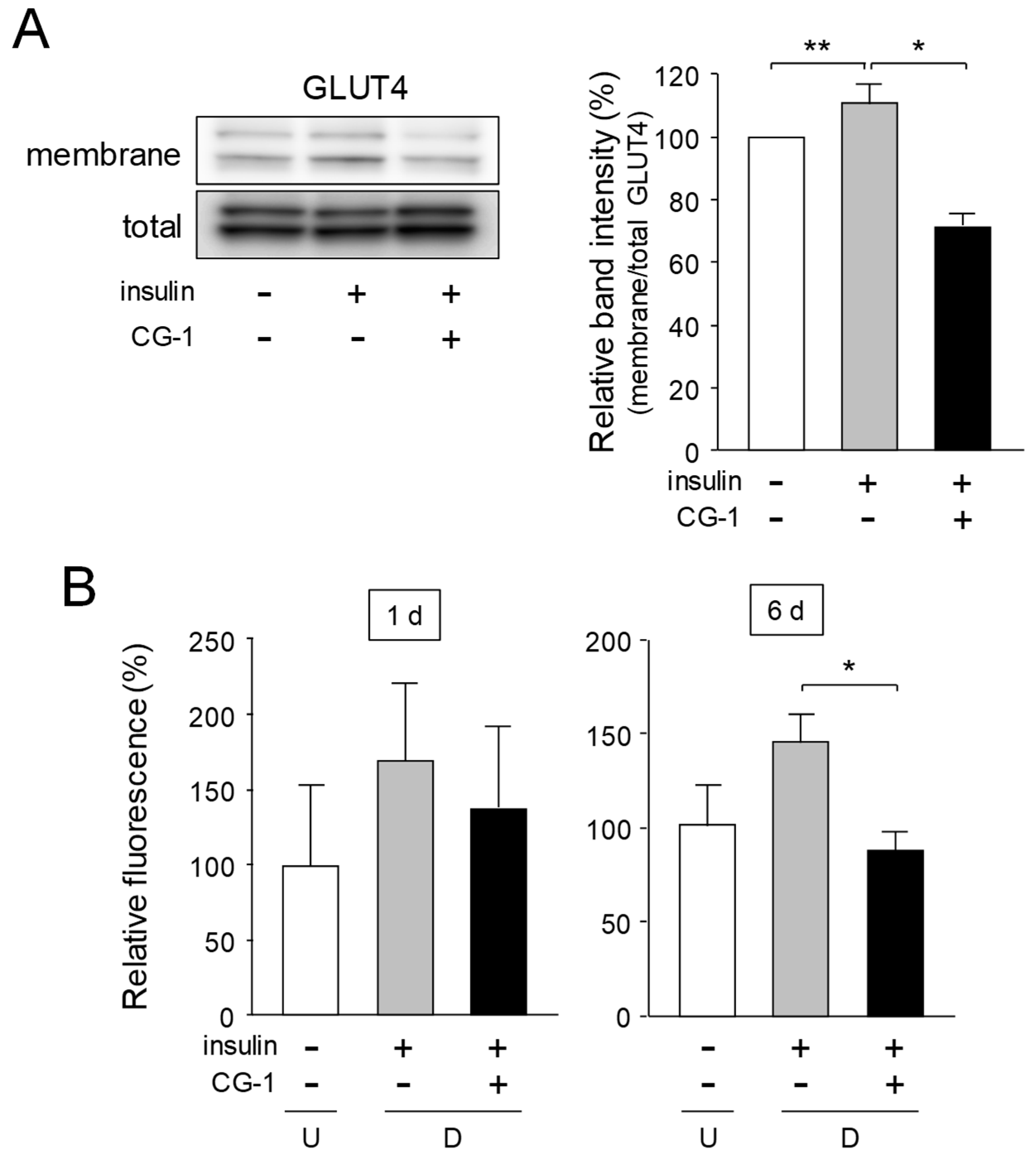
2.6. Inhibition of Translocation of GLUT4 and Glucose Uptake by CG-1
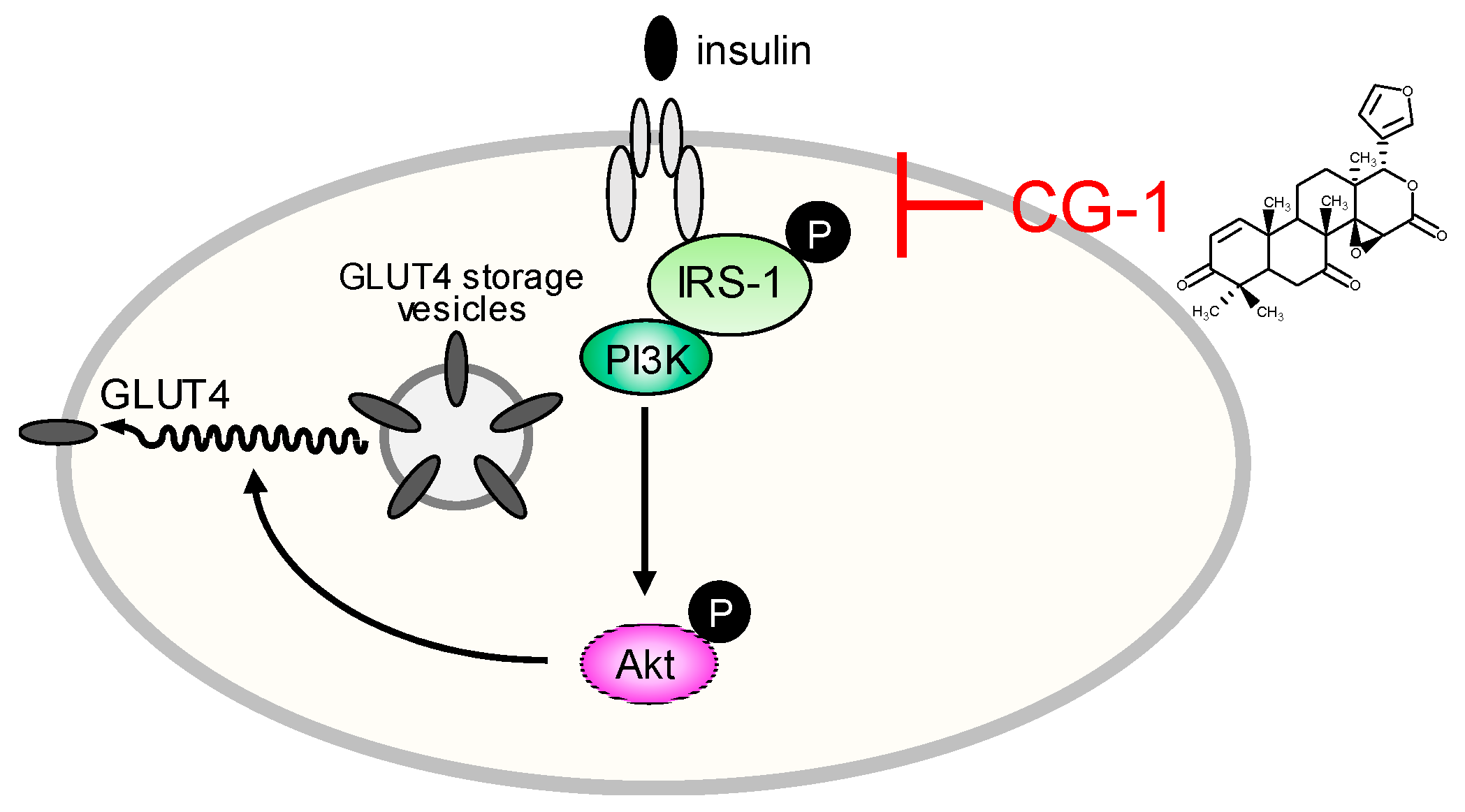
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Materials
5.2. Purification of A Limonoid CG-1 from Andiroba
5.3. Cell Culture
5.4. Cytotoxicity Assay
5.5. Intracellular Triglyceride Level
5.6. Quantitative PCR
5.7. Glycerol Release Assay
5.8. Western Blot Analysis
5.9. GLUT4 Translocation Assay
5.10. Glucose Uptake Assay
5.11. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Finucane, M.M.; Stevens, G.A.; Cowan, M.J.; Danaei, G.; Lin, J.K.; Paciorek, C.J.; Singh, G.M.; Gutierrez, H.R.; Lu, Y.; Bahalim, A.N.; et al. National, regional, and global trends in body-mass index since 1980: Systematic analysis of health examination surveys and epidemiological studies with 960 country-years and 9.1 million participants. Lancet 2011, 377, 557–567. [Google Scholar] [CrossRef]
- Attie, A.D.; Scherer, P.E. Adipocyte metabolism and obesity. J. Lipid Res. 2009, 50, S395–S399. [Google Scholar] [CrossRef]
- Cornier, M.A.; Dabelea, D.; Hernandez, T.L.; Lindstrom, R.C.; Steig, A.J.; Stob, N.R.; Van Pelt, R.E.; Wang, H.; Eckel, R.H. The metabolic syndrome. Endocr. Rev. 2008, 29, 777–822. [Google Scholar] [CrossRef]
- Ali, A.T.; Hochfeld, W.E.; Myburgh, R.; Pepper, M.S. Adipocyte and adipogenesis. Eur. J. Cell Biol. 2013, 92, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Lefterova, M.I.; Lazar, M.A. New developments in adipogenesis. Trends Endocrinol. Metab. 2009, 20, 107–114. [Google Scholar] [CrossRef]
- Haeusler, R.A.; McGraw, T.E.; Accili, D. Biochemical and cellular properties of insulin receptor signalling. Nat. Rev. Mol. Cell Biol. 2018, 19, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Baron, A.D. Hemodynamic actions of insulin. Am. J. Physiol. 1994, 267, E187–E202. [Google Scholar] [CrossRef]
- Jaldin-Fincati, J.R.; Pavarotti, M.; Frendo-Cumbo, S.; Bilan, P.J.; Klip, A. Update on GLUT4 vesicle traffic: A cornerstone of insulin action. Trends Endocrinol. Metab. 2017, 28, 597–611. [Google Scholar] [CrossRef] [PubMed]
