
Modulation of Endocannabinoid-Binding Receptors in Human Neuroblastoma Cells by Tunicamycin

 , , ,
, , , 
Abstract
:1. Introduction
2. Results
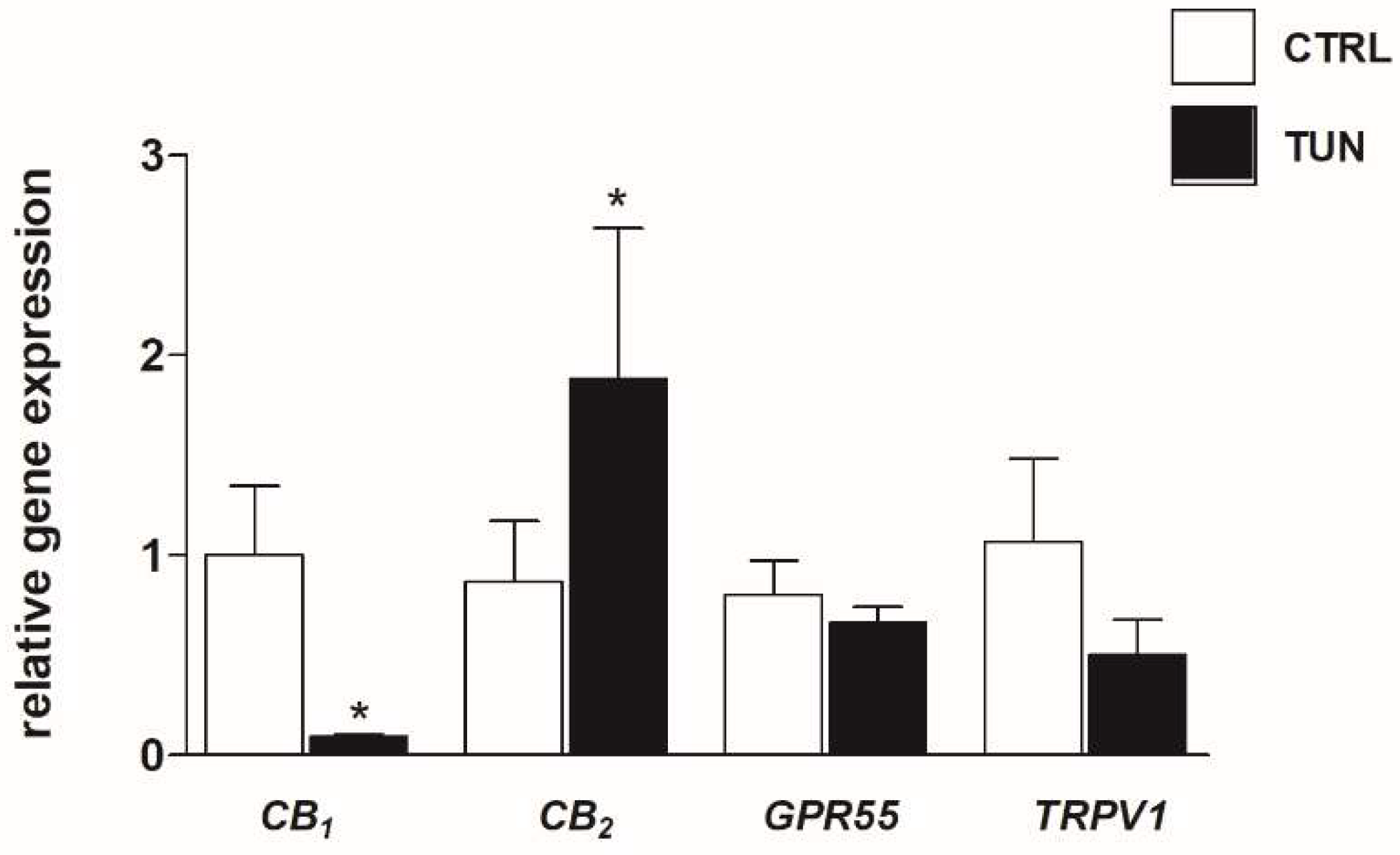
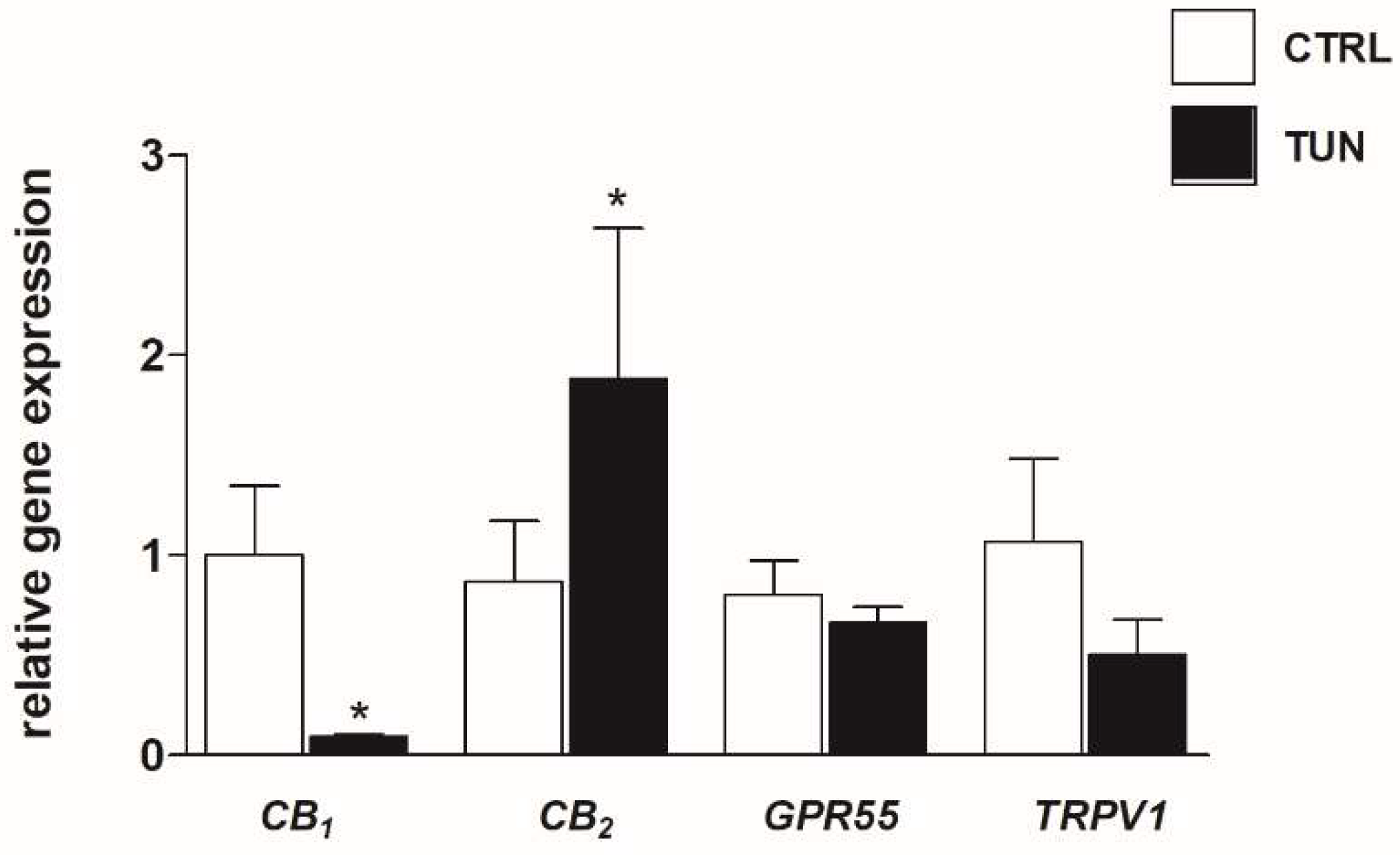
2.1. Effects of Tunicamycin on mRNA and Protein Expression of eCB-Binding Receptors
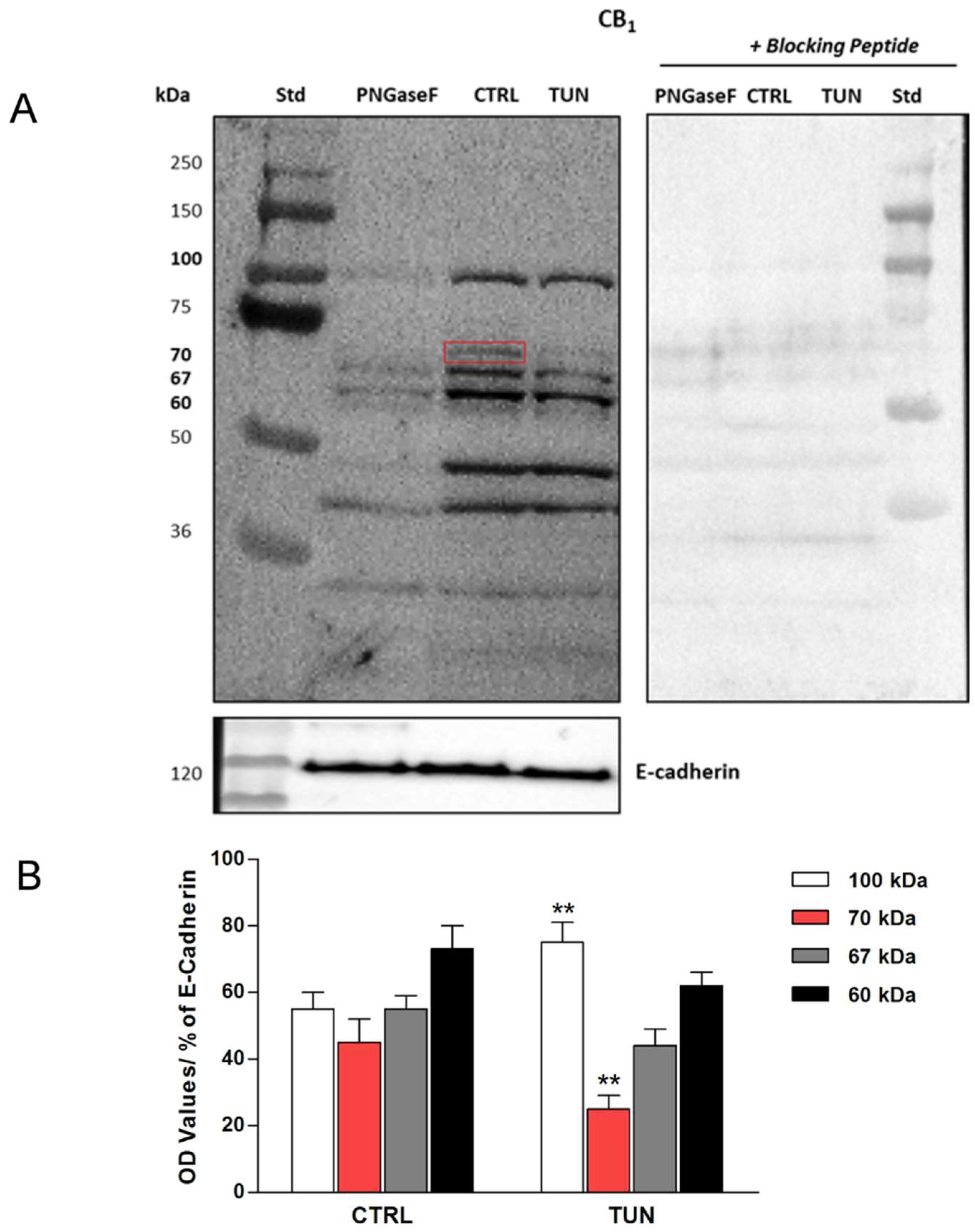
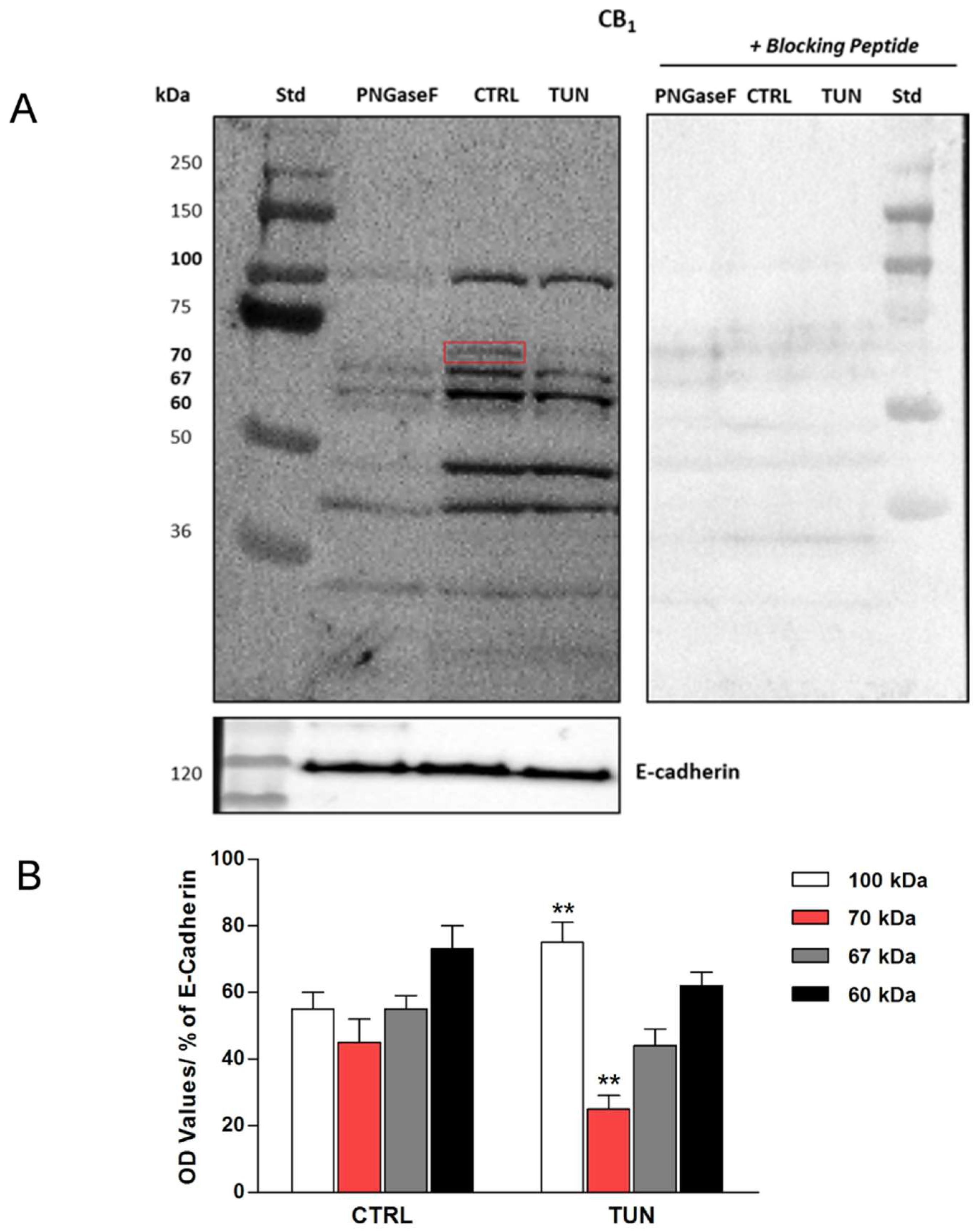
2.2. Assessment of N-linked Glycosylation after Tunicamycin Treatment
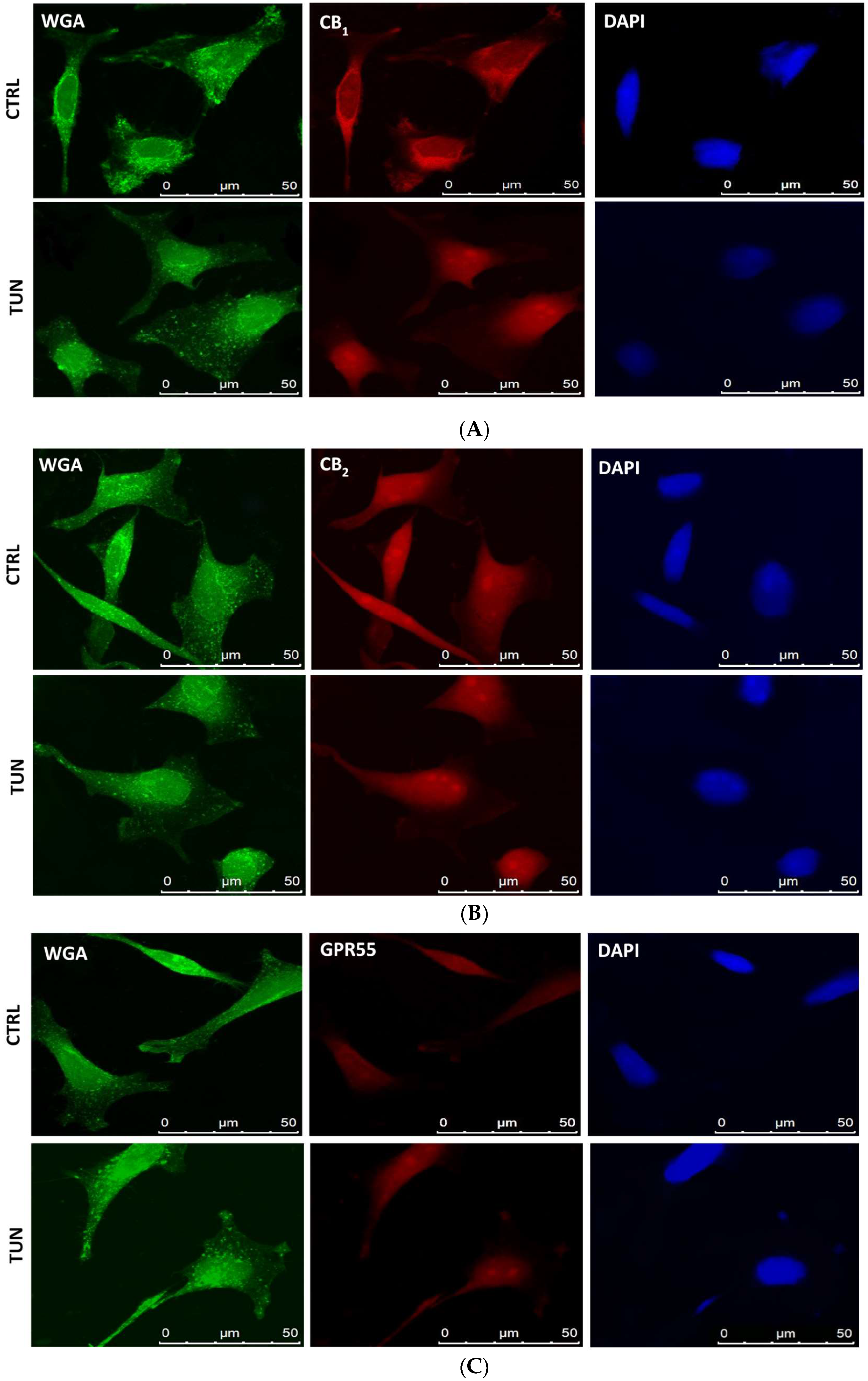
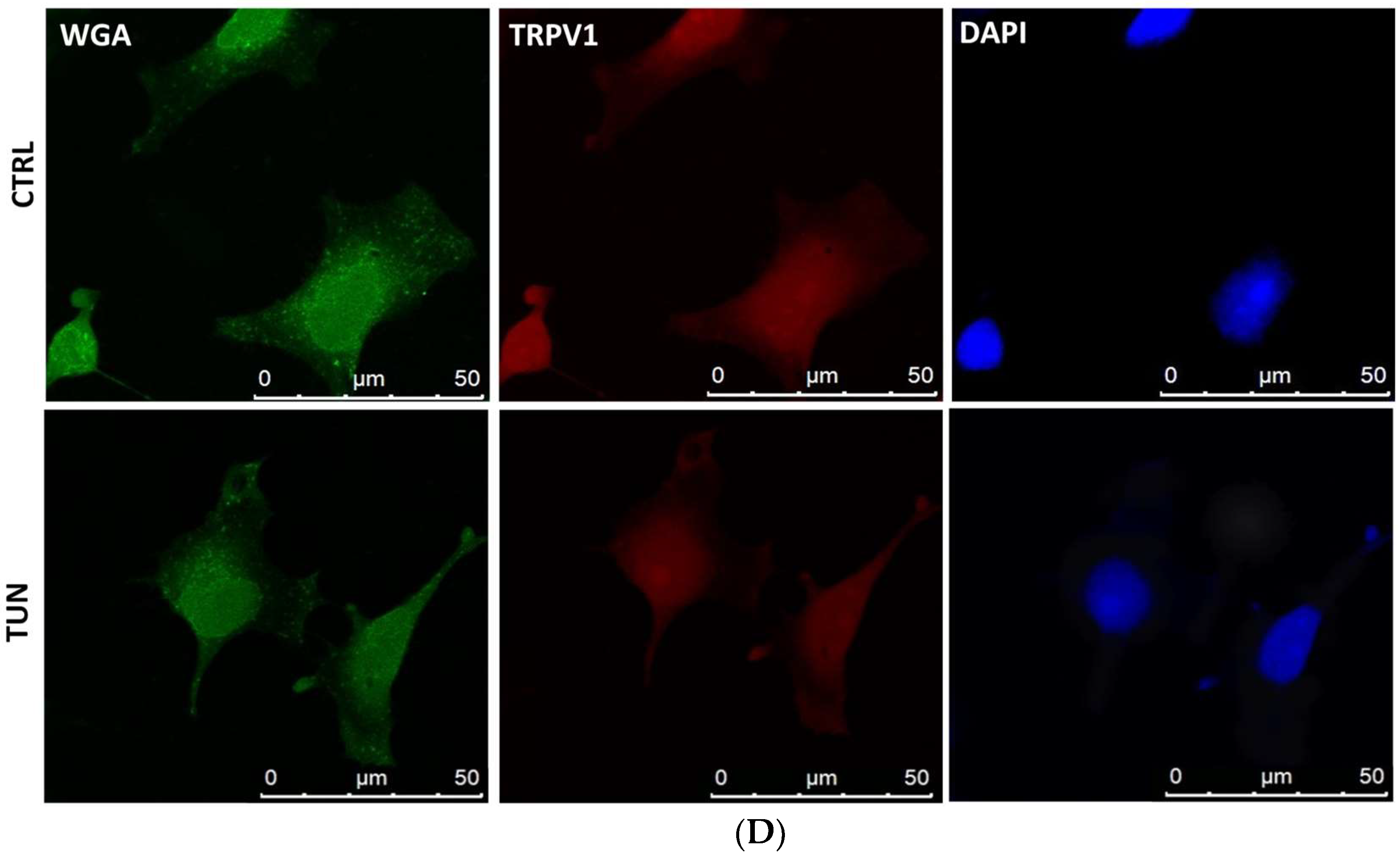
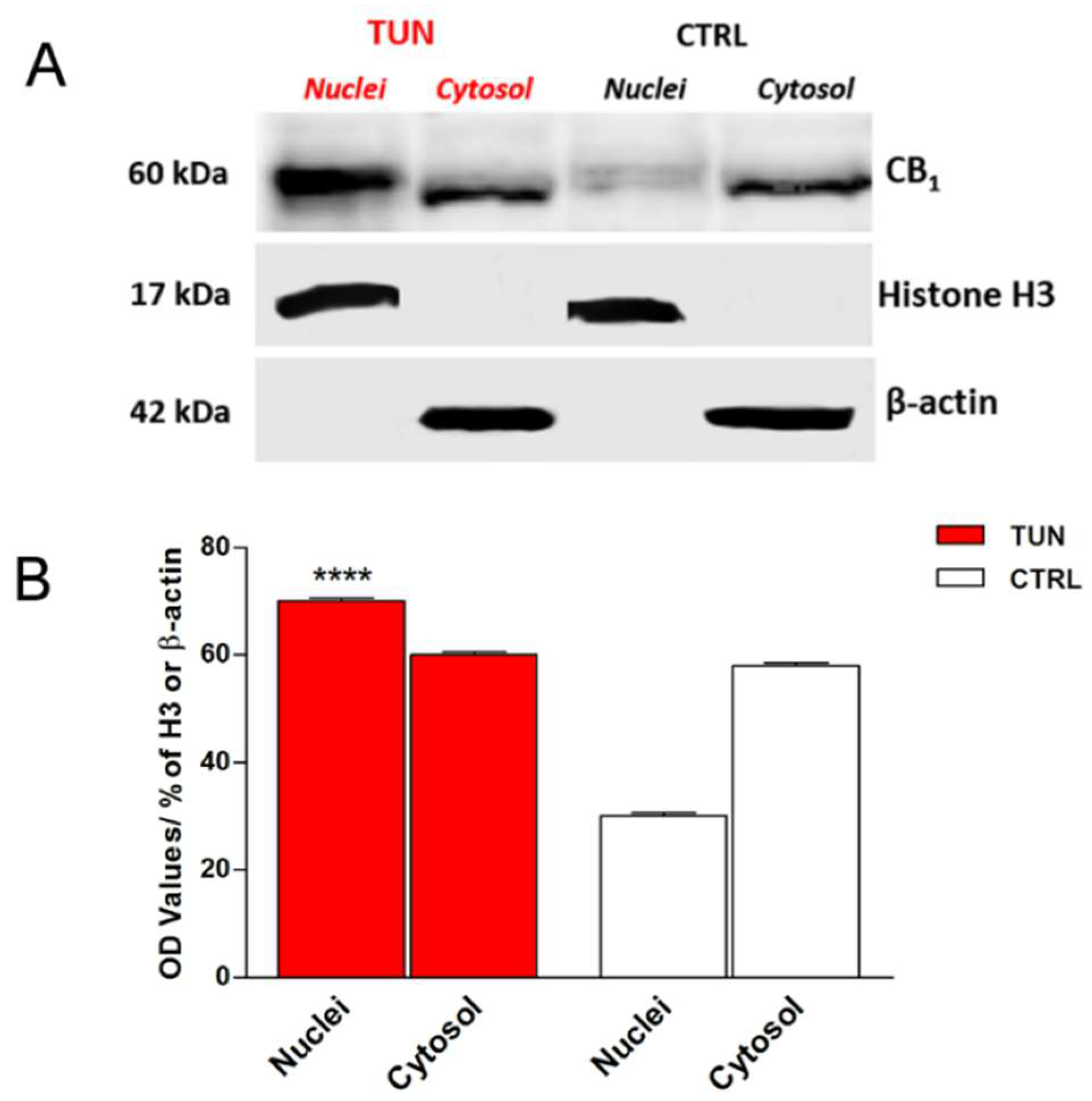
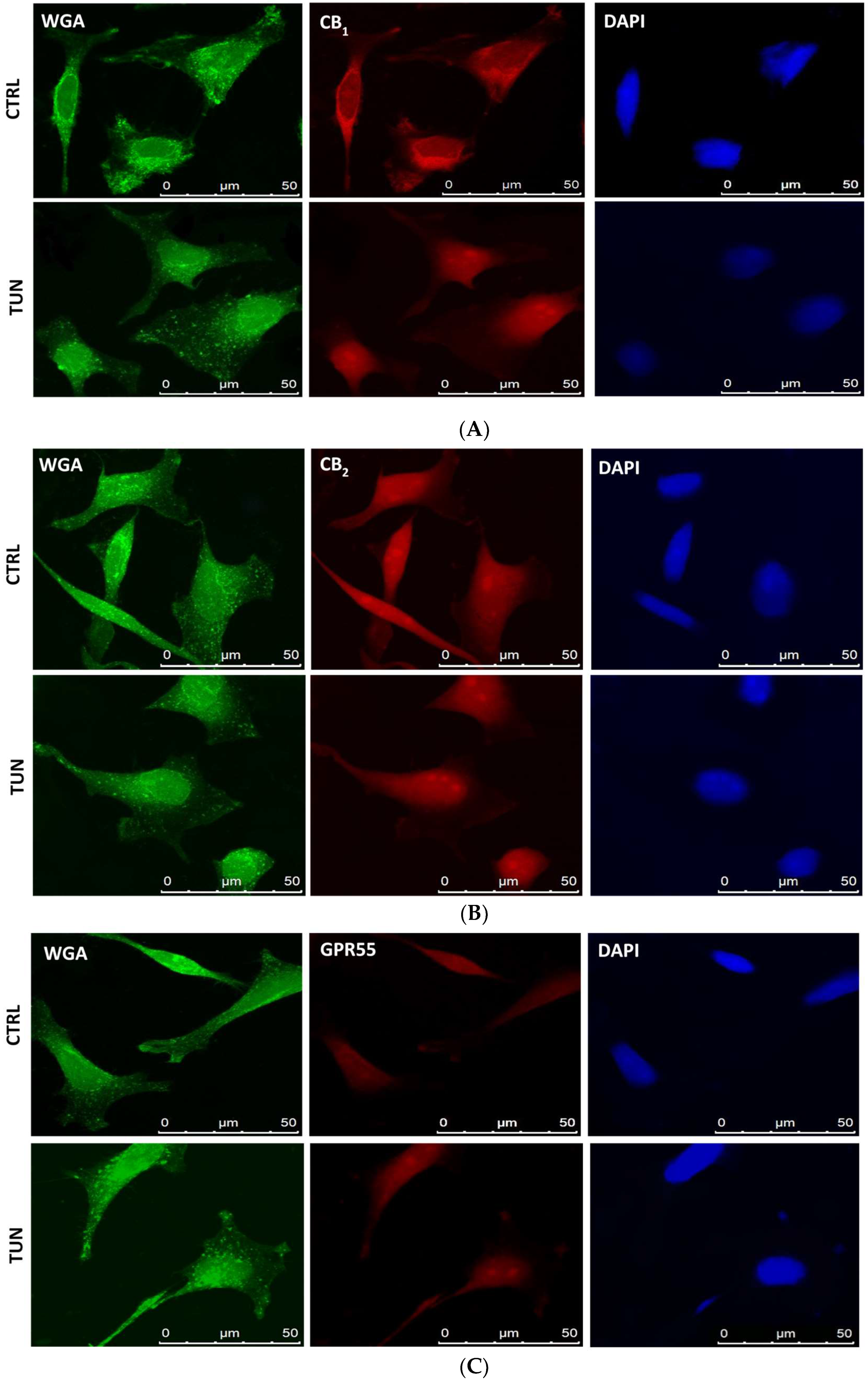
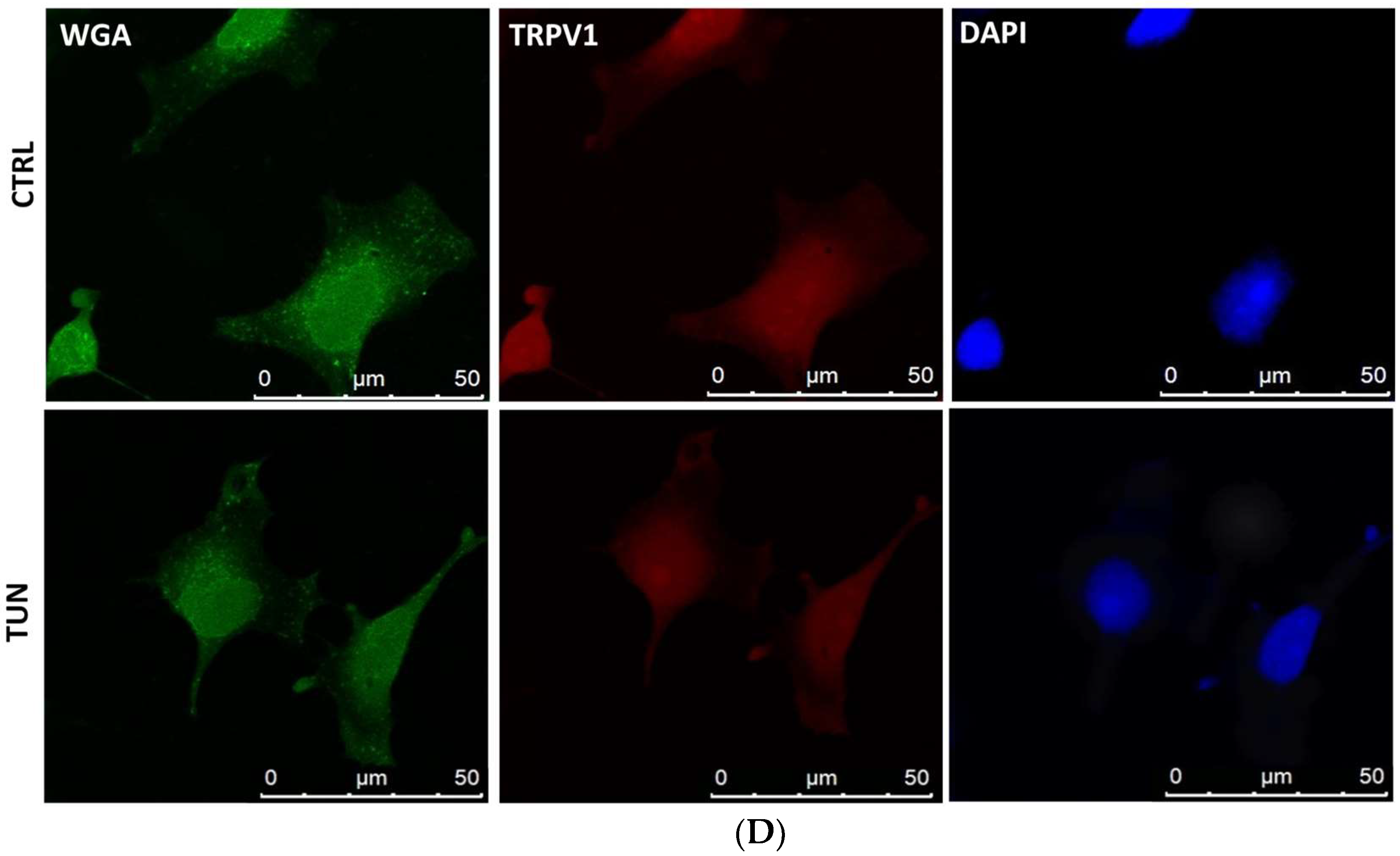
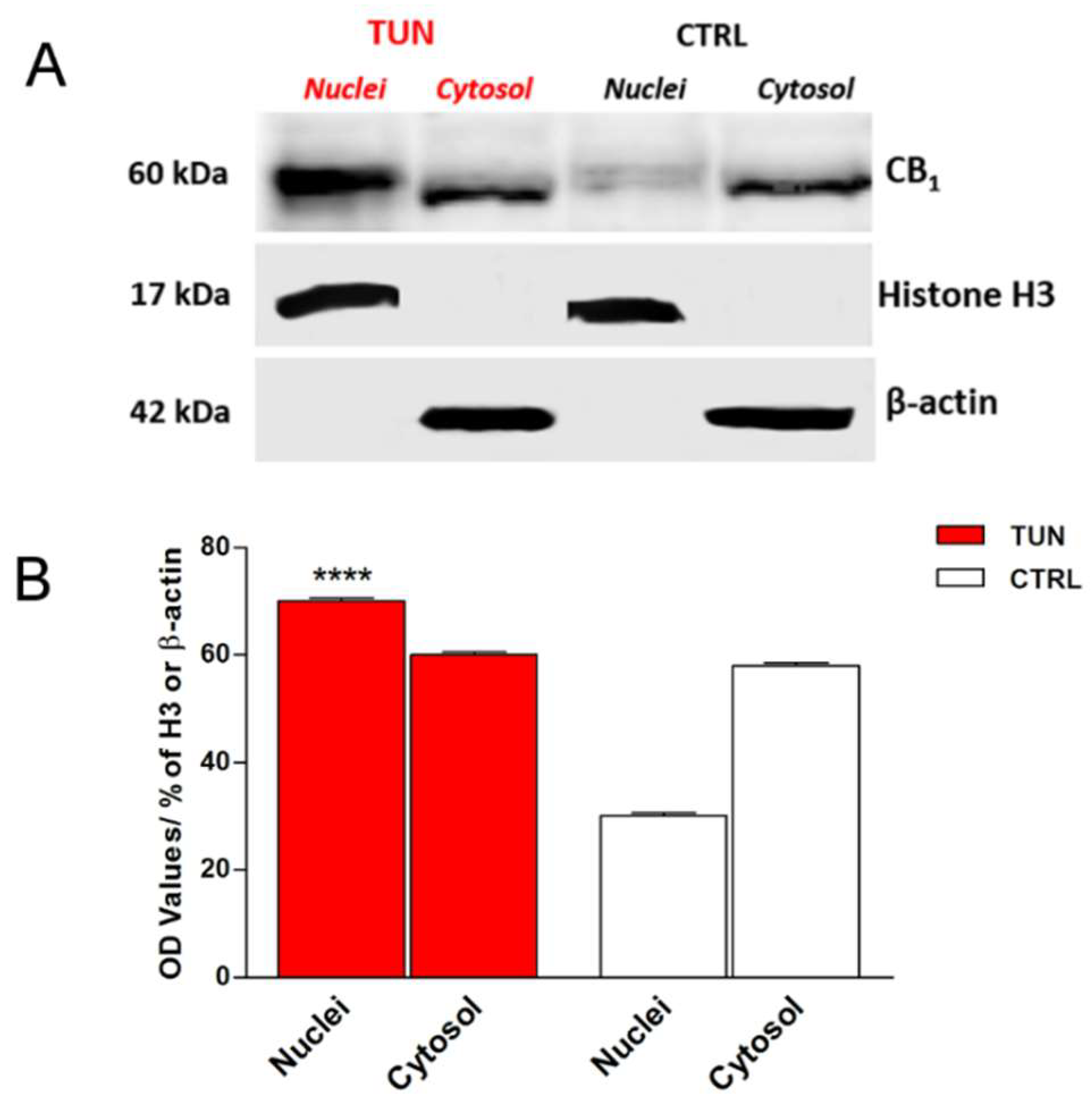
2.3. Localization of eCB-Binding Receptors in SH-SY5Y Cells upon Tunicamycin Treatment
2.4. Effects of Tunicamycin on CB ligand Binding
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Quantitative Real Time-Reverse Transcriptase-Polymerase Chain Reaction (qRT-PCR) Analysis
4.4. Fluorescence-Activated Cell Sorting (FACS) Analysis
4.5. Cellular Fractionation
4.6. Protein Deglycosylation Assay and Immunoblotting
4.7. Confocal Microscopy
4.8. Receptor Binding Assay on Adherent Living Cells
4.9. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pertwee, R.G. Endocannabinoids and Their Pharmacological Actions. Handb. Exp. Pharmacol. 2015, 231, 1–37. [Google Scholar] [CrossRef]
- Fezza, F.; Bari, M.; Florio, R.; Talamonti, E.; Feole, M.; Maccarrone, M. Endocannabinoids, related compounds and their metabolic routes. Molecules 2014, 19, 17078–17106. [Google Scholar] [CrossRef]
- Maccarrone, M.; Guzmán, M.; Mackie, K.; Doherty, P.; Harkany, T. Programming of neural cells by (endo)cannabinoids: From physiological rules to emerging therapies. Nat. Rev. Neurosci. 2014, 15, 786–801. [Google Scholar] [CrossRef]
- Maccarrone, M.; Bab, I.; Bíró, T.; Cabral, G.A.; Dey, S.K.; Di Marzo, V.; Konje, J.C.; Kunos, G.; Mechoulam, R.; Pacher, P.; et al. Endocannabinoid signaling at the periphery: 50 years after THC. Trends Pharmacol. Sci. 2015, 36, 277–296. [Google Scholar] [CrossRef] [Green Version]
