Isothiocyanates and Glucosinolates from Sisymbrium officinale (L.) Scop. (“the Singers’ Plant”): Isolation and in Vitro Assays on the Somatosensory and Pain Receptor TRPA1 Channel




Abstract
1. Introduction
1.1. Sisymbrium officinale (Erysimum), “the Singers’ Plant”
1.2. Glucosinolates and Isothiocyanates in SO
1.3. TRPA1 Channel as the Target of Pungent Principles in Plants
1.4. Aim of This Work
2. Results
2.1. Preliminary Analysis of SO Volatiles
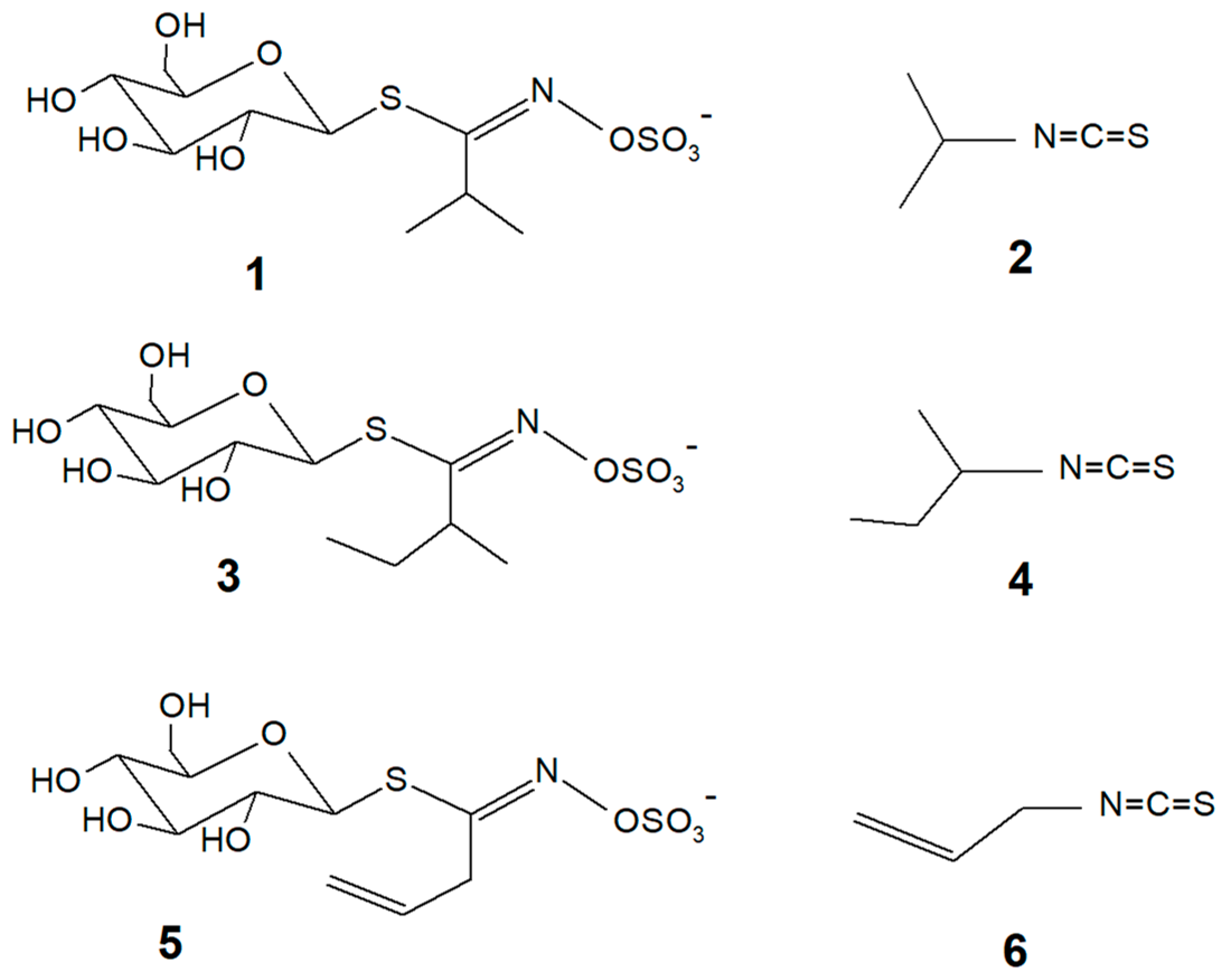
2.2. Isolation and Synthesis of Glucoputranjivin 1
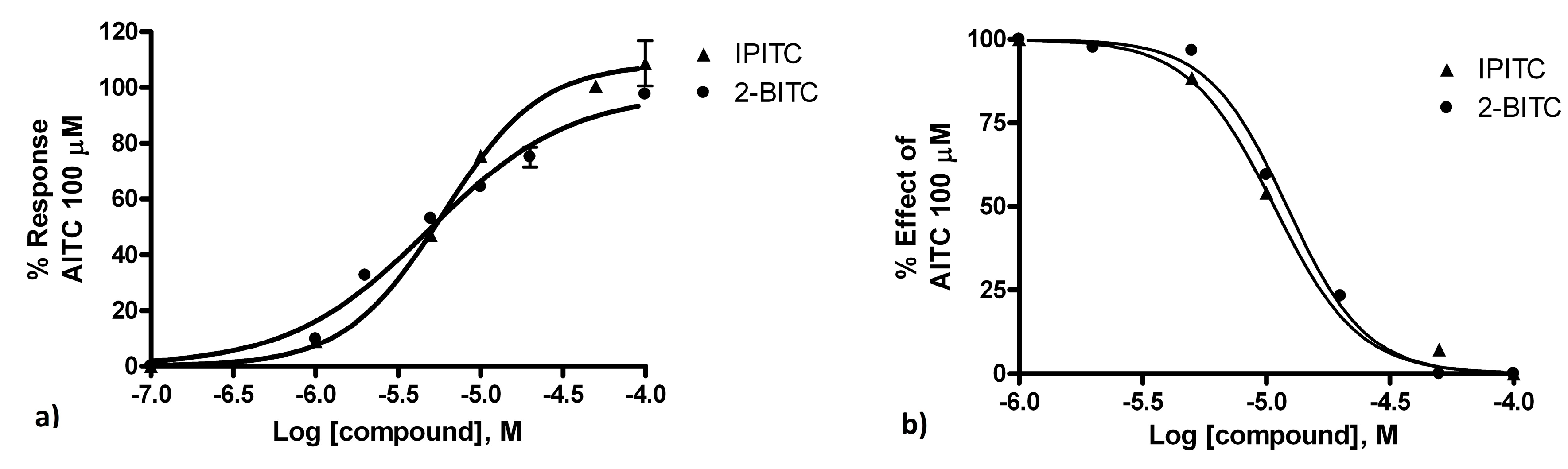
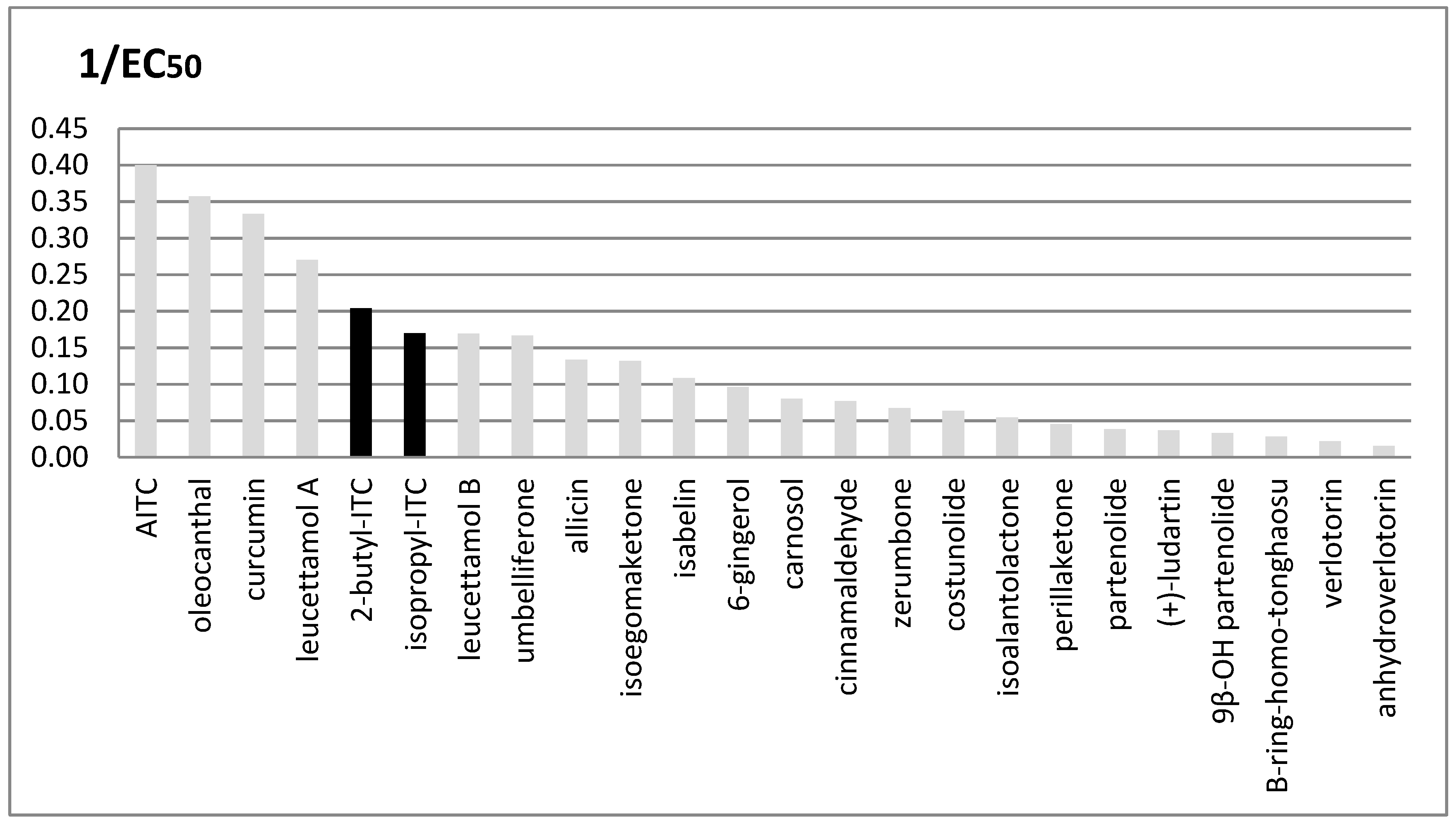
2.3. In Vitro Assays
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Chemicals and Equipment
4.3. HS- SPME Coupled with GCMS Analysis
4.4. Glucoputranjivin, Isolation and Purification
4.5. In Vitro Assays with rTRP Ion Channels
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Erisimo a Milano. Available online: URL www.erisimo-a-milano.it (accessed on 17 February 2019).