- Morigny, P.; Houssier, M.; Mouisel, E.; Langin, D. Adipocyte lipolysis and insulin resistance. Biochimie 2016, 125, 259–266. [Google Scholar] [CrossRef]
- Gualdani, R.; Cavalluzzi, M.M.; Lentini, G.; Habtemariam, S. The chemistry and pharmacology of citrus limonoids. Molecules 2016, 21, 1530. [Google Scholar] [CrossRef] [PubMed]
- Ono, E.; Inoue, J.; Hashidume, T.; Shimizu, M.; Sato, R. Anti-obesity and anti-hyperglycemic effects of the dietary citrus limonoid nomilin in mice fed a high-fat diet. Biochem. Biophys. Res. Commun. 2011, 410, 677–681. [Google Scholar] [CrossRef] [PubMed]
- Higby, R.H. The bitter constituents of navel and Valencia oranges. J. Am. Chem. Soc. 1938, 60, 3013–3018. [Google Scholar] [CrossRef]
- Costa-Silva, J.H.; Lima, C.R.; Silva, E.J.; Araujo, A.V.; Fraga, M.C.; Ribeiro, E.R.A.; Arruda, A.C.; Lafayette, S.S.; Wanderley, A.G. Acute and subacute toxicity of the Carapa guianensis Aublet (Meliaceae) seed oil. J. Ethnopharmacol. 2008, 116, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Henriques, M.; Penido, C. The therapeutic properties of Carapa guianensis. Curr. Pharm. Des. 2014, 20, 850–856. [Google Scholar] [CrossRef]
- Martins, K.; Raposo, A.; Klimas, C.A.; Veasey, E.A.; Kainer, K.; Wadt, L.H. Pollen and seed flow patterns of Carapa guianensis Aublet. (Meliaceae) in two types of Amazonian forest. Genet. Mol. Biol. 2012, 35, 818–826. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Miranda Junior, R.N.; Dolabela, M.F.; da Silva, M.N.; Povoa, M.M.; Maia, J.G. Antiplasmodial activity of the andiroba (Carapa guianensis Aubl., Meliaceae) oil and its limonoid-rich fraction. J. Ethnopharmacol. 2012, 142, 679–683. [Google Scholar] [CrossRef] [PubMed]
- Penido, C.; Costa, K.A.; Pennaforte, R.J.; Costa, M.F.; Pereira, J.F.; Siani, A.C.; Henriques, M.G. Anti-allergic effects of natural tetranortriterpenoids isolated from Carapa guianensis Aublet on allergen-induced vascular permeability and hyperalgesia. Inflamm. Res. 2005, 54, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Penido, C.; Costa, K.A.; Costa, M.F.; Pereira Jde, F.; Siani, A.C.; Henriques, M. Inhibition of allergen-induced eosinophil recruitment by natural tetranortriterpenoids is mediated by the suppression of IL-5, CCL11/eotaxin and NF-κB activation. Int. Immunopharmacol. 2006, 6, 109–121. [Google Scholar] [CrossRef]
