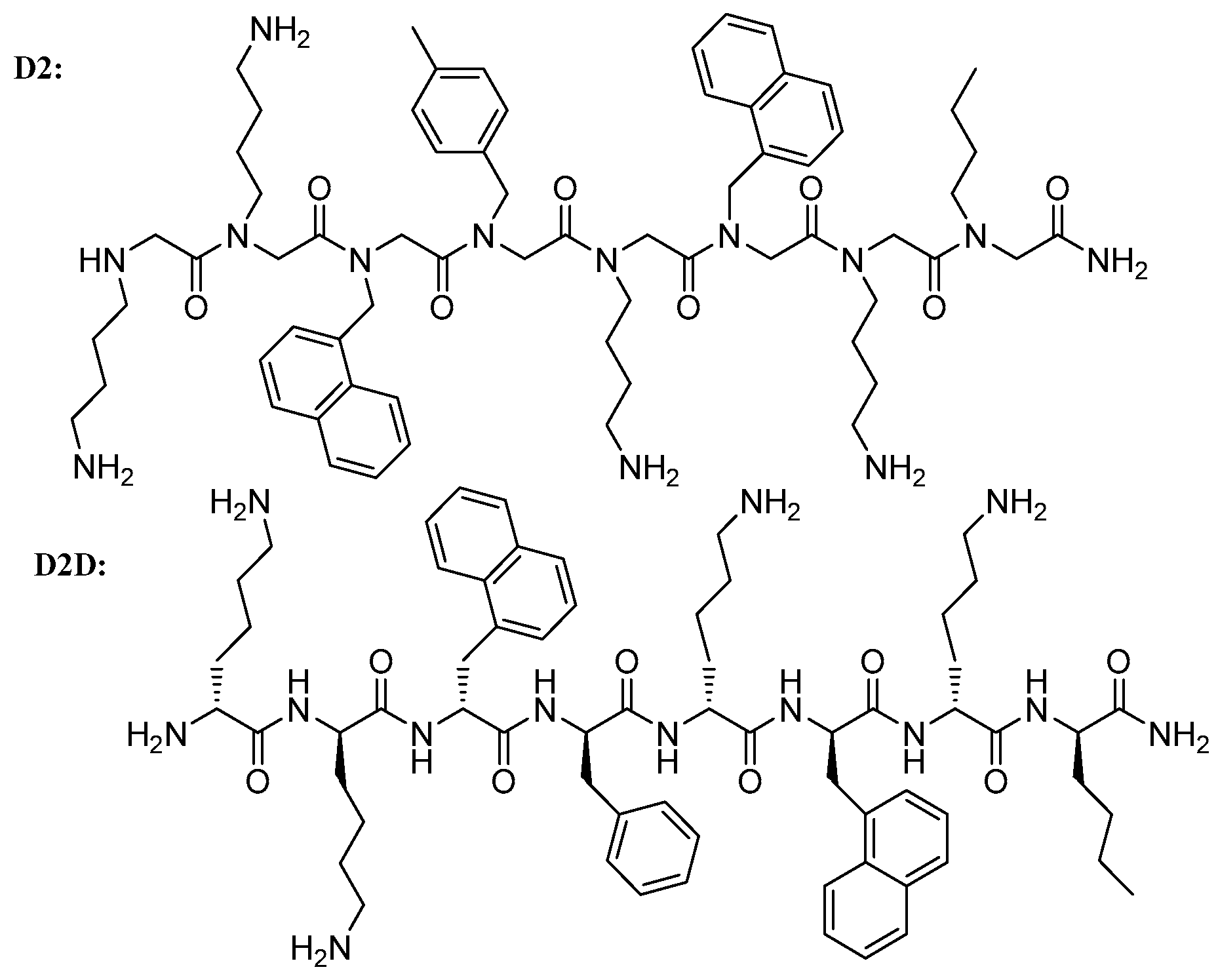
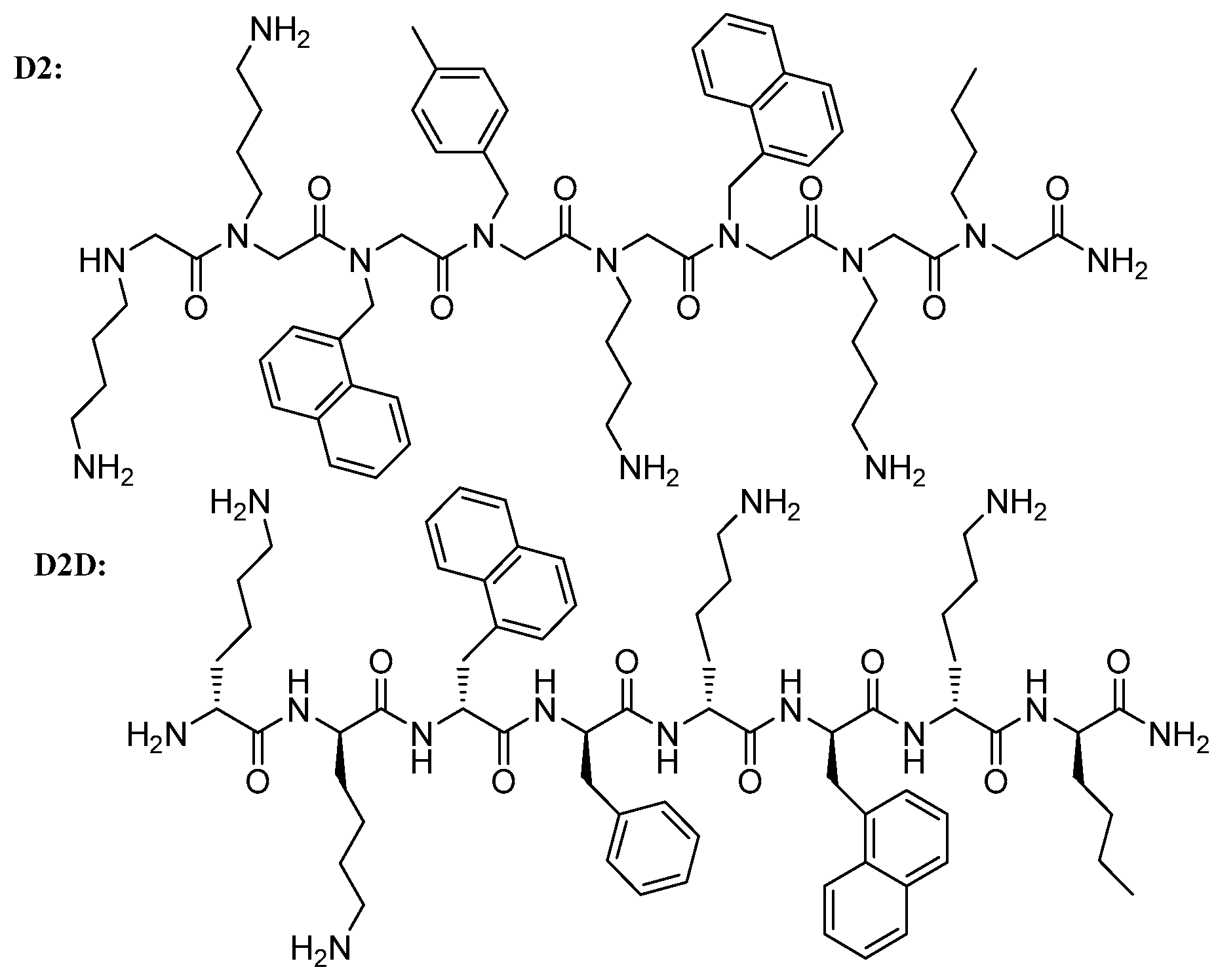
Structure–Activity Study, Characterization, and Mechanism of Action of an Antimicrobial Peptoid D2 and Its d- and l-Peptide Analogues

 , ,
, ,  , ,
, ,
Abstract
:
1. Introduction
2. Results and Discussion
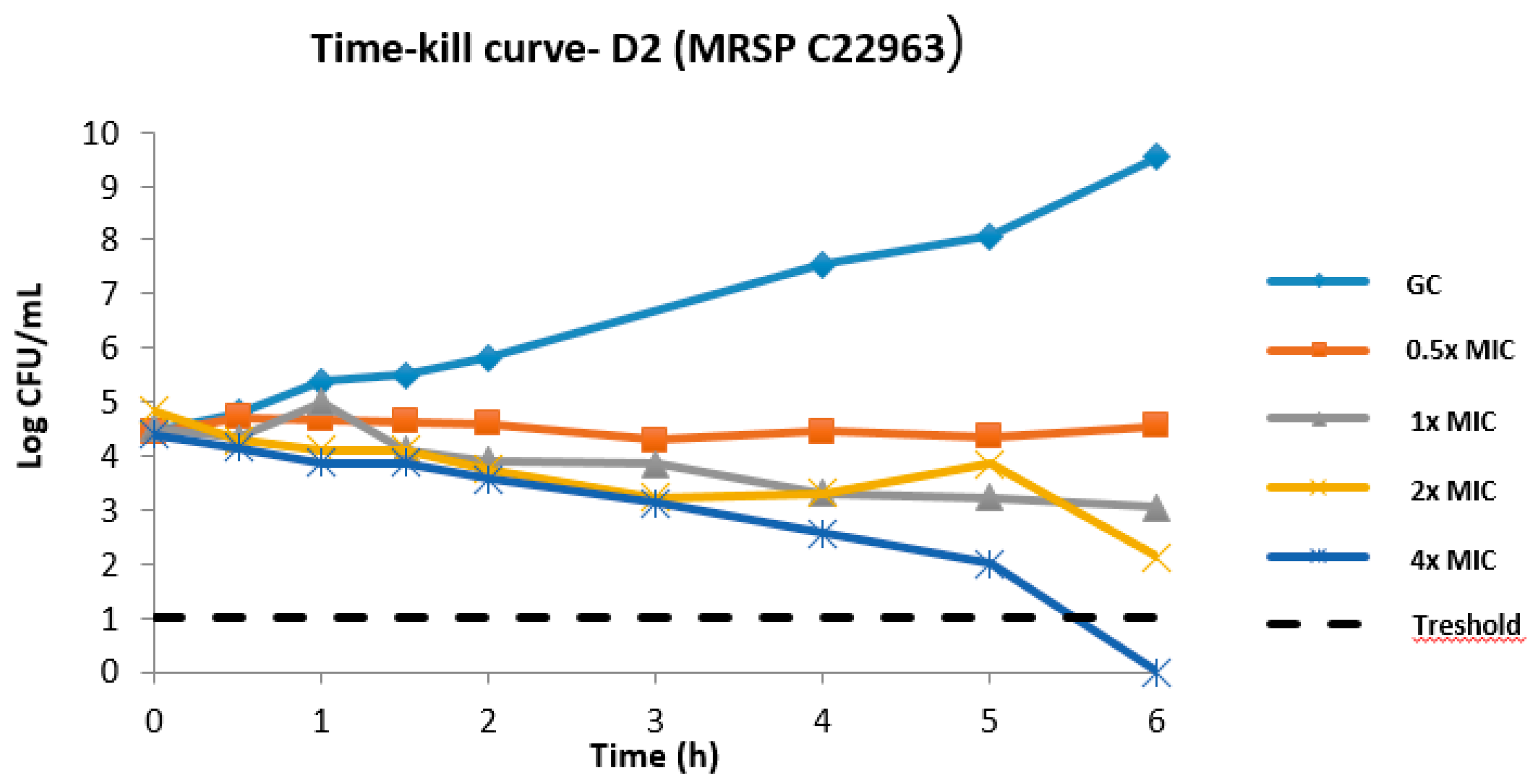
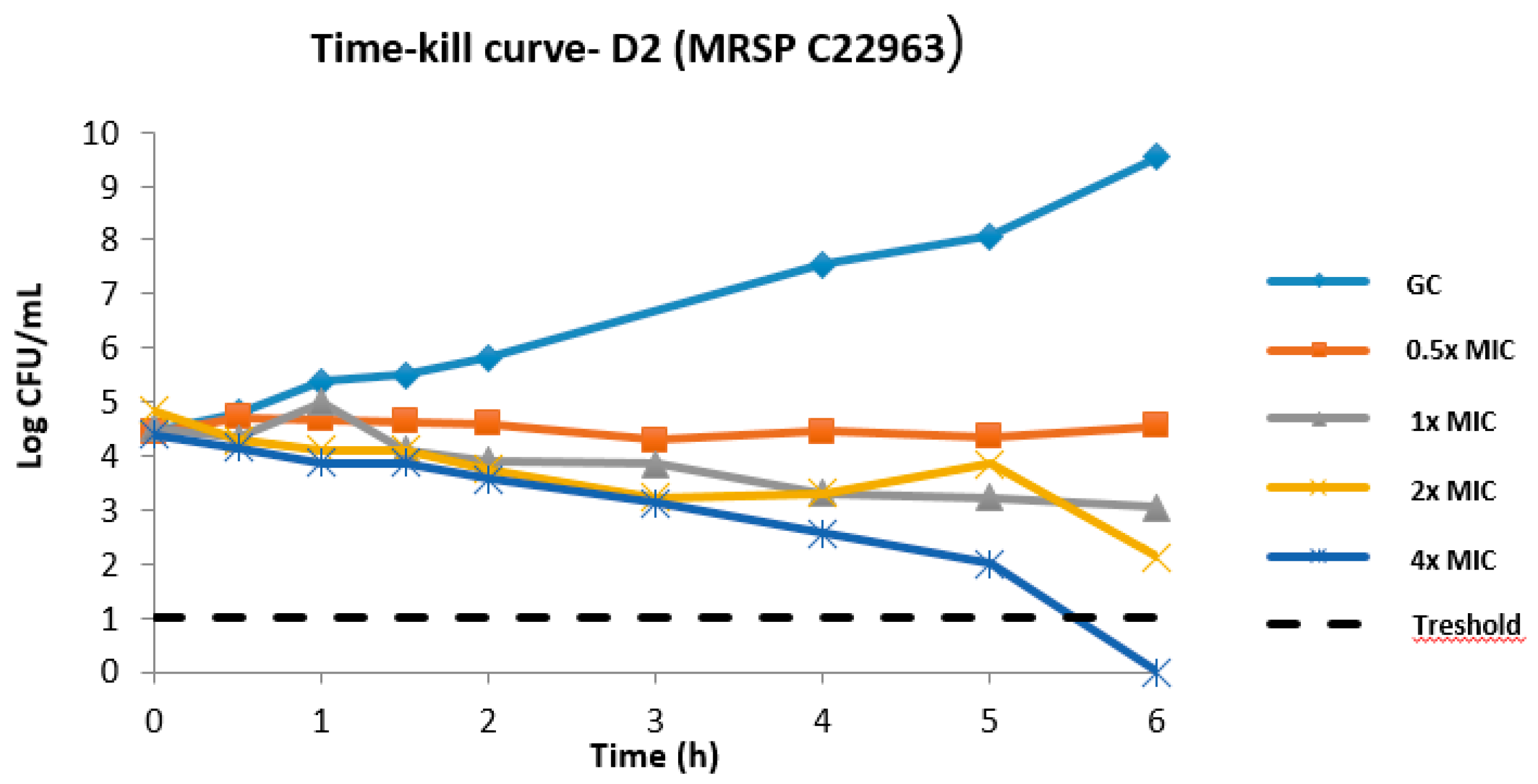
2.1. MIC Distribution of D2 and Time–Kill Kinetics
2.2. Structure–Activity Relationship (SAR) Study of D2
2.2.1. Glycine-Scan of D2
2.2.2. Substitutions in Position 5
2.3. Modification of Backbone Nature of D2
2.4. C- and N-Substituted Analogs
2.5. Antimicrobial Activity of the Best Analogue D2D
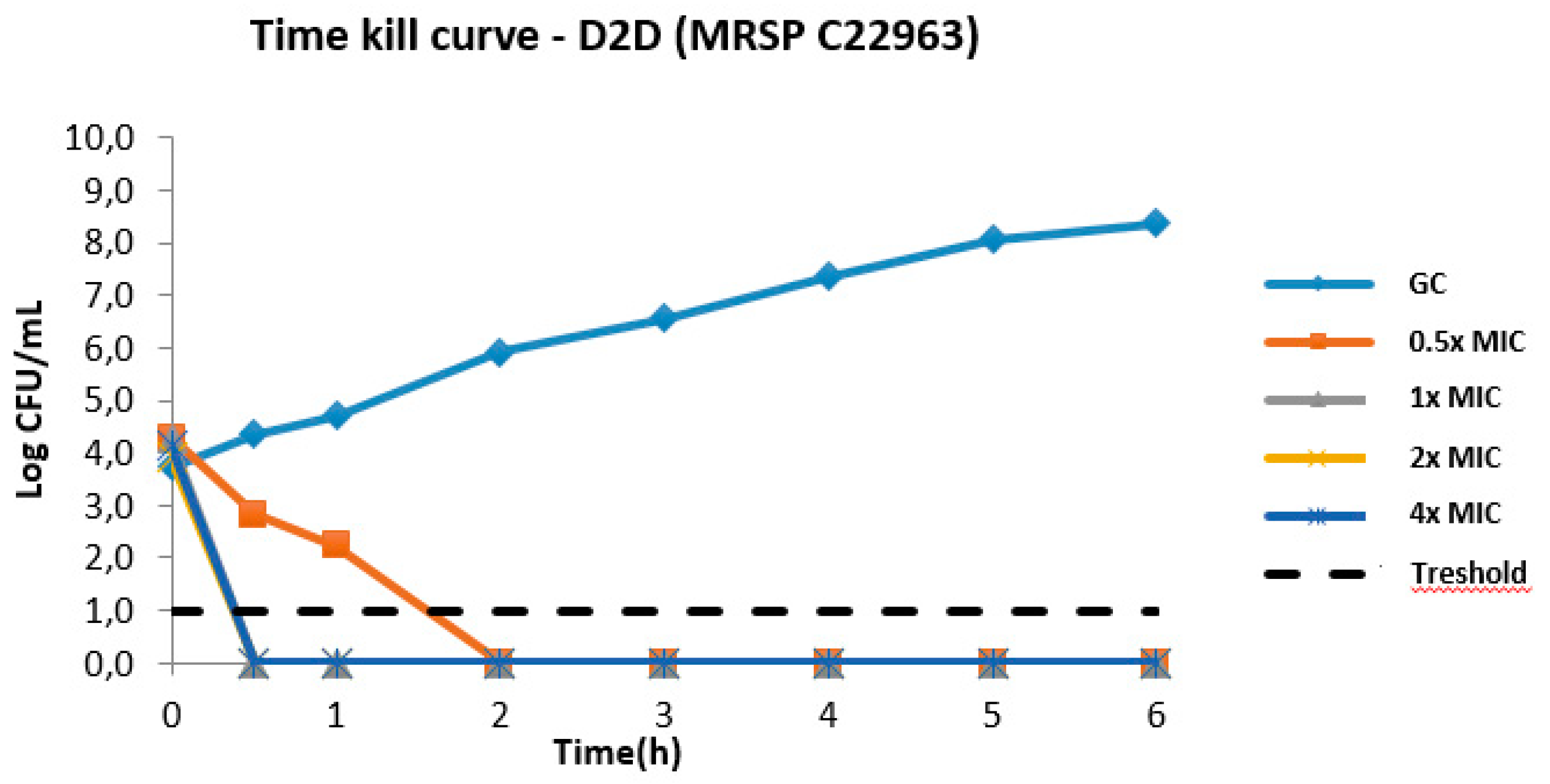
2.5.1. Time–Kill Kinetics of D2D
2.5.2. Antimicrobial Activity of D2 and D2D against Canine Pathogens
2.5.3. MSSP vs. MSSA
2.6. Circular Dichroism of Selected Analogues of D2
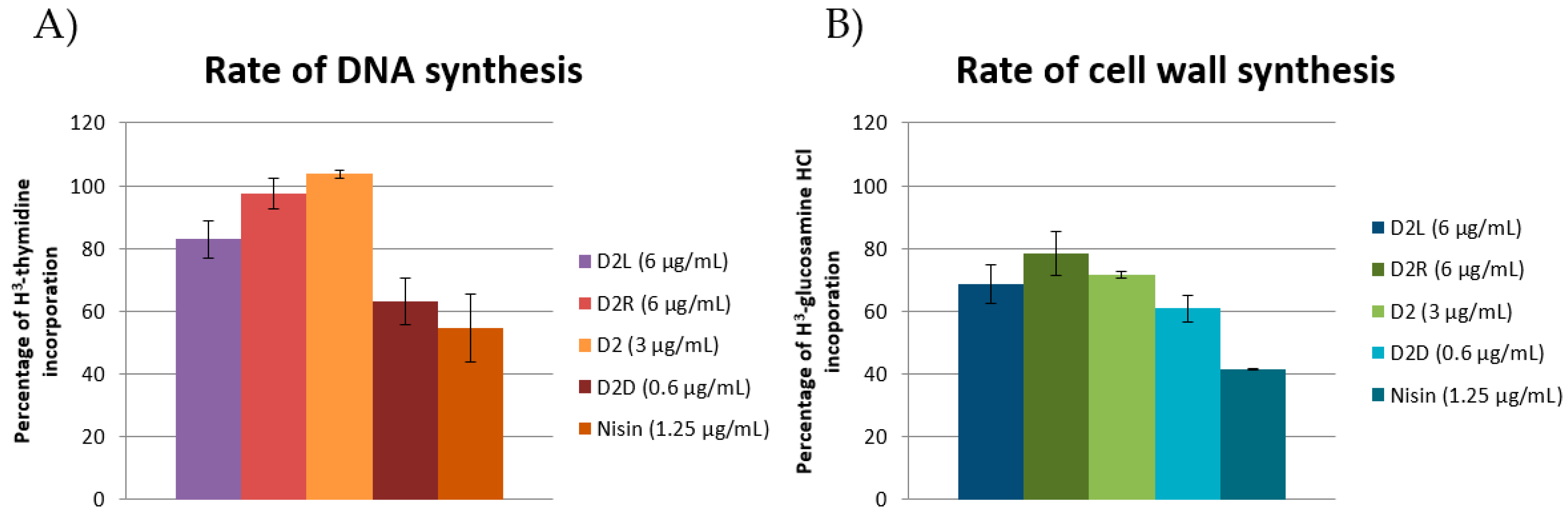
2.7. Investigating the Mechanism of Action by Emission DiSC3(5) Fluorescence and Macromolecule Biosynthesis Assay
2.7.1. Growth Curve Analysis
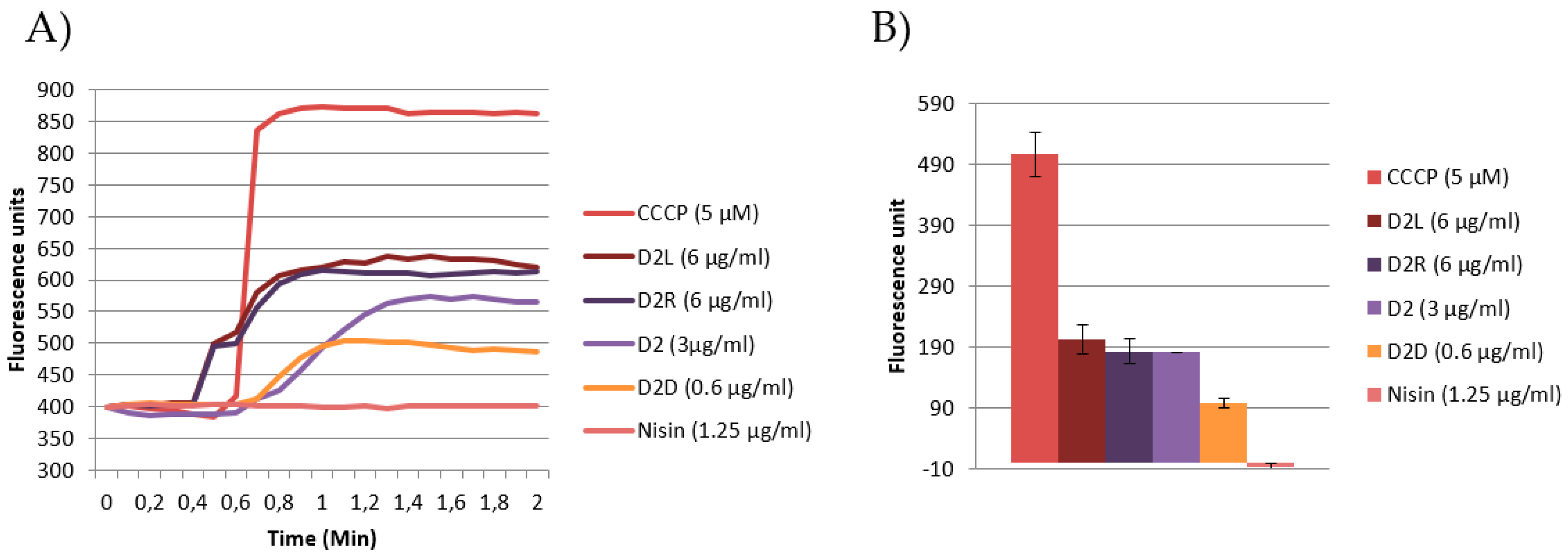
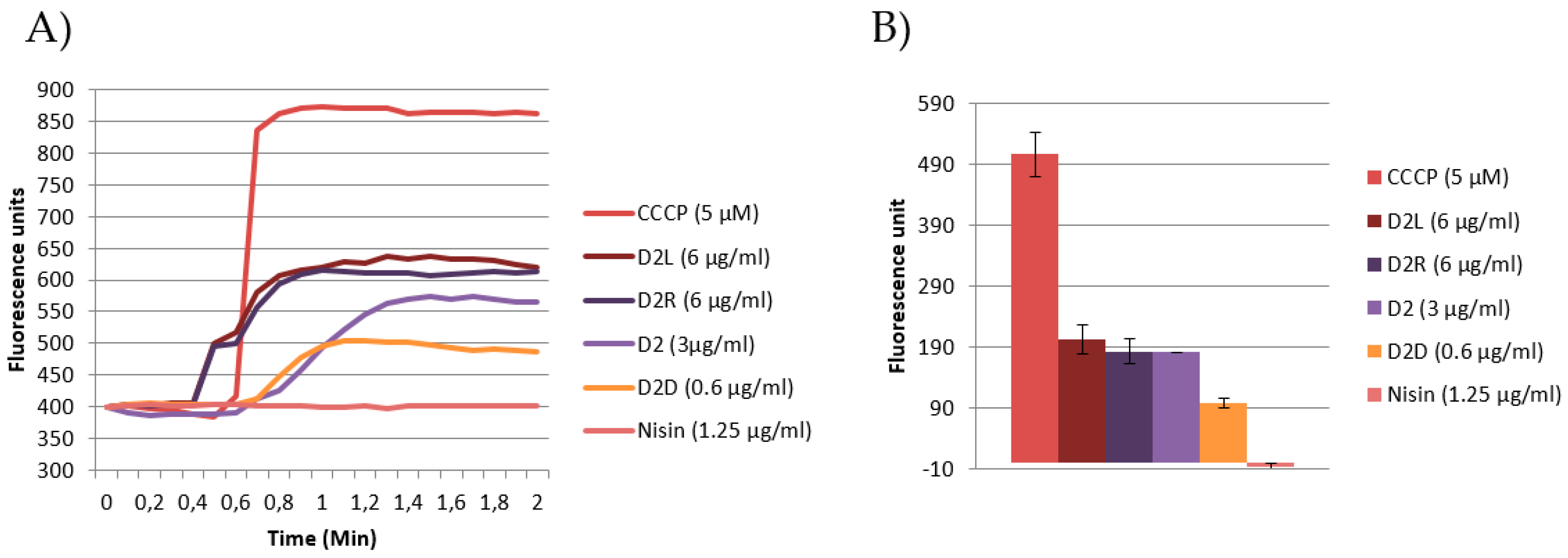
2.7.2. Emission DiSC3(5) Fluorescence Spectroscopy
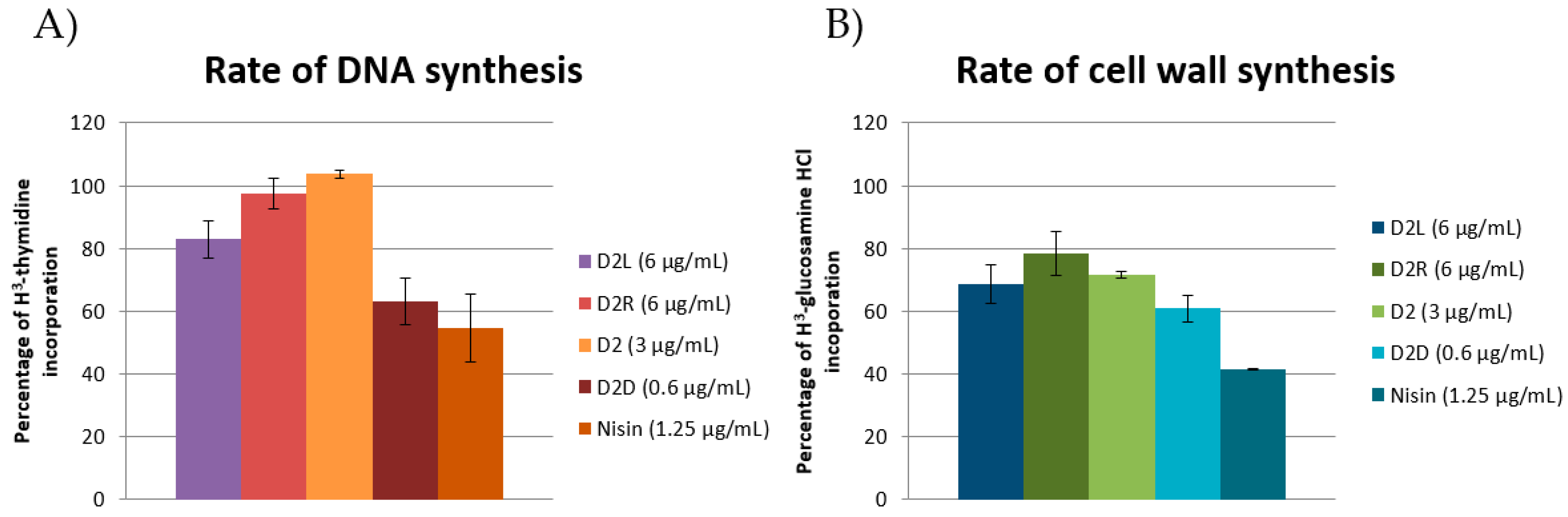
2.7.3. Macromolecule Biosynthesis Assay
3. Materials and Methods
3.1. Materials
3.2. Synthesis
3.3. Antimicrobial Susceptibility Testing
3.4. Time–Kill Kinetics
3.5. Hemolysis of Red Blood Cells
3.6. Growth Curves
Performance of Growth Curves in High-Throughput
3.7. Circular Dichroism (CD) Spectroscopy
3.8. Fluorescence Assay
Studying Bacterial Membrane Potential by Measuring DiSC3(5) Fluorescence
3.9. Macromolecule Biosynthesis Assay
Assay to Measure the Macromolecule Biosynthesis Rate
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Guardabassi, L.; Apley, M.; Olsen, J.E.; Toutain, P.-L.; Weese, S. Optimization of Antimicrobial Treatment to Minimize Resistance Selection. Microbiol. Spectrum 2018, 6. [Google Scholar] [CrossRef]
- Moodley, A.; Damborg, P.; Nielsen, S.S. Antimicrobial resistance in methicillin susceptible and methicillin resistant Staphylococcus pseudintermedius of canine origin: Literature review from 1980 to 2013. Vet. Microbiol. 2014, 171, 337–341. [Google Scholar] [CrossRef]
