Sweroside Alleviated LPS-Induced Inflammation via SIRT1 Mediating NF-κB and FOXO1 Signaling Pathways in RAW264.7 Cells



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
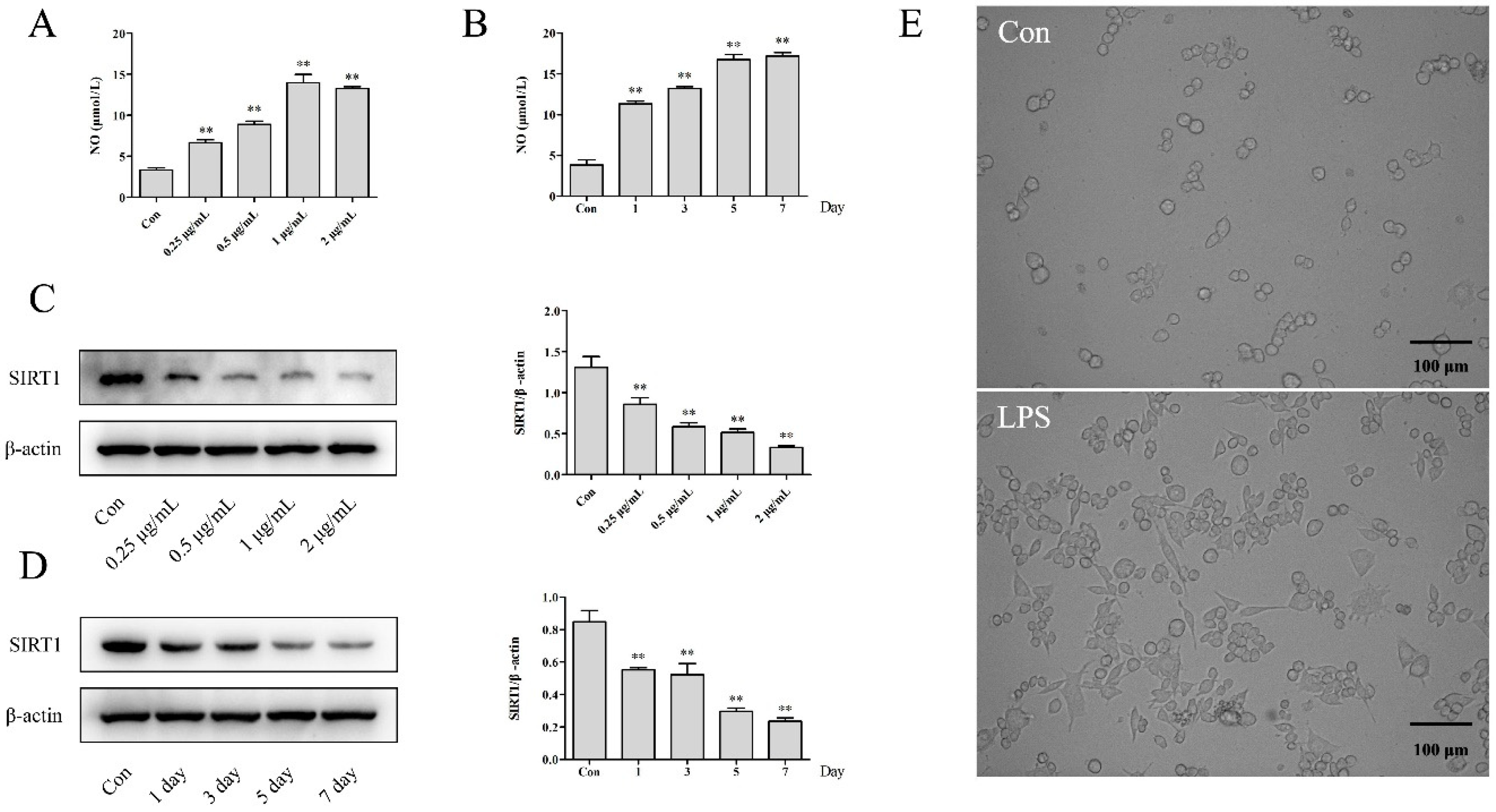
2.1. The Effects of NO Production and Morphology in LPS-Induced RAW264.7 Cells
2.2. The Effects on the Expression of SIRT1 in LPS-Induced RAW264.7 Cells
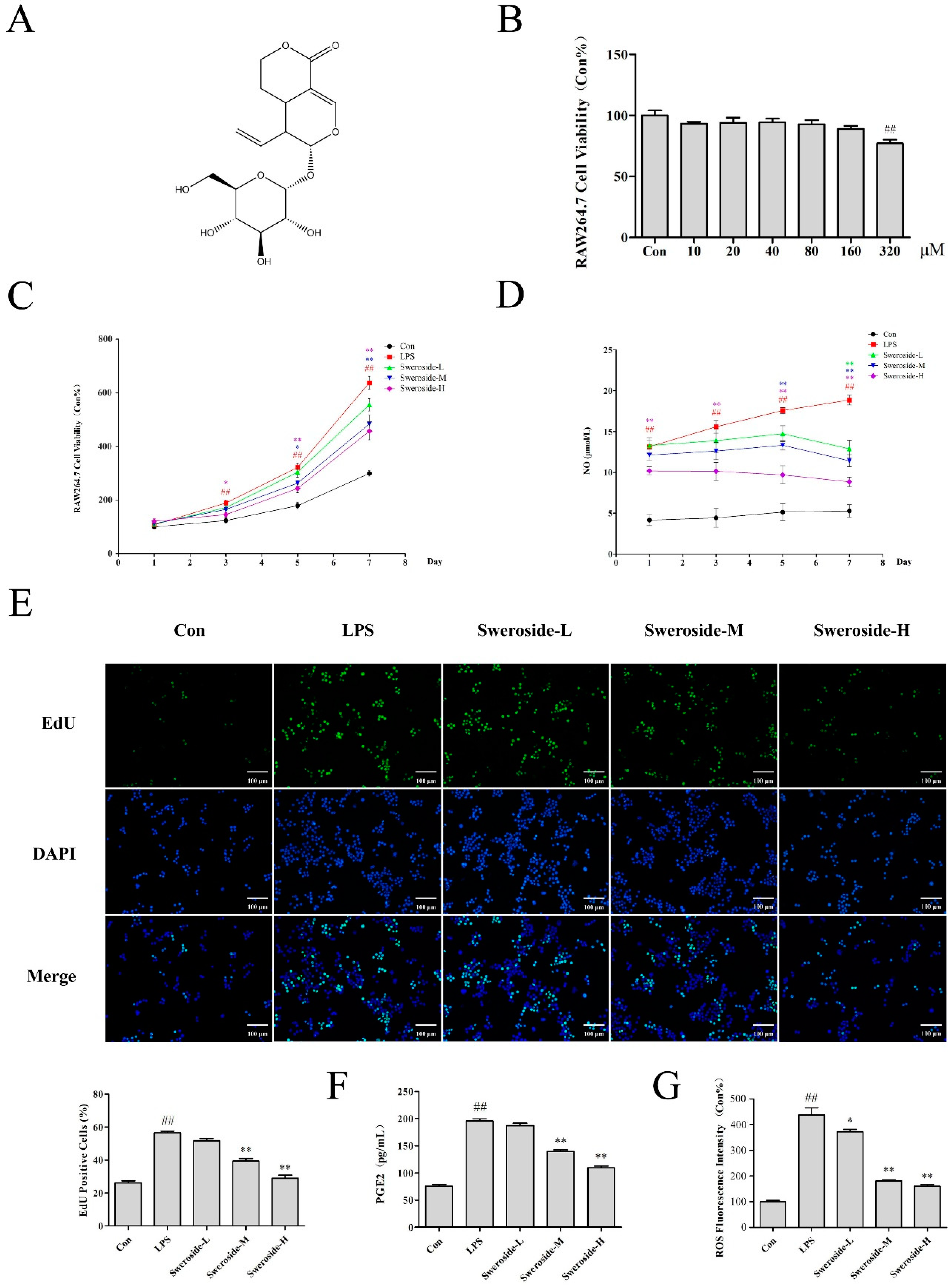
2.3. The Effects of Cell Proliferation and NO Production on Sweroside Treatment in LPS-Induced RAW264.7 Cells
2.4. The Effects of PGE2 and ROS on Sweroside Treatment in LPS-Induced RAW264.7 Cells
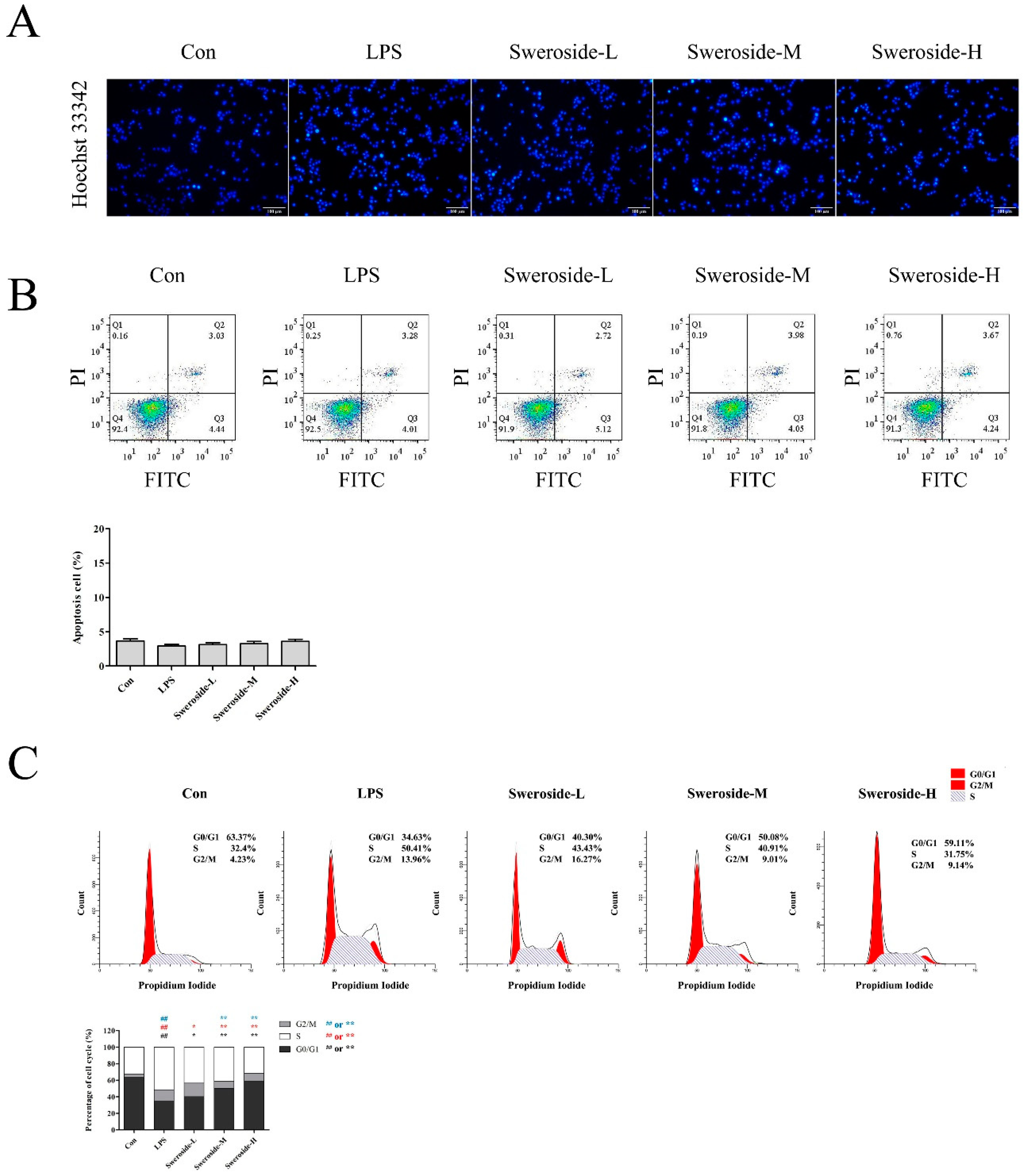
2.5. The Effects of Apoptosis and Cell-Cycle Distribution on Sweroside Treatment in LPS-Induced RAW264.7 Cells
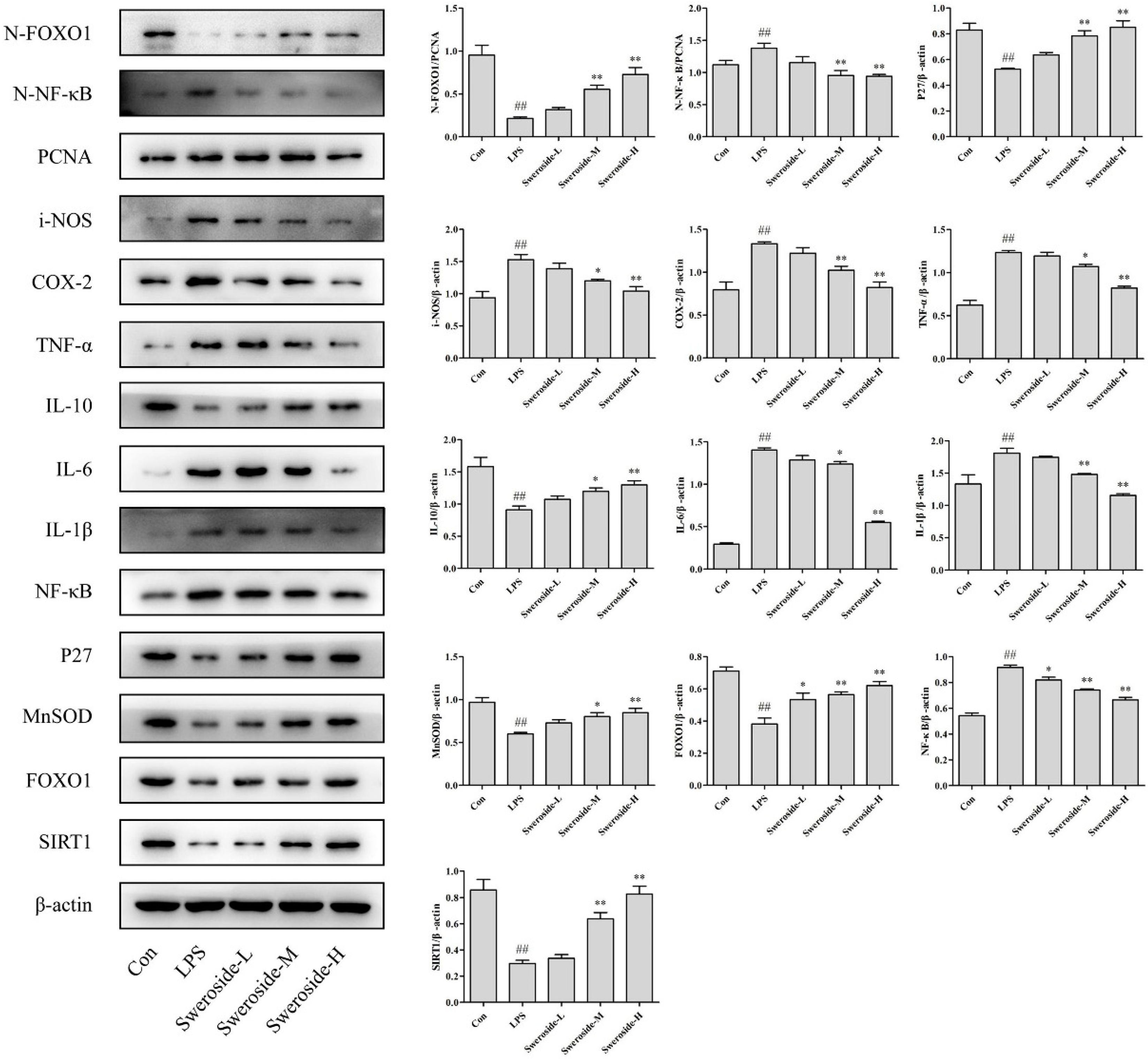
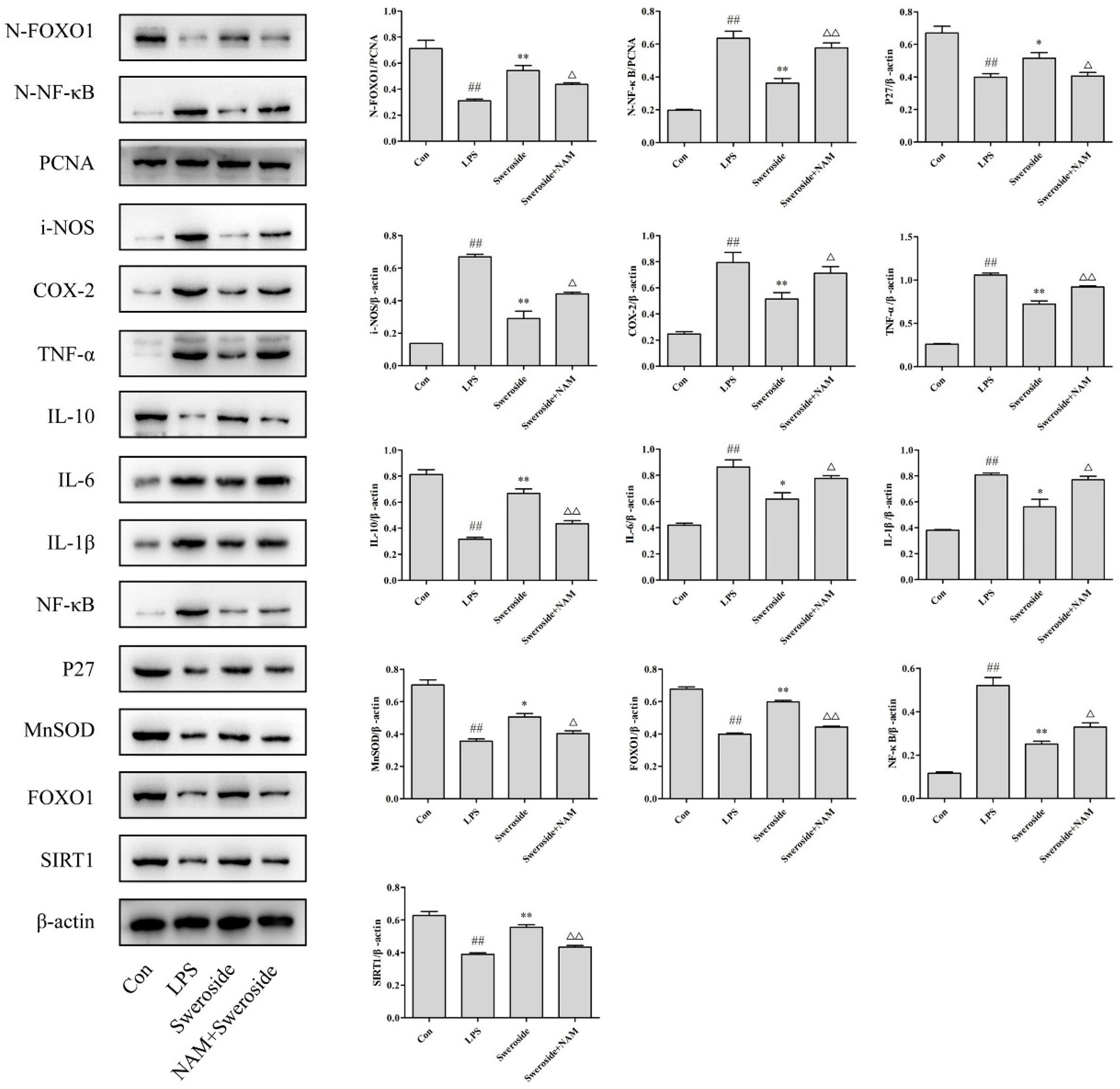
2.6. The Effects of SIRT1/NF-κB on Sweroside Treatment in LPS-Induced RAW264.7 Cells
2.7. The Effects of SIRT1/FOXO1 on Sweroside Treatment in LPS-Induced RAW264.7 Cells
2.8. The Effects of NAM on Sweroside Treatment in LPS-Induced RAW264.7 Cells
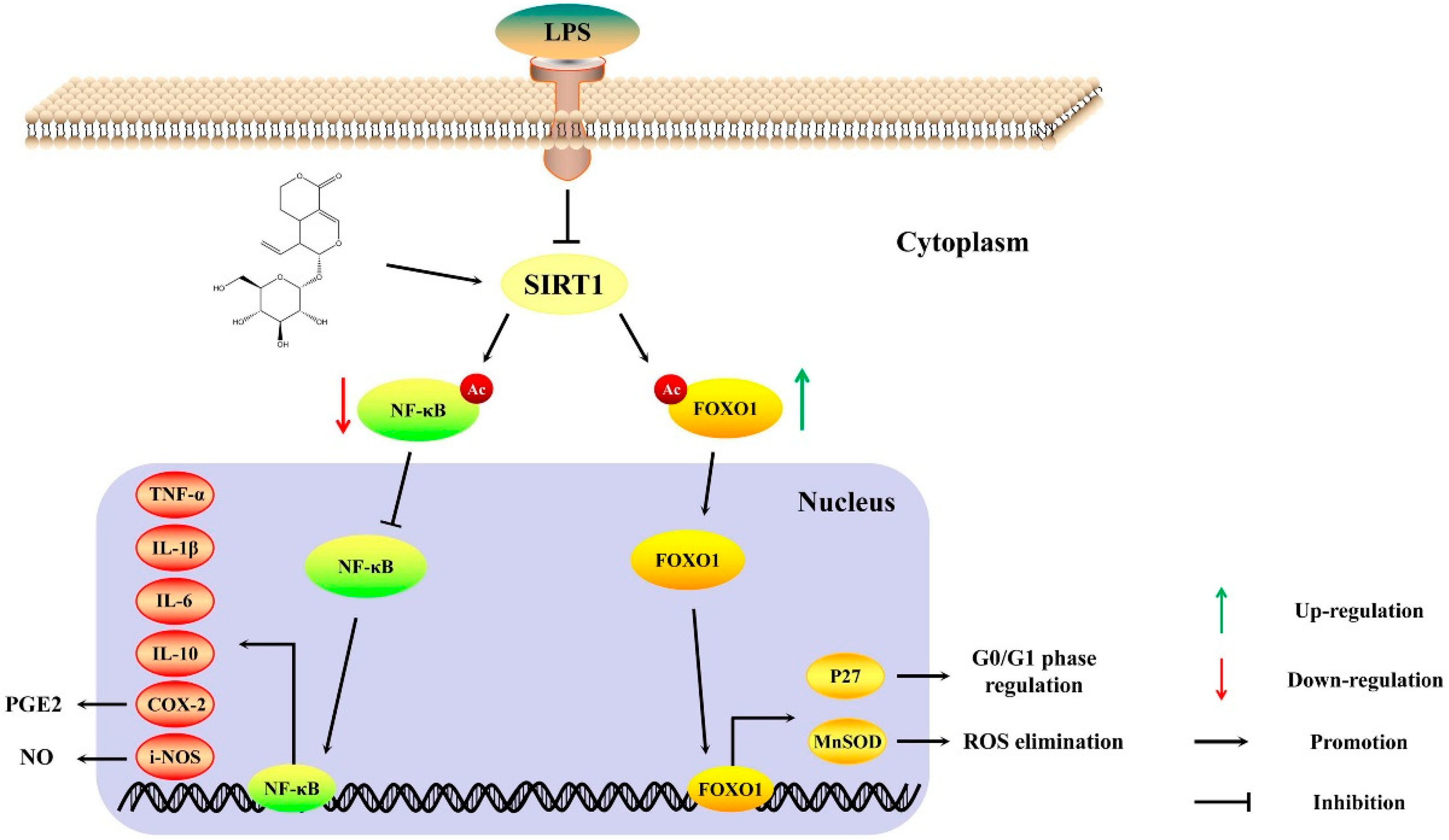
3. Discussion
4. Materials and Methods
4.1. Chemicals and Regents