- Gruenwald, J.; Brendler, T.; Jaenicke, C. Hedge mustard- Sisymbrium officinale. In PDR for Herbal Medicines, 3rd ed.; Thompson: Stamford, CT, USA, 2004; pp. 426–427. ISBN 1-56363-512-7. [Google Scholar]
- Leclerc, H. Précis de Phytothérapie; Masson Editions: Paris, France, 1983; pp. 252–253. ISBN 2225858705. [Google Scholar]
- Biagi, M. Phytotherapy in Arts Medicine. In Proceedings of the International Conference CoMeT, Milano, Italy, 4–6 July 2016; pp. 17–19. [Google Scholar]
- Blazević, I.; Radonić, A.; Mastelić, J.; Zekić, M.; Skocibusić, M.; Maravic, A. Hedge mustard (Sisymbrium officinale): Chemical diversity of volatiles and their antimicrobial activity. Chem. Biodivers. 2010, 7, 2023–2034. [Google Scholar] [CrossRef] [PubMed]
- Di Sotto, A.; Di Giacomo, S.; Vitalone, A.; Nicoletti, M.; Mazzanti, G. Antimutagenic thiocompounds from Sisymbrium officinale. J. Nat. Prod. 2012, 75, 2062–2068. [Google Scholar] [CrossRef] [PubMed]
- Calcinoni, O. Sisymbrium “Singers’ Plant” Efficacy in Reducing Perceived Vocal Tract Disability. J. Otolaryngol.-ENT Res. 2017, 8, 00243. [Google Scholar] [CrossRef]
- Zimbaldi, N. Sisymbrium officinale (L.) Scop: Indagine fitochimica sui glucosinolati caratterizzanti la specie. Master’s Thesis, Università degli Studi di Milano, Herbs Science and Technology, Milano, Italy, 5 April 2018. [Google Scholar]
- Belitz, H.D.; Grosch, W.; Shieberle, P. Food Chemistry, 4th ed.; Springer: Leipzig, Germany, 2009; pp. 789–790. ISBN 978-3-540-69935-4. [Google Scholar]
- Fahey, J.W.; Zalcmann, A.T.; Talalay, P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 2001, 56, 5–51. [Google Scholar] [CrossRef]
- Lee, J.G.; Lim, S.; Lee, E.J. The mechanism of deterioration of the glucosinolate-myrosynase system in radish roots during cold storage after harvest. Food Chem. 2017, 233, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Carnat, A.; Fraisse, D.; Carnat, A.P.; Groubert, A.; Lamaison, J.L. Normalisation de l’erysimum, Sisymbrium officinale L. Ann. Pharm. Fr. 1998, 56, 36–39. [Google Scholar] [PubMed]
- EMA—European Medicines Agency-EMA. Assessment report on Sisymbrium officinale (L.) Scop., Herba; EMA/HMPC/280194/2013 Committee on Herbal Medicinal Products (HMPC): London, UK, 15 January 2014. [Google Scholar]
- Di Sotto, A.; Vitalone, A.; Nicoletti, M.; Piccin, A.; Mozzanti, G. Pharmacological and phytochemical study on a Sisymbrium officinale Scop. Extract. J. Ethnopharmacol. 2010, 127, 731–736. [Google Scholar] [CrossRef]
- Érysimum, Sisymbrium officinale, Pharmacopée Française 1998. ANSM-Agence Nationale du Sécurité du Médicament et des produits de santé: Saint-Denis, France. Available online: https://www.ansm.sante.fr/Mediatheque/Publications/Pharmacopee-francaise-Plan-Preambule-index (accessed on 17 February 2019).