- Penido, C.; Conte, F.P.; Chagas, M.S.; Rodrigues, C.A.; Pereira, J.F.; Henriques, M.G. Antiinflammatory effects of natural tetranortriterpenoids isolated from Carapa guianensis Aublet on zymosan-induced arthritis in mice. Inflamm. Res. 2006, 55, 457–464. [Google Scholar] [CrossRef]
- Araujo-Lima, C.F.; Fernandes, A.S.; Gomes, E.M.; Oliveira, L.L.; Macedo, A.F.; Antoniassi, R.; Wilhelm, A.E.; Aiub, C.A.F.; Felzenszwalb, I. Antioxidant activity and genotoxic assessment of crabwood (andiroba, Carapa guianensis Aublet) seed oils. Oxid. Med. Cell Longev. 2018, 2018, 3246719. [Google Scholar] [CrossRef]
- Oliveira, I.; Moragas Tellis, C.J.; Chagas, M.; Behrens, M.D.; Calabrese, K.D.S.; Abreu-Silva, A.L.; Almeida-Souza, F. Carapa guianensis Aublet (andiroba) seed oil: Chemical composition and antileishmanial activity of limonoid-rich fractions. Biomed. Res. Int. 2018, 2018, 5032816. [Google Scholar] [CrossRef]
- Ninomiya, K.; Miyazawa, S.; Ozeki, K.; Matsuo, N.; Muraoka, O.; Kikuchi, T.; Yamada, T.; Tanaka, R.; Morikawa, T. Hepatoprotective limonoids from andiroba (Carapa guianensis). Int. J. Mol. Sci. 2016, 17, 591. [Google Scholar] [CrossRef] [PubMed]
- Horiba, T.; Katsukawa, M.; Mita, M.; Sato, R. Dietary obacunone supplementation stimulates muscle hypertrophy, and suppresses hyperglycemia and obesity through the TGR5 and PPARγ pathway. Biochem. Biophys. Res. Commun. 2015, 463, 846–852. [Google Scholar] [CrossRef]
- Wong, C.P.; Kaneda, T.; Hadi, A.H.; Morita, H. Ceramicine B, a limonoid with anti-lipid droplets accumulation activity from Chisocheton ceramicus. J. Nat. Med. 2014, 68, 22–30. [Google Scholar] [CrossRef]
- Baba, S.; Ueno, Y.; Kikuchi, T.; Tanaka, R.; Fujimori, K. A limonoid Kihadanin B from immature Citrus unshiu peels suppresses adipogenesis through repression of the Akt-FOXO1-PPARγ axis in adipocytes. J. Agric. Food Chem. 2016, 64, 9607–9615. [Google Scholar] [CrossRef]