- Ross, R.A. The enigmatic pharmacology of GPR55. Trends Pharmacol. Sci. 2009, 30, 156–163. [Google Scholar] [CrossRef]
- Di Marzo, V.; De Petrocellis, L. Endocannabinoids as regulators of transient receptor potential (TRP) channels: A further opportunity to develop new endocannabinoid-based therapeutic drugs. Curr. Med. Chem. 2010, 17, 1430–1449. [Google Scholar] [CrossRef]
- Maccarrone, M.; Battista, N.; Centonze, D. The endocannabinoid pathway in Huntington’s disease: A comparison with other neurodegenerative diseases. Prog. Neurobiol. 2007, 81, 349–379. [Google Scholar] [CrossRef]
- Bisogno, T.; Di Marzo, V. Cannabinoid receptors and endocannabinoids: Role in neuroinflammatory and neurodegenerative disorders. CNS Neurol. Disord. Drug Targets 2010, 9, 564–573. [Google Scholar] [CrossRef]
- Min, C.; Zheng, M.; Zhang, X.; Guo, S.; Kwon, K.J.; Shin, C.Y.; Kim, H.S.; Cheon, S.H.; Kim, K.M. N-linked Glycosylation on the N-terminus of the dopamine D2 and D3 receptors determines receptor association with specific microdomains in the plasma membrane. Biochim. Biophys. Acta 2015, 1853, 41–51. [Google Scholar] [CrossRef]
- He, J.; Xu, J.; Castleberry, A.M.; Lau, A.G.; Hall, R.A. Glycosylation of beta(1)-adrenergic receptors regulates receptor surface expression and dimerization. Biochem. Biophys. Res. Commun. 2002, 297, 565–572. [Google Scholar] [CrossRef]
- Wanamaker, C.P.; Green, W.N. N-linked glycosylation is required for nicotinic receptor assembly but not for subunit associations with calnexin. J. Biol. Chem. 2005, 280, 33800–33810. [Google Scholar] [CrossRef]
- Barrington, W.W.; Jacobson, K.A.; Stiles, G.L. Glycoprotein nature of the A2-adenosine receptor binding subunit. Mol. Pharmacol. 1990, 38, 177–183. [Google Scholar]
- Soto, A.G.; Smith, T.H.; Chen, B.; Bhattacharya, S.; Cordova, I.C.; Kenakin, T.; Vaidehi, N.; Trejo, J. N-linked glycosylation of protease-activated receptor-1 at extracellular loop 2 regulates G-protein signaling bias. Proc. Natl. Acad. Sci. USA 2015, 112, E3600–E3608. [Google Scholar] [CrossRef]