- Venkatachalam, K.; Montell, C. TRP channels. Annu Rev. Biochem. 2007, 76, 387–417. [Google Scholar] [CrossRef]
- Nilius, B.; Owsianik, G. The transient receptor potential family of ion channels. Genome Biol. 2011, 12, 218–229. [Google Scholar] [CrossRef]
- Nilius, B.; Appendino, G. Spices: The savory and beneficial science of pungency. Rev. Physiol. Biochem. Pharmacol. 2013, 164, 1–76. [Google Scholar] [CrossRef]
- Nilius, B.; Appendino, G.; Owsianik, G. The Transient Receptor Potential Channel TRPA1: From gene to pathophysiology. Pflugers Arch. 2012, 464, 425–458. [Google Scholar] [CrossRef] [PubMed]
- Anand, U.; Otto, W.R.; Facer, P.; Zebda, N.; Selmer, I.; Gunthorpe, M.J.; Chessell, I.P.; Sinisi, M.; Birch, R.; Anand, P. TRPA1 receptor localisation in the human peripheral nervous system and functional studies in cultured human and rat sensory neurons. Neurosci. Lett. 2008, 438, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Atoyan, R.; Shander, D.; Botchkareva, N.V. Non-neuronal expression of transient receptor potential type A1 (TRPA1) in human skin. J. Invest. Dermatol. 2009, 129, 2312–2315. [Google Scholar] [CrossRef] [PubMed]
- Nassini, R.; Pedretti, P.; Moretto, N.; Fusi, C.; Carnini, C.; Facchinetti, F.; Viscomi, A.R.; Pisano, A.R.; Stokesberry, S.; Brunmark, C.; et al. Transient receptor potential ankyrin 1 channel localized to non-neuronal airway cells promotes non-neurogenic inflammation. PLoS ONE 2012, 7, e42454. [Google Scholar] [CrossRef] [PubMed]
- Hinman, A.; Chuang, H.H.; Bautista, D.M.; Julius, D. TRP channel activation by reversible covalent modification. Proc. Natl. Acad. Sci. USA 2006, 103, 19564–19568. [Google Scholar] [CrossRef] [PubMed]
- Vriens, J.; Nilius, B.; Vennekens, R. Herbal compounds and toxins modulating TRP channels. Curr. Neuropharmacol. 2008, 6, 79–96. [Google Scholar] [CrossRef] [PubMed]
- Jordt, S.E.; Bautista, D.M.; Chuang, H.H.; McKemy, D.D.; Zygmunt, P.M.; Högestätt, E.D.; Meng, I.D.; Julius, D. Mustard oils and cannabinoids excite sensory nerve fibres through the TRP channel ANKTM1. Nature 2004, 427, 260–265. [Google Scholar] [CrossRef]
- Pawliszyn, J. Solid Phase Microextraction: Theory and Practice; Wiley VCH: New York, NY, USA, 1997; ISBN 0-471-19034-9. [Google Scholar]
- Rohloff, J.; Bones, A.M. Volatile profiling of Arabidopsis thaliana–Putative olfactory compounds in plant communications. Phytochemistry 2005, 66, 1941–1955. [Google Scholar] [CrossRef]
- Borgonovo, G.; Zimbaldi, N.; Bassoli, A.; (University of Milan, Milano, Italy). Unpublised work, 2019.
- Kiddle, G.; Bennett, R.N.; Botting, N.P.; Davidson, N.E.; Robertson, A.A.; Wallsgrove, R.M. High-performance liquid chromatographic separation of natural and synthetic desulpho- glucosinolates and their chemical vadation by UV, NMR and chemical ionisation-MS methods. Phytoche. Anal. 2001, 12, 226–242. [Google Scholar] [CrossRef]
- Dauvergne, X.; Cérantola, S.; Salaün, S.; Magné, C.; Kervarec, N.; Bessièrs, M.; Deslandes, E. General occurrence of the glucosinolate glucocochlearin within the Cochlearia genus. Carbohydr. Res. 2006, 341, 2166–2169. [Google Scholar] [CrossRef]
- Davidson, N.E.; Rutherford, T.J.; Botting, N.P. Synthesis, analysis and rearrangement of novel unnatural glucosinolates. Carbohydr. Res. 2001, 330, 295–307. [Google Scholar] [CrossRef]