4.2. Cell Culture and Treatment
4.3. Cytotoxicity Assay
4.4. Determination of NO Production
4.5. Microscopic Observation
4.6. Cell Proliferation Assay for Treatment of Sweroside in LPS-Induced RAW264.7 Cells
4.7. PGE2 and ROS Analysis
4.8. Cell Apoptotic Analysis by Hoechst 33342 Stain and Flow Cytometer
4.9. Cell-Cycle Distribution Analysis by Flow Cytometer
4.10. Western Blot Analysis
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zhang, T.; Li, H.; Shi, J.; Li, S.; Li, M.; Zhang, L.; Zheng, L.; Zheng, D.; Tang, F.; Zhang, X.; et al. p53 predominantly regulates IL-6 production and suppresses synovial inflammation in fibroblast-like synoviocytes and adjuvant-induced arthritis. Arthritis. Res. Ther. 2016, 18, 271. [Google Scholar] [CrossRef] [PubMed]
- Firestein, G.S. Evolving concepts of rheumatoid arthritis. Nature 2003, 423, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Mateen, S.; Zafar, A.; Moin, S.; Khan, A.Q.; Zubair, S. Understanding the role of cytokines in the pathogenesis of rheumatoid arthritis. Clin. Chim. Acta 2016, 455, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Weyand, C.M.; Zeisbrich, M.; Goronzy, J.J. Metabolic signatures of T-cells and macrophages in rheumatoid arthritis. Curr. Opin. Immunol. 