- Brockhausen, I. Pathways of O-glycan biosynthesis in cancer cells. Biochim. Biophys. Acta 1999, 1473, 67–95. [Google Scholar] [CrossRef]
- Dube, D.H.; Bertozzi, C.R. Glycans in cancer and inflammation—Potential for therapeutics and diagnostics. Nat. Rev. Drug Discov. 2005, 4, 477–488. [Google Scholar] [CrossRef]
- Packer, N.H.; von der Lieth, C.W.; Aoki-Kinoshita, K.F.; Lebrilla, C.B.; Paulson, J.C.; Raman, R.; Rudd, P.; Sasisekharan, R.; Taniguchi, N.; York, W.S. Frontiers in glycomics: Bioinformatics and biomarkers in disease. An Nih white paper prepared from discussions by the focus groups at a workshop on the Nih campus, Bethesda, MD (September 11–13, 2006). Proteomics 2008, 8, 8–20. [Google Scholar] [CrossRef]
- Helenius, A.; Aebi, M. Intracellular functions of N-linked glycans. Science 2001, 291, 2364–2369. [Google Scholar] [CrossRef]
- Hauselmann, I.; Borsig, L. Altered tumor-cell glycosylation promotes metastasis. Front. Oncol. 2014, 4, 28. [Google Scholar] [CrossRef]
- Berois, N.; Osinga, E. Glycobiology of neuroblastoma: Impact on tumor behavior, prognosis, and therapeutic stratergies. Front. Oncol. 2014, 4, 114. [Google Scholar] [CrossRef]
- Esko, J.D.; Bertozzi, C.R. Chemical tools for inhibiting glycosylation. In Essentials of Glycobiology, 2nd ed.; Varki, A., Cummings, R.D., Esko, J.D., Freeze, H.H., Stanley, P., Bertozzi, C.R., Hart, G.W., Etzler, M.E., Eds.; Cold Spring Harbor: New York, NY, USA, 2008; pp. 707–708. [Google Scholar]
- Oslowski, C.M.; Urano, F. Measuring ER stress and the unfolded protein response using mammalian tissue culture system. Methods Enzymol. 2011, 490, 71–92. [Google Scholar] [CrossRef] [Green Version]
- Guha, P.; Kaptan, E.; Gade, P.; Kalvakolanu, D.V.; Ahmed, H. Tunicamycin induced endoplasmic reticulum stress promotes apoptosis of prostate cancer cells by activating mTORC1. Oncotarget 2017, 15, 4068191–4068207. [Google Scholar] [CrossRef]
- Yan, K.; Khoshnoodi, J.; Ruotsalainen, V.; Tryggvason, K.J. N-linked glycosylation is critical for the plasma membrane localization of nephrin. Am. Soc. Nephrol. 2002, 13, 1385–1389. [Google Scholar] [CrossRef]