- Tanaka, Y.; Yamada, T.; In, Y.; Muraoka, O.; Kajimoto, T.; Tanaka, R. Absolute stereostructure of andirolides A–G from the flower of Carapa guianensis (Meliaceae). Tetrahedron 2011, 67, 782–792. [Google Scholar] [CrossRef]
- Boucher, J.; Kleinridders, A.; Kahn, C.R. Insulin receptor signaling in normal and insulin-resistant states. Cold Spring Harb. Perspect. Biol. 2014, 6. [Google Scholar] [CrossRef]
- Mackenzie, R.W.; Elliott, B.T. Akt/PKB activation and insulin signaling: A novel insulin signaling pathway in the treatment of type 2 diabetes. Diabetes Metab. Syndr. Obes. 2014, 7, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Czech, M.P. The GLUT4 glucose transporter. Cell Metab. 2007, 5, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Miki, H.; Yamauchi, T.; Suzuki, R.; Komeda, K.; Tsuchida, A.; Kubota, N.; Terauchi, Y.; Kamon, J.; Kaburagi, Y.; Matsui, J.; et al. Essential role of insulin receptor substrate 1 (IRS-1) and IRS-2 in adipocyte differentiation. Mol. Cell. Biol. 2001, 21, 2521–2532. [Google Scholar] [CrossRef]
- Rosen, E.D.; MacDougald, O.A. Adipocyte differentiation from the inside out. Nat. Rev. Mol. Cell Biol. 2006, 7, 885–896. [Google Scholar] [CrossRef] [PubMed]
- Petersen, K.F.; Shulman, G.I. Etiology of insulin resistance. Am. J. Med. 2006, 119, S10–S16. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, P.G.; Festuccia, W.T.; Houde, V.P.; St-Pierre, P.; Brule, S.; Turcotte, V.; Cote, M.; Bellmann, K.; Marette, A.; Deshaies, Y. Major involvement of mTOR in the PPARgamma-induced stimulation of adipose tissue lipid uptake and fat accretion. J. Lipid Res. 2012, 53, 1117–1125. [Google Scholar] [CrossRef] [PubMed]
- Yao-Borengasser, A.; Rassouli, N.; Varma, V.; Bodles, A.M.; Rasouli, N.; Unal, R.; Phanavanh, B.; Ranganathan, G.; McGehee, R.E., Jr.; Kern, P.A. Stearoyl-coenzyme A desaturase 1 gene expression increases after pioglitazone treatment and is associated with peroxisomal proliferator-activated receptor-gamma responsiveness. J. Clin. Endocrinol. Metab. 2008, 93, 4431–4439. [Google Scholar] [CrossRef] [PubMed]