2017, 46, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hu, J.; Lin, J.; Fang, W.; Du, G. Anti-inflammatory and analgesic effects of ethanol and aqueous extracts of Pterocephalus hookeri (C.B. Clarke) Höeck. J. Ethnopharmacol. 2009, 123, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yu, H.; Guo, F.; Wu, Y.; Li, Y. Antinociceptive and anti-inflammatory activities of a standardizedextract of bis-iridoids from Pterocephalus hookeri. J. Ethnopharmacol. 2018, 216, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Fan, G.; Li, Q.; Su, J.; Meng, X.; Zhang, Y. Simultaneous determination of ten compounds in two main medicinal plant parts of Tibetan herb, Pterocephalus hookeri (CB Clarke) Höeck, by ultra-high performance liquid chromatography-photodiode array. Trop. J. Pharm. Res. 2017, 16, 1407–1416. [Google Scholar] [CrossRef]
- Yang, Q.L.; Yang, F.; Gong, J.T.; Tang, X.W.; Wang, G.Y.; Wang, Z.T.; Yang, L. Sweroside ameliorates α-naphthylisothiocyanate-induced cholestatic liver injury in mice by regulating bile acids and suppressing pro-inflammatory responses. Acta Pharmacol. Sin. 2016, 37, 1218–1228. [Google Scholar] [CrossRef] [PubMed]