- dos Santos, T.P.; Damborg, P.; Moodley, A.; Guardabassi, L. Systematic Review on Global Epidemiology of Methicillin-Resistant Staphylococcus pseudintermedius: Inference of Population Structure from Multilocus Sequence Typing Data. Front. Microbiol. 2016, 7, 1599. [Google Scholar] [CrossRef]
- Stegmann, R.; Burnens, A.; Maranta, C.A.; Perreten, V. Human infection associated with methicillin-resistant Staphylococcus pseudintermedius ST71. J. Antimicrob. Chemother. 2010, 65, 2047–2048. [Google Scholar] [CrossRef]
- Hillier, A.; Alcorn, J.R.; Cole, L.K.; Kowalski, J.J. Pyoderma caused by Pseudomonas aeruginosa infection in dogs: 20 cases. Vet. Dermatol. 2006, 17, 432–439. [Google Scholar] [CrossRef]
- Starlander, G.; Borjesson, S.; Gronlund-Andersson, U.; Tellgren-Roth, C.; Melhusa, A. Cluster of Infections Caused by Methicillin-Resistant Staphylococcus pseudintermedius in Humans in a Tertiary Hospital. J. Clin. Microbiol. 2014, 52, 3118–3120. [Google Scholar] [CrossRef]
- Haney, E.F.; Mansour, S.C.; Hancock, R.E.W. Antimicrobial Peptides: An Introduction. In Antimicrobial Peptides: Methods and Protocols; Hansen, P.R., Ed.; Springer New York: New York, NY, USA, 2017; pp. 3–22. [Google Scholar]
- Matsuzaki, K. Control of cell selectivity of antimicrobial peptides. Biochim. Biophys. Acta (BBA) Biomembr. 2009, 1788, 1687–1692. [Google Scholar] [CrossRef]
- Wimley, W.C. Describing the Mechanism of Antimicrobial Peptide Action with the Interfacial Activity Model. ACS Chem. Biol. 2010, 5, 905–917. [Google Scholar] [CrossRef]
- Mahlapuu, M.; Hakansson, J.; Ringstad, L.; Bjorn, C. Antimicrobial Peptides: An Emerging Category of Therapeutic Agents. Front. Cell. Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef]
- Molchanova, N.; Hansen, P.R.; Franzyk, H. Advances in Development of Antimicrobial Peptidomimetics as Potential Drugs. Molecules 2017, 22, 1430. [Google Scholar] [CrossRef]
- Zuckermann, R.N.; Kodadek, T. Peptoids as potential therapeutics. Curr. Opin. Mol. Ther. 2009, 11, 299–307. [Google Scholar]
- Bolt, H.L.; Eggimann, G.A.; Jahoda, C.A.B.; Zuckermann, R.N.; Sharples, G.J.; Cobb, S.L. Exploring the links between peptoid antibacterial activity and toxicity. MedChemComm 2017, 8, 886–896. [Google Scholar] [CrossRef]