- Del Grosso, F.; De Mariano, M.; Passoni, L.; Luksch, R.; Tonini, G.P.; Longo, L. Inhibition of N-linked glycosylation impairs ALK phosphorylation and disrupts pro-survival signaling in neuroblastoma cell lines. BMC Cancer 2011, 11, 525. [Google Scholar] [CrossRef]
- Pasquariello, N.; Catanzaro, G.; Marzano, V.; Amadio, D.; Barcaroli, D.; Oddi, S.; Federici, G.; Urbani, A.; Finazzi Agrò, A.; Maccarrone, M. Characterization of the endocannabinoid system in human neuronal cells and proteomic analysis of anandamide-induced apoptosis. J. Biol. Chem. 2009, 23, 29413–29426. [Google Scholar] [CrossRef]
- Pertwee, R.G. Receptors and channels targeted by synthetic cannabinoid receptor agonists and antagonists. Curr. Med. Chem. 2010, 17, 1360–1381. [Google Scholar] [CrossRef]
- Onaivi, E.S.; Chakrabarti, A.; Chaudhuri, G. Cannabinoid receptor genes. Prog. Neurobiol. 1996, 48, 275–305. [Google Scholar] [CrossRef]
- Onaivi, E.S.; Leonard, C.M.; Ishiguro, H.; Zhang, P.W.; Lin, Z.; Akinshola, B.E.; Uhl, G.R. Endocannabinoids and cannabinoid receptor genetics. Prog. Neurobiol. 2002, 66, 307–344. [Google Scholar] [CrossRef]
- González-Mariscal, I.; Krzysik-Walker, S.M.; Doyle, M.E.; Liu, Q.R.; Cimbro, R.; Santa-Cruz Calvo, S.; Ghosh, S.; Cieśla, Ł.; Moaddel, R.; Carlson, O.D.; et al. Human CB1 Receptor Isoforms, present in Hepatocytes and β-cells, are Involved in Regulating Metabolism. Sci. Rep. 2016, 6, 33302. [Google Scholar] [CrossRef] [Green Version]
- Ruehle, S.; Wager-Miller, J.; Straiker, A.; Farnsworth, J.; Murphy, M.N.; Loch, S.; Monory, K.; Mackie, K.; Lutz, B. Discovery and characterization of two novel CB1 receptor splice variants with modified N-termini in mouse. J. Neurochem. 2017, 142, 521–533. [Google Scholar] [CrossRef]
- Howlett, A.C.; Champion-Dorow, T.M.; McMahon, L.L.; Westlake, T.M. The cannabinoid receptor: Biochemical and cellular properties in neuroblastoma cells. Pharmacol. Biochem. Behav. 1991, 40, 565–569. [Google Scholar] [CrossRef]