- Mahendran, G.; Thamotharan, G.; Sengottuvelu, S.; Narmatha Bai, V. Anti-diabetic activity of Swertia corymbosa (Griseb.) Wight ex C.B. Clarke aerial parts extract in streptozotocin induced diabetic rats. J. Ethnopharmacol. 2014, 151, 1175–1183. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Wang, C.; Jiang, H.; Tian, X.; Li, W.; Liang, W.; Yang, J.; Zhong, C.; Chen, Y.; Li, T. Protective Effects of Sweroside on IL-1β-Induced Inflammation in Rat Articular Chondrocytes Through Suppression of NF-κB and mTORC1 Signaling Pathway. Inflammation 2018, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Haigis, M.C.; Sinclair, D.A. Mammalian sirtuins: Biological insights and disease relevance. Annu. Rev. Pathol. 2010, 5, 253–295. [Google Scholar] [CrossRef] [PubMed]
- Rajendrasozhan, S.; Yang, S.; Kinnula, V.L.; Rahman, I. SIRT1, an Antiinflammatory and Antiaging Protein, Is Decreased in Lungs of Patients with Chronic Obstructive Pulmonary Disease. Am. J. Resp. Crit. Care 2008, 177, 861–870. [Google Scholar] [CrossRef] [PubMed]