- Ryge, T.S.; Hansen, P.R. Potent antibacterial lysine-peptoid hybrids identified from a positional scanning combinatorial library. Bioorg. Med. Chem. 2006, 14, 4444–4451. [Google Scholar] [CrossRef]
- Molchanova, N.; Hansen, P.R.; Damborg, P.; Nielsen, H.M.; Franzyk, H. Lysine-Based α-peptide/β-peptoid peptidomimetics: Influence of hydrophobicity, fluorination and distribution of cationic charge on antimicrobial activity and cytotoxicity. ChemMedChem 2017, 20, 312–318. [Google Scholar] [CrossRef]
- Jahnsen, R.D.; Sandberg-Schaal, A.; Vissing, K.J.; Nielsen, H.M.; Frimodt-Møller, N.; Franzyk, H. Tailoring Cytotoxicity of Antimicrobial Peptidomimetics with High Activity against Multidrug-Resistant Escherichia coli. J. Med. Chem. 2014, 57, 2864–2873. [Google Scholar] [CrossRef]
- Henninot, A.; Collins, J.C.; Nuss, J.M. The Current State of Peptide Drug Discovery: Back to the Future? J. Med. Chem. 2017, 61, 1382–1414. [Google Scholar] [CrossRef]
- Greco, I.; Hummel, B.; Vasir, J.; Watts, J.; Koch, J.; Hansen, J.; Nielsen, H.; Damborg, P.; Hansen, P. In Vitro ADME Properties of Two Novel Antimicrobial Peptoid-Based Compounds as Potential Agents against Canine Pyoderma. Molecules 2018, 23, 630. [Google Scholar] [CrossRef] [PubMed]
- Molchanova, N.; Hansen, P.; Damborg, P.; Franzyk, H. Fluorinated antimicrobial lysine-based peptidomimetics with activity against methicillin-resistant Staphylococcus pseudintermedius. J. Peptide Sci. 2018, 24, e3098. [Google Scholar] [CrossRef]
- Mohamed, M.F.; Hammac, G.K.; Guptill, L.; Seleem, M.N. Antibacterial Activity of Novel Cationic Peptides against Clinical Isolates of Multi-Drug Resistant Staphylococcus pseudintermedius from Infected Dogs. PLoS ONE 2014, 9, e116259. [Google Scholar] [CrossRef] [PubMed]
- Greco, I.; Emborg, A.; Jana, B.; Molchanova, N.; Oddo, A.; Damborg, P.; Guardabassi, L.; Hansen, P.R. Characterization, mechanism of action and optimization of activity of a novel antimicrobial peptide-peptoid hybrid against bacterial pathogens involved in canine skin infections. Sci. Rep. 2019, 9, 3679. [Google Scholar] [CrossRef]
- Cabassi, C.S.; Sala, A.; Santospirito, D.; Alborali, G.L.; Carretto, E.; Ghibaudo, G.; Taddei, S. Activity of AMP2041 against human and animal multidrug resistant Pseudomonas aeruginosa clinical isolates. Ann. Clin. Microbiol. Antimicrob. 2017, 16, 17. [Google Scholar] [CrossRef]
