P-glycoprotein Inhibitor Tariquidar Potentiates Efficacy of Astragaloside IV in Experimental Autoimmune Encephalomyelitis Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
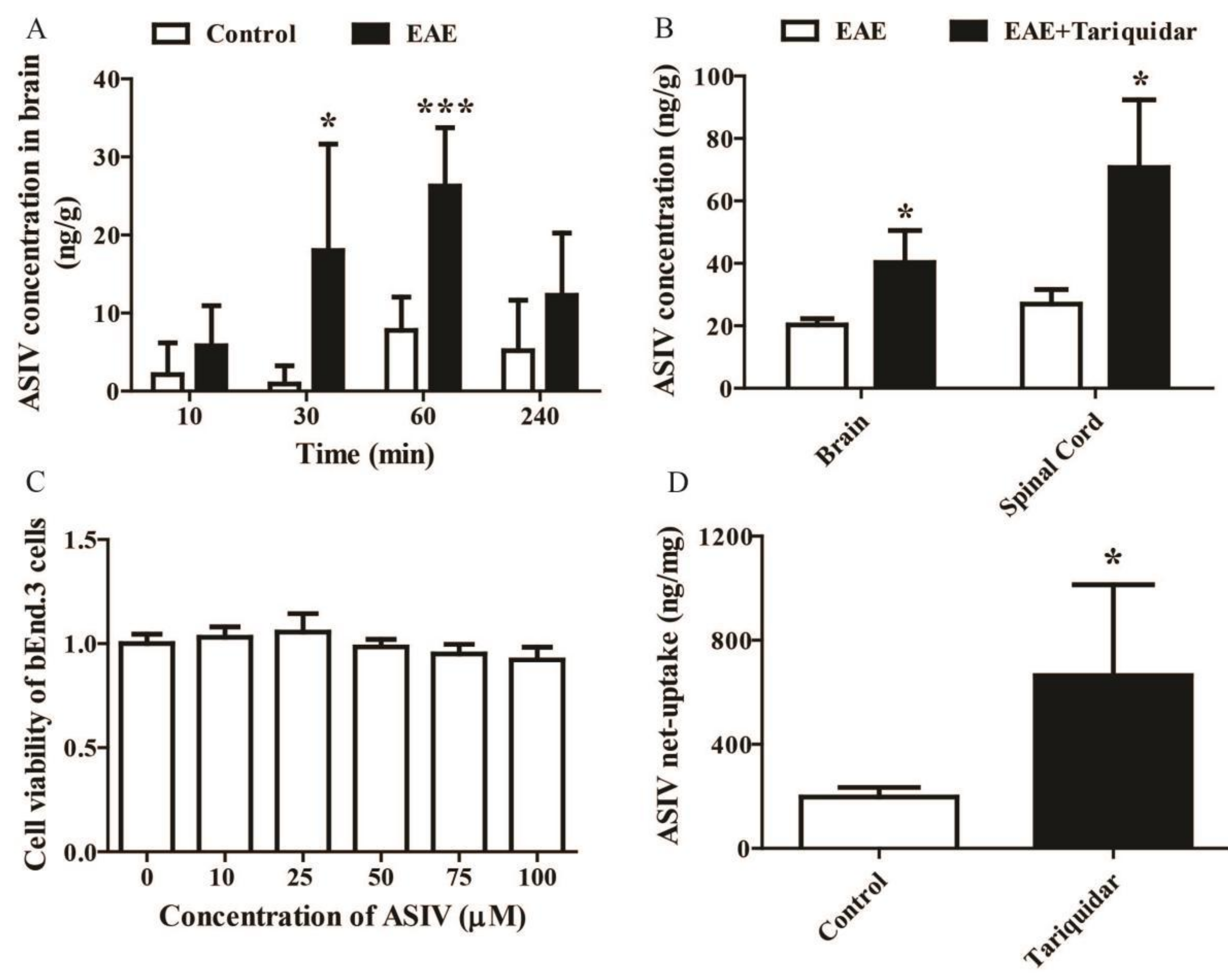