- Ferrero-Miliani, L.; Nielsen, O.H.; Andersen, P.S.; Girardin, S.E. Chronic inflammation: Importance of NOD2 and NALP3 in interleukin-1β generation. Clin. Exp. Immunol. 2006, 147, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Underhill, D.M.; Goodridge, H.S. Information processing during phagocytosis. Nat. Rev. Immunol. 2012, 12, 492–502. [Google Scholar] [CrossRef] [PubMed]
- Papathanassiu, A.E.; Ko, J.; Imprialou, M.; Bagnati, M.; Srivastava, P.K.; Vu, H.A.; Cucchi, D.; McAdoo, S.P.; Ananieva, E.A.; Mauro, C.; et al. BCAT1 controls metabolic reprogramming in activated human macrophages and is associated with inflammatory diseases. Nat. Commun. 2017, 8, 16040. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, P.; Chen, Y.; Chen, Y.; Chung, L.; Tamilselvi, S.; Shen, C.; Day, C.H.; Chen, R.; Viswanadha, V.P.; Kuo, W.; et al. The multifaceted link between inflammation and human diseases. J. Cell. Physiol. 2018, 233, 6458–6471. [Google Scholar] [CrossRef] [PubMed]
- Bascands, J.; Bachvarova, M.; Neau, E.; Schanstra, J.P.; Bachvarov, D. Molecular determinants of LPS-induced acute renal inflammation: Implication of the kinin B1 receptor. Biochem. Biophys. Res. Commun. 