- Gerling, U.I.M.; Brandenburg, E.; Berlepsch, H.V.; Pagel, K.; Koksch, B. Structure Analysis of an Amyloid-Forming Model Peptide by a Systematic Glycine and Proline Scan. Biomacromolecules 2011, 12, 2988–2996. [Google Scholar] [CrossRef] [PubMed]
- Oddo, A.; Hansen, P.R. Hemolytic Activity of Antimicrobial Peptides. Methods Mol Biol. 2017, 1548, 427–435. [Google Scholar] [PubMed]
- Oren, Z.; Hong, J.; Shai, Y. A comparative study on the structure and function of a cytolytic -helical peptide and its antimicrobial-sheet diastereomer. Eur. J. Biochem. 1999, 259, 360–369. [Google Scholar] [CrossRef] [PubMed]
- Aalbaek, B.; Bemis, D.A.; Schjaerff, M.; Kania, S.A.; Frank, L.A.; Guardabassi, L. Coryneform bacteria associated with canine otitis externa. Vet. Microbiol 2010, 145, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, C.L.; Charlebois, A.; Masson, L.; Archambault, M. Characterization of hospital-associated lineages of ampicillin-resistant Enterococcus faecium from clinical cases in dogs and humans. Front. Microbiol. 2013, 4, 245. [Google Scholar] [CrossRef] [PubMed]
- Malisova, B.; Santavy, P.; Loveckova, Y.; Hladky, B.; Kotaskova, I.; Pol, J.; Lonsky, V.; Nemec, P.; Freiberger, T. Human native endocarditis caused by Streptococcus canis-a case report. Apmis 2019, 127, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Harada, K.; Niina, A.; Shimizu, T.; Mukai, Y.; Kuwajima, K.; Miyamoto, T.; Kataoka, Y. Phenotypic and molecular characterization of antimicrobial resistance in Proteus mirabilis isolates from dogs. J. Med. Microbiol. 2014, 63, 1561–1567. [Google Scholar] [CrossRef]
- Sisido, M.; Egusa, S.; Imanishi, Y. One-dimensional aromatic crystals in solution. 2. Synthesis, conformation, and spectroscopic properties of poly(L-2-naphthylalanine). J. Am. Chem. Soc. 1983, 105, 4077–4082. [Google Scholar] [CrossRef]
- Huang, H.W. Molecular mechanism of antimicrobial peptides: The origin of cooperativity. Biochim. Biophys. Acta (BBA) Biomembr. 2006, 1758, 1292–1302. [Google Scholar] [CrossRef]
- Matsuzaki, K.; Murase, O.; Fujii, N.; Miyajima, K. An Antimicrobial Peptide, Magainin 2, Induced Rapid Flip-Flop of Phospholipids Coupled with Pore Formation and Peptide Translocation. Biochemistry 1996, 35, 11361–11368. [Google Scholar] [CrossRef] [PubMed]