- Zhang, R.; Kim, T.K.; Qiao, Z.H.; Cai, J.; Pierce, W.M., Jr.; Song, Z.H. Biochemical and mass spectrometric characterization of the human CB2 cannabinoid receptor expressed in Pichia pastoris—Importance of correct processing of the N-terminus. Protein Expr. Purif. 2007, 55, 225–235. [Google Scholar] [CrossRef]
- Kedei, N.; Szabo, T.; Lile, J.D.; Treanor, J.J.; Olah, Z.; Iadarola, M.J.; Blumberg, P.M. Analysis of the native quaternary structure of vanilloid receptor 1. J. Biol. Chem. 2001, 276, 28613–28619. [Google Scholar] [CrossRef]
- Wirkner, K.; Hognestad, H.; Jahnel, R.; Hucho, F.; Illes, P. Characterization of rat transient receptor potential vanilloid 1 receptors lacking the N-glycosylation site N604. Neuroreport 2005, 16, 997–1001. [Google Scholar] [CrossRef]
- Veldhuis, N.A.; Lew, M.J.; Abogadie, F.C.; Poole, D.P.; Jennings, E.A.; Ivanusic, J.J.; Eilers, H.; Bunnett, N.W.; McIntyre, P. N-glycosylation determines ionic permeability and desensitization of the TRPV1 capsaicin receptor. J. Biol. Chem. 2012, 287, 21765–21772. [Google Scholar] [CrossRef]
- Free, R.B.; Hazelwood, L.A.; Cabrera, D.M.; Spalding, H.N.; Namkung, Y.; Rankin, M.L.; Sibley, D.R. D1 and D2 dopamine receptor expression is regulated by direct interaction with the chaperone protein calnexin. J. Biol. Chem. 2007, 282, 21285–21300. [Google Scholar] [CrossRef]
- Russo, D.; Chazenbalk, G.D.; Nagayama, Y.; Wadsworth, H.L.; Rapoport, B. Site-directed mutagenesis of the human thyrotropin receptor: Role of asparagine-linked oligosaccharides in the expression of a functional receptor. Mol. Endocrinol. 1991, 5, 29–33. [Google Scholar] [CrossRef]
- Chiang, N.; Tai, H.H. The role of N-glycosylation of human thromboxane A2 receptor in ligand binding. Arch. Biochem. Biophys. 1998, 352, 207–213. [Google Scholar] [CrossRef]
- Karpa, K.D.; Lidow, M.S.; Pickering, M.T.; Levenson, R.; Bergson, C. N-linked glycosylation is required for plasma membranelocalization of D5, but not D1, dopamine receptors in transfectedmammalian cells. Mol. Pharmacol. 1999, 56, 1071–1078. [Google Scholar] [CrossRef]