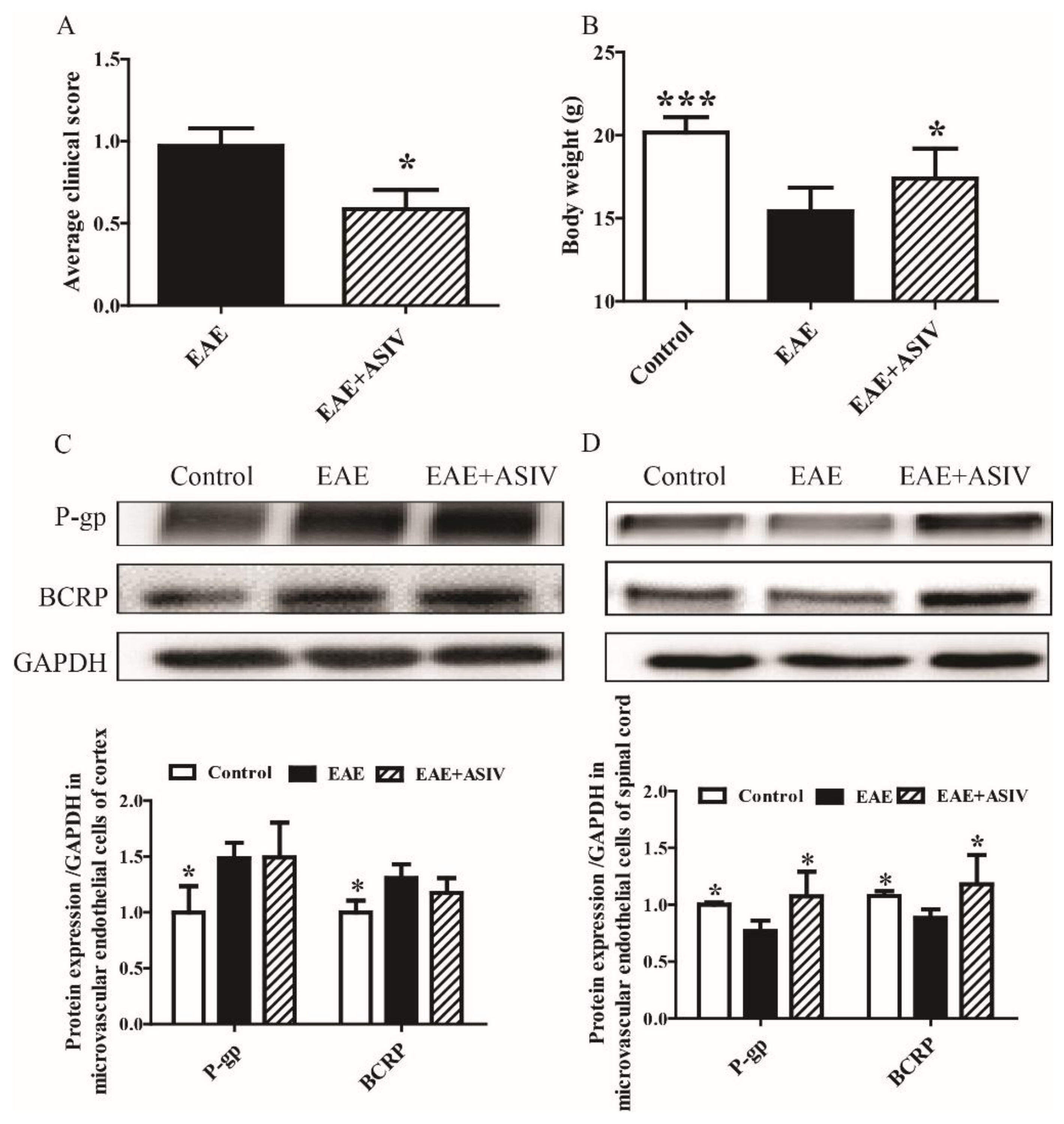
2.1. ASIV Changed the Expression Pattern of P-gp and BCRP in BSCB but