- Le, C.F.; Fang, C.M.; Sekaran, S.D. Intracellular Targeting Mechanisms by Antimicrobial Peptides. Antimicrob. Agents Chemother. 2017, 61, e02340-16. [Google Scholar] [CrossRef] [PubMed]
- Marcellini, L.; Giammatteo, M.; Aimola, P.; Mangoni, M.L. Fluorescence and Electron Microscopy Methods for Exploring Antimicrobial Peptides Mode(s) of Action. In Antimicrobial Peptides: Methods and Protocols; Giuliani, A., Rinaldi, A.C., Eds.; Humana Press: Totowa, NJ, USA, 2010; pp. 249–266. [Google Scholar]
- Jana, B.; Baker, K.R.; Guardabassi, L. Macromolecule Biosynthesis Assay and Fluorescence Spectroscopy Methods to Explore Antimicrobial Peptide Mode(s) of Action. In Antimicrobial Peptides: Methods and Protocols; Hansen, P.R., Ed.; Springer New York: New York, NY, USA, 2017; pp. 181–190. [Google Scholar]
- Brochmann, R.P.; Helmfrid, A.; Jana, B.; Magnowska, Z.; Guardabassi, L. Antimicrobial synergy between carprofen and doxycycline against methicillin-resistant Staphylococcus pseudintermedius ST71. BMC Vet. Res. 2016, 12, 126. [Google Scholar] [CrossRef] [PubMed]
- Breukink, E.; de Kruijff, B. Lipid II as a target for antibiotics. Nat. Rev. Drug Discov. 2006, 5, 321–323. [Google Scholar] [CrossRef] [PubMed]
- ‘t Hart, P.; Oppedijk, S.F.; Breukink, E.; Martin, N.I. New Insights into Nisin’s Antibacterial Mechanism Revealed by Binding Studies with Synthetic Lipid II Analogues. Biochemistry 2016, 55, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Breukink, E.; Wiedemann, I.; Kraaij, C.V.; Kuipers, O.P.; Sahl, H.G.; de Kruijff, B. Use of the Cell Wall Precursor Lipid II by a Pore-Forming Peptide Antibiotic. Science 1999, 286, 2361. [Google Scholar] [CrossRef]
- Šíp, M.; Heřman, P.; Plášek, J.; Hrouda, V. Transmembrane potential measurement with carbocyanine dye diS-C3-(5): Fast fluorescence decay studies. J. Photochem. Photobiol. B Biol. 1990, 4, 321–328. [Google Scholar] [CrossRef]
- Strahl, H.; Hamoen, L.W. Membrane potential is important for bacterial cell division. Proc. Natl. Acad. Sci. USA 2010, 107, 12281–12286. [Google Scholar] [CrossRef]
- Ling, L.L.; Schneider, T.; Peoples, A.J.; Spoering, A.L.; Engels, I.; Conlon, B.P.; Mueller, A.; Schaberle, T.F.; Hughes, D.E.; Epstein, S.; et al. A new antibiotic kills pathogens without detectable resistance. Nature 2015, 517, 455–459. [Google Scholar] [CrossRef]