- Michineau, S.; Muller, L.; Pizard, A.; Alhenc-Gélas, F.; Rajerison, R.M. N-linked glycosylation of the human bradykinin B2 receptor is required for optimal cell-surface expression and coupling. Biol. Chem. 2004, 385, 49–57. [Google Scholar] [CrossRef]
- Lanctot, P.M.; Leclerc, P.C.; Clément, M.; Auger-Messier, M.; Escher, E.; Leduc, R.; Guillemette, G. Importance of N-glycosylation positioning for cell-surface expression, targeting, affinity and quality control of the human AT1 receptor. Biochem. J. 2005, 390, 367–376. [Google Scholar] [CrossRef]
- Dutton, A.C.; Massoura, A.N.; Dover, T.J.; Andrews, N.A.; Barnes, N.M. Identification and functional significance of N-glycosylation of the 5-ht5A receptor. Neurochem. Int. 2008, 52, 419–425. [Google Scholar] [CrossRef]
- Böer, U.; Neuschäfer-Rube, F.; Möller, U.; Püschel, G.P. Requirement of N-glycosylation of the prostaglandin E2 receptor EP3beta for correct sorting to the plasma membrane but not for correct folding. Biochem. J. 2000, 15, 839–847. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Inflammation and endoplasmic reticulum stress in obesity and diabetes. Int. J. Obes. (Lond.) 2008, 32, S52–S54. [Google Scholar] [CrossRef] [Green Version]
- Pacher, P.; Mechoulam, R. Is lipid signaling through cannabinoid 2 receptors part of a protective system? Prog. Lipid Res. 2011, 50, 193–211. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, H.; Yamashita, Y. Role of N-glycosylation in cell surface expression and protection against proteolysis of the intestinal anion exchanger SLC26A3. Am. J. Physiol. Cell Physiol. 2012, 302, C781–C795. [Google Scholar] [CrossRef]
- Pupo, M.; Bodmer, A.; Berto, M.; Maggiolini, M.; Dietrich, P.Y.; Picard, D. A genetic polymorphism repurposes the G-protein coupled and membrane-associated estrogen receptor GPER to a transcription factor-like molecule promoting paracrine signaling between stroma and breast carcinoma cells. Oncotarget 2017, 8, 46728–46744. [Google Scholar] [CrossRef] [Green Version]