- Boonen, B.; Startek, J.B.; Talavera, K. Chemical activation of sensory TRP channels. In Taste and Smell; Kratwurst, D., Ed.; Springer International Publishing: Basel, Switzerland, 2016; pp. 73–113. ISBN 978-3-319-48925-4. [Google Scholar]
- Giorgi, A.; Bassoli, A.; Borgonovo, G.; Panseri, S.; Manzo, A.; Pentimalli, D.; Schiano Moriello, A.; De Petrocellis, L. Extracts and compounds active on TRP ion channels from Waldehimia glabra, a ritual medicinal plant from Himalaya. Phytomedicine 2017, 32, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Guarise, M.; Borgonovo, G.; Bassoli, A.; Ferrante, A. Evaluation of two ecotypes of hedge mustard (Sisymbrium officinale (L.) Scop.) as a potential leafy vegetable. Horticulturae 2019, 5, 13. [Google Scholar] [CrossRef]
- Benn, M.H.; Meakin, D. Glucoputranjivin. Can. J. Chem. 1965, 43, 1874–1877. [Google Scholar] [CrossRef]
- Schiano-Moriello, A.; De Petrocellis, L. Assay of TRPV1 Receptor Signaling. Methods Mol. Biol. 2016, 1412, 65–76. [Google Scholar] [CrossRef]
Sample Availability: Sample of compound 1 is available from the authors. |




{kind=link}
{kind=link}
{kind=link}
| C | Assignment | δC (ppm) | δH (ppm, Molteplicity, (J in Hz)) |
|---|---|---|---|
| Aliphatic moiety | |||
| 1 | C | 165.68 | |
| 2 | CH | 34.03 | 2.97, m (6.90) |
| 3 | CH3 | 21.33 * | 1.298 *, d (6.90) |
| 4 | CH3 | 22.45 * | 1.296 *, d (6.90) |
| Glucose moiety | |||
| 1’ |  | 4.89, d (9.75) | |
| 2’ |  | ||
| 3’ | 62.78–86.91 | ||
| 4’ | |||
| 5’ | |||
| 6’ | |||
| TRPA1 | ||||
|---|---|---|---|---|
| cpd | Name | Efficacy (%AITC 100μM) | Potency EC50 μM | IC50 Inhibition TRPA1 μM (AITC 100μM) |
| 5 | (−)-Sinigrin | <10 | NA | >100 |
| 1 | Gluco-putranjivine | <10 | NA | >100 |
| 2 | Isopropyl isothiocyanate | 108.8 ± 1.6 | 5.9 ± 0.4 | 10.8 ± 0.4 |
| 4 | 2-buthyl isothiocyanate | 98.0 ± 1.0 | 4.9 ± 0.2 | 12.1 ± 0.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borgonovo, G.; Zimbaldi, N.; Guarise, M.; De Nisi, P.; De Petrocellis, L.; Schiano Moriello, A.; Bassoli, A. Isothiocyanates and Glucosinolates from Sisymbrium officinale (L.) Scop. (“the Singers’ Plant”): Isolation and in Vitro Assays on the Somatosensory and Pain Receptor TRPA1 Channel. Molecules 2019, 24, 949. https://doi.org/10.3390/molecules24050949
Borgonovo G, Zimbaldi N, Guarise M, De Nisi P, De Petrocellis L, Schiano Moriello A, Bassoli A. Isothiocyanates and Glucosinolates from Sisymbrium officinale (L.) Scop. (“the Singers’ Plant”): Isolation and in Vitro Assays on the Somatosensory and Pain Receptor TRPA1 Channel. Molecules. 2019; 24(5):949. https://doi.org/10.3390/molecules24050949
Chicago/Turabian StyleBorgonovo, Gigliola, Nathan Zimbaldi, Marta Guarise, Patrizia De Nisi, Luciano De Petrocellis, Aniello Schiano Moriello, and Angela Bassoli. 2019. "Isothiocyanates and Glucosinolates from Sisymbrium officinale (L.) Scop. (“the Singers’ Plant”): Isolation and in Vitro Assays on the Somatosensory and Pain Receptor TRPA1 Channel" Molecules 24, no. 5: 949. https://doi.org/10.3390/molecules24050949
APA StyleBorgonovo, G., Zimbaldi, N., Guarise, M., De Nisi, P., De Petrocellis, L., Schiano Moriello, A., & Bassoli, A. (2019). Isothiocyanates and Glucosinolates from Sisymbrium officinale (L.) Scop. (“the Singers’ Plant”): Isolation and in Vitro Assays on the Somatosensory and Pain Receptor TRPA1 Channel. Molecules, 24(5), 949. https://doi.org/10.3390/molecules24050949