- Darzynkiewicz, Z.; Traganos, F.; Zhao, H.; Halicka, H.D.; Li, J. Cytometry of DNA replication and RNA synthesis: Historical perspective and recent advances based on “click chemistry”. Cytom. Part A 2011, 79, 328–337. [Google Scholar] [CrossRef]
- Schneider, T.; Kruse, T.; Wimmer, R.; Wiedemann, I.; Sass, V.; Pag, U.; Jansen, A.; Nielsen, A.K.; Mygind, P.H.; Ravents, D.S.; et al. Plectasin, a Fungal Defensin, Targets the Bacterial Cell Wall Precursor Lipid II. Science 2010, 328, 1168–1172. [Google Scholar] [CrossRef] [PubMed]
- Subbalakshmi, C.; Sitaram, N. Mechanism of antimicrobial action of indolicidin. Fems Microbiol. Lett. 1998, 160, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Wiedemann, I.; Benz, R.; Sahl, H.G. Lipid II-mediated pore formation by the peptide antibiotic nisin: A black lipid membrane study. J. Bacteriol. 2004, 186, 3259–3261. [Google Scholar] [CrossRef] [PubMed]
- Manabe, T.; Kawasaki, K. D-form KLKLLLLLKLK-NH2 peptide exerts higher antimicrobial properties than its L-form counterpart via an association with bacterial cell wall components. Sci. Rep. 2017, 7, 43384. [Google Scholar] [CrossRef]
- Oddo, A.; Thomsen, T.T.; Kjelstrup, S.; Gorey, C.; Franzyk, H.; Frimodt-Møller, N.; Løbner-Olesen, A.; Hansen, P.R. An all-D amphipathic undecapeptide shows promising activity against colistin-resistant strains of Acinetobacter baumannii and a dual mode of action. Antimicrob. Agents Chemother. 2016, 60, 592–599. [Google Scholar] [CrossRef]
- Ivankin, A.; Livne, L.; Mor, A.; Caputo, G.A.; DeGrado, W.F.; Meron, M.; Lin, B.; Gidalevitz, D. Role of the Conformational Rigidity in the Design of Biomimetic Antimicrobial Compounds. Angew. Chem. Int. Ed. 2010, 49, 8462–8465. [Google Scholar] [CrossRef]
- Jeong, M.C.; Jeon, D.; Shin, A.; Jin, S.; Shin, S.Y.; Park, Y.S.; Kim, Y. Effects of Hydrophobic Peptoid Substitutions on the Bacterial Cell Selectivity and Antimicrobial Activity of Piscidin 1. Bull. Korean Chem. Soc. 2016, 37, 1545–1551. [Google Scholar] [CrossRef]
- Mojsoska, B.; Carretero, G.; Larsen, S.; Mateiu, R.V.; Jenssen, H. Peptoids successfully inhibit the growth of gram negative E. coli causing substantial membrane damage. Sci. Rep. 2017, 7, 42332. [Google Scholar] [CrossRef]