- Oddi, S.; Dainese, E.; Sandiford, S.; Fezza, F.; Lanuti, M.; Chiurchiù, V.; Totaro, A.; Catanzaro, G.; Barcaroli, D.; De Laurenzi, V.; et al. Effects of palmitoylation of Cys(415) in helix 8 of the CB(1) cannabinoid receptor on membrane localization and signalling. Br. J. Pharmacol. 2012, 165, 2635–2651. [Google Scholar] [CrossRef]
- Oddi, S.; Dainese, E.; Fezza, F.; Lanuti, M.; Barcaroli, D.; De Laurenzi, V.; Centonze, D.; Maccarrone, M. Functional characterization of putative cholesterol binding sequence (CRAC) in human type-1 cannabinoid receptor. J. Neurochem. 2011, 116, 858–865. [Google Scholar] [CrossRef] [Green Version]
- Heakal, Y.; Woll, M.P.; Fox, T.; Seaton, K.; Levenson, R.; Kester, M. Neurotensin receptor-1 inducible palmitoylation is required for efficient receptor-mediated mitogenic-signaling within structured membrane microdomains. Cancer Biol. Ther. 2011, 12, 427–435. [Google Scholar] [CrossRef] [Green Version]
- Araque, A.; Castillo, P.E.; Manzoni, O.J.; Tonini, R. Synaptic functions of endocannabinoid signaling in health and disease. Neuropharmacology 2017, 124, 13–24. [Google Scholar] [CrossRef]
- Donvito, G.; Nass, S.R.; Wilkerson, J.L.; Curry, Z.A.; Schurman, L.D.; Kinsey, S.G.; Lichtman, A.H. The endogenous cannabinoid system: A budding source of targets for treating inflammatory and neuropathic pain. Neuropsychopharmacology 2018, 43, 52–79. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Dimauro, I.; Pearson, T.; Caporossi, D.; Jackson, M.J. A simple protocol for the subcellular fractionation of skeletal muscle and cells and tissue. BMC Res. Notes 2012, 5, 513. [Google Scholar] [CrossRef]
- Catani, V.M.; Gasperi, V. Assay of CB1 receptor binding. Methods Mol. Biol. 2016, 1412, 41–55. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds tunicamycin and PNGase F are available from the authors. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Group | CB1 | CB2 | GPR55 | TRPV1 | GlcNAc |
|---|---|---|---|---|---|
| CTRL | 70.60 ± 2.12 | 16.70 ± 0.50 | 30.00 ± 0.90 | 19.30 ± 0.60 | 146.00 ± 4.38 |
| TUN | 55.80 ± 1.67 * | 20.40 ± 0.61 | 22.60 ± 0.68 ** | 20.40 ± 0.61 | 131.40 ± 3.94 * |