- Deng, D.; Yan, N. GLUT, SGLT, and SWEET: Structural and mechanistic investigations of the glucose transporters. Protein Sci. 2016, 25, 546–558. [Google Scholar] [CrossRef]
- Abel, E.D.; Peroni, O.; Kim, J.K.; Kim, Y.B.; Boss, O.; Hadro, E.; Minnemann, T.; Shulman, G.I.; Kahn, B.B. Adipose-selective targeting of the GLUT4 gene impairs insulin action in muscle and liver. Nature 2001, 409, 729–733. [Google Scholar] [CrossRef] [PubMed]
- Guo, S. Insulin signaling, resistance, and the metabolic syndrome: Insights from mouse models into disease mechanisms. J. Endocrinol. 2014, 220, T1–T23. [Google Scholar] [CrossRef] [PubMed]
- Rask-Madsen, C.; Kahn, C.R. Tissue-specific insulin signaling, metabolic syndrome, and cardiovascular disease. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 2052–2059. [Google Scholar] [CrossRef]
- Kaestner, K.H.; Christy, R.J.; Lane, M.D. Mouse insulin-responsive glucose transporter gene: Characterization of the gene and trans-activation by the CCAAT/enhancer binding protein. Proc. Natl. Acad. Sci. USA 1990, 87, 251–255. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Watanabe, M.; Hisatake, M.; Fujimori, K. Fisetin suppresses lipid accumulation in mouse adipocytic 3T3-L1 cells by repressing GLUT4-mediated glucose uptake through inhibition of mTOR-C/EBPα signaling. J. Agric. Food Chem. 2015, 63, 4979–4987. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Nishiumi, S.; Ashida, H. Rapid preparation of a plasma membrane fraction from adipocytes and muscle cells: Application to detection of translocated glucose transporter 4 on the plasma membrane. Biosci. Biotechnol. Biochem. 2007, 71, 2343–2346. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession No. * | Forward Primer | Reverse Primer |
|---|---|---|---|
| PPARγ | NM_011146 | 5’-CAAGAATACCAAAGTGCGATCAA-3′ | 5′-GAGCTGGGTCTTTTCAGAATAATAAG-3′ |
| C/EBPα | NM_007678 | 5′-CTGGAAAGAAGGCCACCTC-3′ | 5′-AAGAGAAGGAAGCGGTCCA-3′ |
| aP2 | NM_024406 | 5′-CAGCCTTTCTCACCTGGAAG-3′ | 5′-TTGTGGCAAAGCCCACTC-3′ |
| GLUT4 | NM_009204 | 5′-GACGGACACTCCATCTGTTG-3′ | 5′-GCCACGATGGAGACATAGC-3′ |