2009, 386, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Zhang, X.; Zhang, L. Negative regulation of inflammation by SIRT1. Pharmacol. Res. 2013, 67, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Shan, Y.; Wu, Y.; Xu, C.; Yu, X.; Zhao, J.; Yan, J.; Shang, W. Berberine suppresses LPS-induced inflammation through modulating Sirt1/NF-κB signaling pathway in RAW264.7 cells. Int. Immunopharmacol. 2017, 52, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Kandikattu, H.K.; Rachitha, P.; Jayashree, G.V.; Krupashree, K.; Sukhith, M.; Majid, A.; Amruta, N.; Khanum, F. Anti-inflammatory and anti-oxidant effects of Cardamom (Elettaria repens (Sonn.) Baill) and its phytochemical analysis by 4D GCXGC TOF-MS. Biomed. Pharmacother. 2017, 91, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Aziz, R.S.; Siddiqua, A.; Shahzad, M.; Shabbir, A.; Naseem, N. Oxyresveratrol ameliorates ethanol-induced gastric ulcer via downregulation of IL-6, TNF-α, NF-ĸB, and COX-2 levels, and upregulation of TFF-2 levels. Biomed. Pharmacother. 2019, 110, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage plasticity, polarization, and function in health and disease. J. Cell. Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef] [PubMed]