Not BBB of EAE Mice
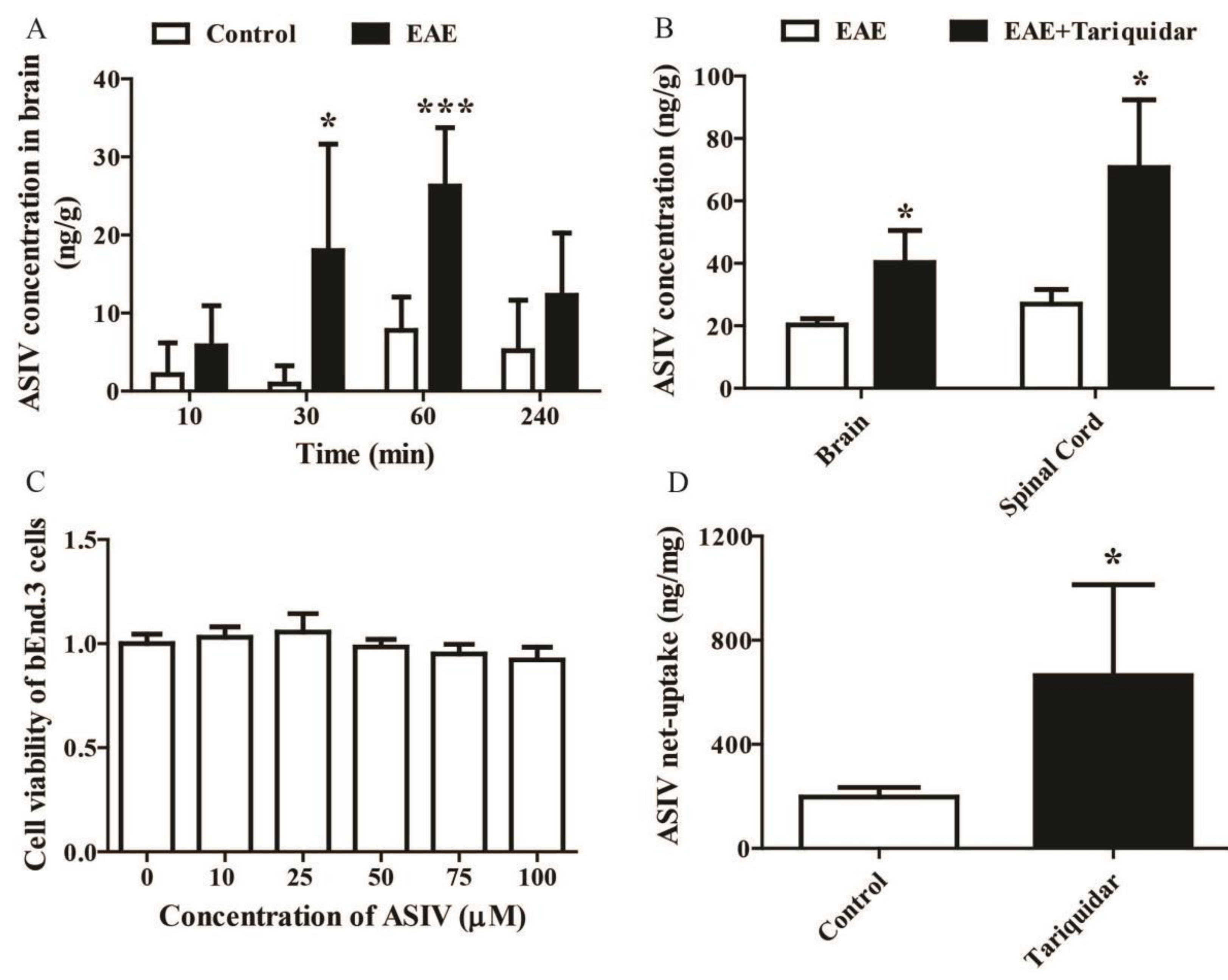
2.2. Tariquidar Facilitated the Penetration of