| Receptor | UNIPROT Entry | Putative Glycosylated Asn Residues | Asn Position |
|---|---|---|---|
| CB1 | P21554 | 3 | 77, 83, 112 |
| CB2 | P34972 | 1 | 11 |
| GPR55 | Q9Y2T6 | 2 | 5, 171 |
| TRPV1 | Q8NER1 | 1 | 604 |
| Experimental Group | CB Binding Activity (pmol/mg of Protein) |
|---|---|
| CTRL | 17.40 ± 1.79 |
| TUN | 19.87 ± 0.36 |
| Human Gene | Forward (5′→3′) | Reverse (3′→5′) |
|---|---|---|
| CB1 | CCTTTTGCTGCCTAAATCCAC | CCACTGCTCAAACATCTGAC |
| CB2 | TCAACCCTGTCATCTATGCTC | AGTCAGTCCCAACACTCATC |
| GPR55 | ATCTACATGATCAACCTGGC | ATGAAGCAGATGGTGAAGACGC |
| TRPV1 | TCACCTACATCCTCCTGCTC | AAGTTCTTCCAGTGTCTGCC |
| β-Actin | TTCTACAATGAGCTGCGTG | AGAGGCGTACAGGGATAGCA |
| GAPDH | GATTCCACCCATGGCAAATTC | TGGGATTTCCATTGATGACAAG |
| 18S rRNA | CGCCGCTAGAGGTGAAATTCT | CGAACCTCCGACTTTCGTTCT |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rapino, C.; Castellucci, A.; Lizzi, A.R.; Sabatucci, A.; Angelucci, C.B.; Tortolani, D.; Rossi, G.; D’Andrea, G.; Maccarrone, M. Modulation of Endocannabinoid-Binding Receptors in Human Neuroblastoma Cells by Tunicamycin. Molecules 2019, 24, 1432. https://doi.org/10.3390/molecules24071432
Rapino C, Castellucci A, Lizzi AR, Sabatucci A, Angelucci CB, Tortolani D, Rossi G, D’Andrea G, Maccarrone M. Modulation of Endocannabinoid-Binding Receptors in Human Neuroblastoma Cells by Tunicamycin. Molecules. 2019; 24(7):1432. https://doi.org/10.3390/molecules24071432
Chicago/Turabian StyleRapino, Cinzia, Annalisa Castellucci, Anna Rita Lizzi, Annalaura Sabatucci, Clotilde B. Angelucci, Daniel Tortolani, Gianna Rossi, Gabriele D’Andrea, and Mauro Maccarrone. 2019. "Modulation of Endocannabinoid-Binding Receptors in Human Neuroblastoma Cells by Tunicamycin" Molecules 24, no. 7: 1432. https://doi.org/10.3390/molecules24071432
APA StyleRapino, C., Castellucci, A., Lizzi, A. R., Sabatucci, A., Angelucci, C. B., Tortolani, D., Rossi, G., D’Andrea, G., & Maccarrone, M. (2019). Modulation of Endocannabinoid-Binding Receptors in Human Neuroblastoma Cells by Tunicamycin. Molecules, 24(7), 1432. https://doi.org/10.3390/molecules24071432