| ACC | NM_133360 | 5′-GCGTCGGGTAGATCCAGTT-3′ | 5′-CTCAGTGGGGCTTAGCTCTG-3′ |
| FAS | NM_007988 | 5′-GTTGGGGGTGTCTTCAACC-3′ | 5′-GAAGAGCTCTGGGGTCTGG-3′ |
| SCD | NM_009127 | 5′-CGTCTGGAGGAACATCATTCT-3′ | 5′-CAGAGCGCTGGTCATGTAGT-3′ |
| ATGL | NM_001163689 | 5′-TGACCATCTGCCTTCCAGA-3′ | 5′-TGTAGGTGGCGCAAGACA-3′ |
| HSL | NM_010719 | 5′-GCACTGTGACCTGCTTGGT-3′ | 5′-CTGGCACCCTCACTCCATA-3′ |
| MGL | NM_011844 | 5′-TCGGAACAAGTCGGAGGT-3′ | 5′-TCAGCAGCTGTATGCCAAAG-3′ |
| TBP | NM_013684 | 5′-GTGATGTGAAGTTCCCCATAAGG-3′ | 5′-CTACTGAACTGCTGGTGGGTCA-3′ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsumoto, C.; Koike, A.; Tanaka, R.; Fujimori, K. A Limonoid, 7-Deacetoxy-7-Oxogedunin (CG-1) from Andiroba (Carapa guianensis, Meliaceae) Lowers the Accumulation of Intracellular Lipids in Adipocytes via Suppression of IRS-1/Akt-Mediated Glucose Uptake and a Decrease in GLUT4 Expression. Molecules 2019, 24, 1668. https://doi.org/10.3390/molecules24091668
Matsumoto C, Koike A, Tanaka R, Fujimori K. A Limonoid, 7-Deacetoxy-7-Oxogedunin (CG-1) from Andiroba (Carapa guianensis, Meliaceae) Lowers the Accumulation of Intracellular Lipids in Adipocytes via Suppression of IRS-1/Akt-Mediated Glucose Uptake and a Decrease in GLUT4 Expression. Molecules. 2019; 24(9):1668. https://doi.org/10.3390/molecules24091668
Chicago/Turabian StyleMatsumoto, Chihiro, Atsushi Koike, Reiko Tanaka, and Ko Fujimori. 2019. "A Limonoid, 7-Deacetoxy-7-Oxogedunin (CG-1) from Andiroba (Carapa guianensis, Meliaceae) Lowers the Accumulation of Intracellular Lipids in Adipocytes via Suppression of IRS-1/Akt-Mediated Glucose Uptake and a Decrease in GLUT4 Expression" Molecules 24, no. 9: 1668. https://doi.org/10.3390/molecules24091668
APA StyleMatsumoto, C., Koike, A., Tanaka, R., & Fujimori, K. (2019). A Limonoid, 7-Deacetoxy-7-Oxogedunin (CG-1) from Andiroba (Carapa guianensis, Meliaceae) Lowers the Accumulation of Intracellular Lipids in Adipocytes via Suppression of IRS-1/Akt-Mediated Glucose Uptake and a Decrease in GLUT4 Expression. Molecules, 24(9), 1668. https://doi.org/10.3390/molecules24091668