- Baatar, D.; Siddiqi, M.Z.; Im, W.T.; Ul Khaliq, N.; Hwang, S.G. Anti-Inflammatory Effect of Ginsenoside Rh2-Mix on Lipopolysaccharide-Stimulated RAW 264.7 Murine Macrophage Cells. J. Med. Food. 2018, 21, 951–960. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.J.; Lee, J.E.; Lee, A.S.; Kang, K.P.; Lee, S.; Park, S.K.; Lee, S.Y.; Han, M.K.; Kim, D.H.; Kim, W. SIRT1 overexpression decreases cisplatin-induced acetylation of NF-κB p65 subunit and cytotoxicity in renal proximal tubule cells. Biochem. Biophys. Res. Commun. 2012, 419, 206–210. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Gong, Y.; Xu, L.; Zhou, M.; Li, J.; Song, J. Bidirectional regulation of osteogenic differentiation by the FOXO subfamily of Forkhead transcription factors in mammalian MSCs. Cell. Proliferat. 2018, e12540. [Google Scholar] [CrossRef] [PubMed]
- Giannakou, M.E.; Partridge, L. The interaction between FOXO and SIRT1: Tipping the balance towards survival. Trends Cell. Biol. 2004, 14, 408–412. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, M.A.; Zhu, Q.; Hammond, J.M. FoxO1a Can Alter Cell Cycle Progression by Regulating the Nuclear Localization of p27kip in Granulosa Cells. Mol. Endocrinol. 2004, 18, 1756–1767. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Shi, X.; Xu, X.; Lin, Y.; Shao, Z.; Wu, R.; Huang, L. Liraglutide ameliorates early renal injury by the activation of renal FoxO1 in a type 2 diabetic kidney disease rat model. Diabetes Res. Clin. Pract. 2018, 137, 173–182. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, R.; Dong, Z.; Lan, X.; Liao, Z.; Chen, M. Sweroside Alleviated LPS-Induced Inflammation via SIRT1 Mediating NF-κB and FOXO1 Signaling Pathways in RAW264.7 Cells. Molecules 2019, 24, 872. https://doi.org/10.3390/molecules24050872
Wang R, Dong Z, Lan X, Liao Z, Chen M. Sweroside Alleviated LPS-Induced Inflammation via SIRT1 Mediating NF-κB and FOXO1 Signaling Pathways in RAW264.7 Cells. Molecules. 2019; 24(5):872. https://doi.org/10.3390/molecules24050872
Chicago/Turabian StyleWang, Rui, Zhaoyue Dong, Xiaozhong Lan, Zhihua Liao, and Min Chen. 2019. "Sweroside Alleviated LPS-Induced Inflammation via SIRT1 Mediating NF-κB and FOXO1 Signaling Pathways in RAW264.7 Cells" Molecules 24, no. 5: 872. https://doi.org/10.3390/molecules24050872
APA StyleWang, R., Dong, Z., Lan, X., Liao, Z., & Chen, M. (2019). Sweroside Alleviated LPS-Induced Inflammation via SIRT1 Mediating NF-κB and FOXO1 Signaling Pathways in RAW264.7 Cells. Molecules, 24(5), 872. https://doi.org/10.3390/molecules24050872