- Blondeau, J.M.; Borsos, S.; Blondeau, L.D.; Blondeau, B.J. In vitro killing of Escherichia coli, Staphylococcus pseudintermedius and Pseudomonas aeruginosa by enrofloxacin in combination with its active metabolite ciprofloxacin using clinically relevant drug concentrations in the dog and cat. Vet. Microbiol. 2012, 155, 284–290. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | SEQUENCE a | MIC | HA f | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MSSP b | MRSP c | SA d | PA e | ||||||||||
| D2 | Nlys | Nlys | N1Nal | N4MePhe | Nlys | N1Nal | Nlys | NNle | 2–4 | 4 | 8 | 16 | 24 |
| 1 | Gly | Nlys | N1Nal | N4MePhe | Nlys | N1Nal | Nlys | NNle | 4 | 4 | 16 | 32 | 42 |
| 2 | Nlys | Gly | N1Nal | N4MePhe | Nlys | N1Nal | Nlys | NNle | 16 | 8 | 16 | 64 | 48 |
| 3 | Nlys | Nlys | Gly | N4MePhe | Nlys | N1Nal | Nlys | NNle | 64 | 64 | >64 | >64 | 56 |
| 4 | Nlys | Nlys | N1Nal | Gly | Nlys | N1Nal | Nlys | NNle | 64 | 32 | >64 | >64 | 40 |
| 5 | Nlys | Nlys | N1Nal | N4MePhe | Gly | N1Nal | Nlys | NNle | 4 | 4 | 32 | 16 | 25 |
| 6 | Nlys | Nlys | N1Nal | N4MePhe | Nlys | Gly | Nlys | NNle | >64 | >64 | >64 | >64 | 6 |
| 7 | Nlys | Nlys | N1Nal | N4MePhe | Nlys | N1Nal | Gly | NNle | 8 | 4 | 16 | 16 | 85 |
| 8 | Nlys | Nlys | N1Nal | N4MePhe | Nlys | N1Nal | Nlys | Gly | 8 | 8 | 32 | 32–64 | 49 |
| ID | SEQUENCE a | MIC | HA f | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MSSP b | MRSP c | SA d | PA e | ||||||||||
| D2 | Nlys | Nlys | N1Nal | N4MePhe | Nlys | N1Nal | Nlys | NNle | 2–4 | 4 | 8 | 16 | 24 |
| 9 | NMe | 4 | 4 | 16 | 32 | 16 | |||||||
| 10 | N4MePhe | 8 | 8 | 8 | 16 | 69 | |||||||
| 11 | N1Nal | 8 | 8 | 8 | 16 | 87 | |||||||
| 12 | NEtOH | 8 | 8 | 32 | >64 | 5 | |||||||
| ID | SEQUENCE a | MIC | HA f | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MSSP b | MRSP c | SA d | PA e | ||||||||||
| D2 | Nlys | Nlys | N1Nal | N4MePhe | Nlys | N1Nal | Nlys | NNle | 2–4 | 4 | 8 | 16 | 24 |
| D2L | Lys | Lys | 1Nal | Phe | Lys | 1Nal | Lys | Nle | 16 | 8 | 32 | 16 | 35 |
| D2D | lys | lys | 1nal | phe | lys | 1nal | lys | nle | 1–4 | 4 | 4 | 4 | 35 |
| D2R | Nle | Lys | 1Nal | Lys | Phe | 1Nal | Lys | Lys | 16 | 4 | 32 | 32–64 | 23 |
| 13 | Nlys | Nlys | N1Nal | N4MePhe | lys | 1nal | lys | nle | 4 | 4 | 8 | 8 | 34 |
| 14 | lys | lys | 1nal | phe | Nlys | N1Nal | Nlys | NNle | 4 | 4 | 16 | 64 | 5 |
| 15 | Nlys | Nlys | 1nal | phe | Nlys | 1nal | Nlys | nle | 4 | 4 | 32 | >64 | 6 |
| 16 | lys | lys | N1Nal | N4MePhe | lys | N1Nal | lys | NNle | 4 | 4 | 16 | 64 | 4 |
| Bacteria | D2 | D2D |
|---|---|---|
| Acinetobacter baumannii, 27065, 16 D1, dog, wound, 2010 | >64 | 16 |
| Corynebacterium auriscanis, 31551, 54 C6, dog, ear, 2013 | 2 | 2 |
| E. coli, 30235, 23 A6, dog, wound, 2012 | 64 | 4 |
| Enterococcus faecalis, 27404, 17 C7, dog, wound, 2011 | >64 | 8 |
| Enterococcus faecium, 30951, 24 C1, dog, ear, 2013 | 16 | 4 |
| Klebsiella pneumoniae, 26233, 11 H5, dog, wound, 2010 | >64 | 8 |
| Proteus mirabilis, 25178, 9 A4, dog, ear, 2009 | >64 | >64 |
| Pseudomonas aeruginosa (26314, 12 C5, dog, urine, 2010) | 32 | 8 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Greco, I.; Hansen, J.E.; Jana, B.; Molchanova, N.; Oddo, A.; Thulstrup, P.W.; Damborg, P.; Guardabassi, L.; Hansen, P.R. Structure–Activity Study, Characterization, and Mechanism of Action of an Antimicrobial Peptoid D2 and Its d- and l-Peptide Analogues. Molecules 2019, 24, 1121. https://doi.org/10.3390/molecules24061121
Greco I, Hansen JE, Jana B, Molchanova N, Oddo A, Thulstrup PW, Damborg P, Guardabassi L, Hansen PR. Structure–Activity Study, Characterization, and Mechanism of Action of an Antimicrobial Peptoid D2 and Its d- and l-Peptide Analogues. Molecules. 2019; 24(6):1121. https://doi.org/10.3390/molecules24061121
Chicago/Turabian StyleGreco, Ines, Johannes E. Hansen, Bimal Jana, Natalia Molchanova, Alberto Oddo, Peter W. Thulstrup, Peter Damborg, Luca Guardabassi, and Paul R. Hansen. 2019. "Structure–Activity Study, Characterization, and Mechanism of Action of an Antimicrobial Peptoid D2 and Its d- and l-Peptide Analogues" Molecules 24, no. 6: 1121. https://doi.org/10.3390/molecules24061121
APA StyleGreco, I., Hansen, J. E., Jana, B., Molchanova, N., Oddo, A., Thulstrup, P. W., Damborg, P., Guardabassi, L., & Hansen, P. R. (2019). Structure–Activity Study, Characterization, and Mechanism of Action of an Antimicrobial Peptoid D2 and Its d- and l-Peptide Analogues. Molecules, 24(6), 1121. https://doi.org/10.3390/molecules24061121