ASIV into CNS of EAE Mice
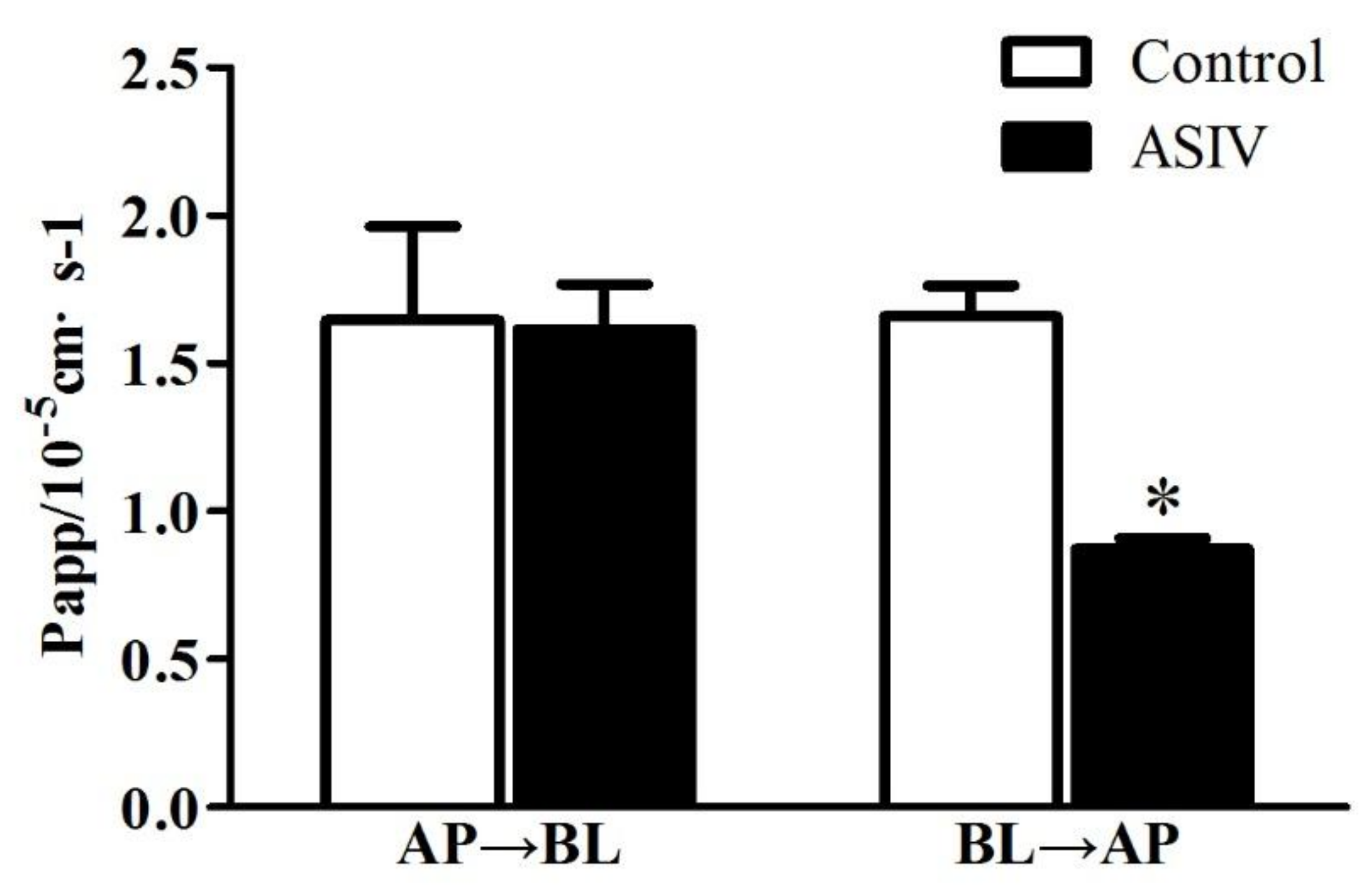
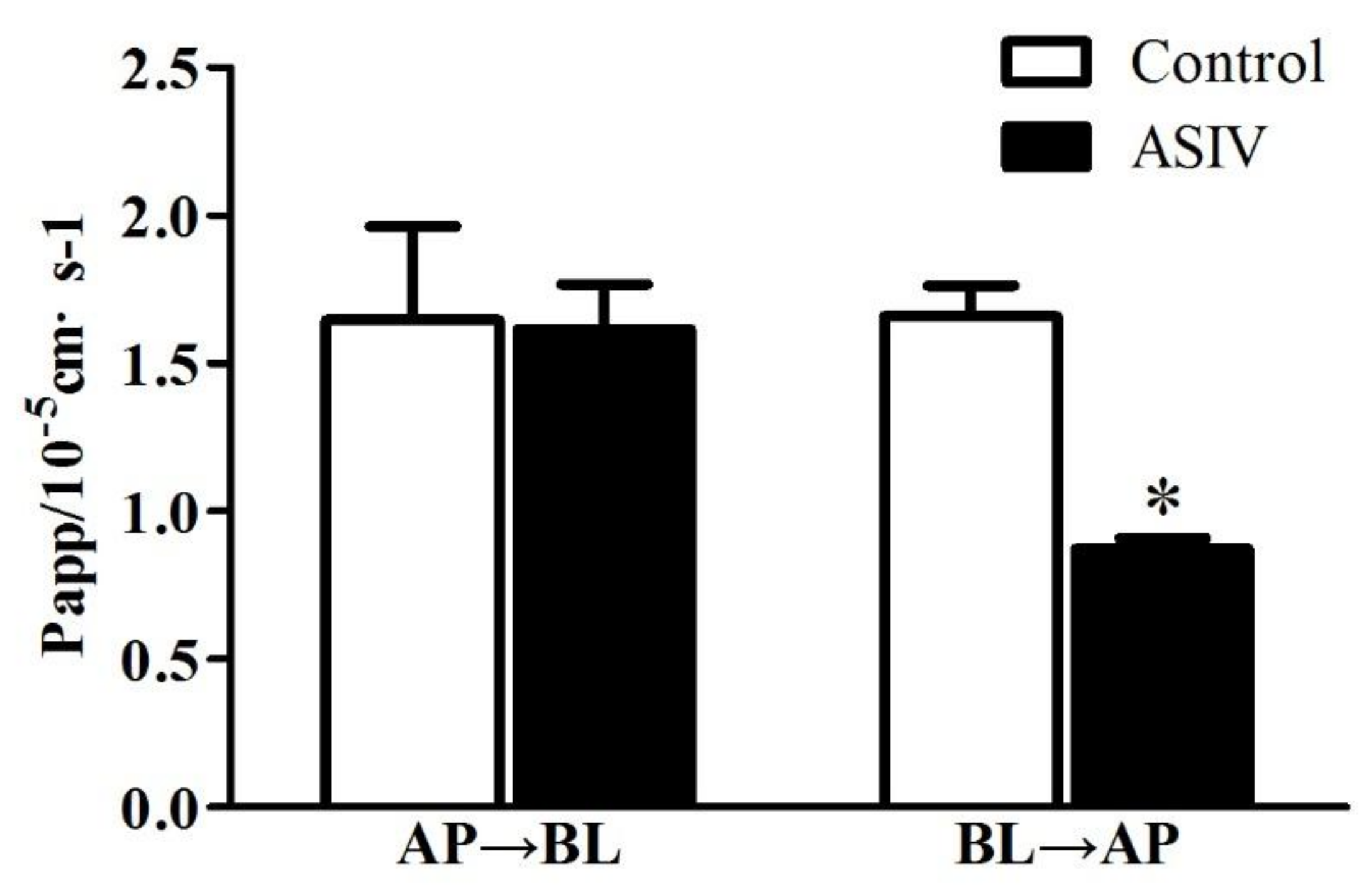
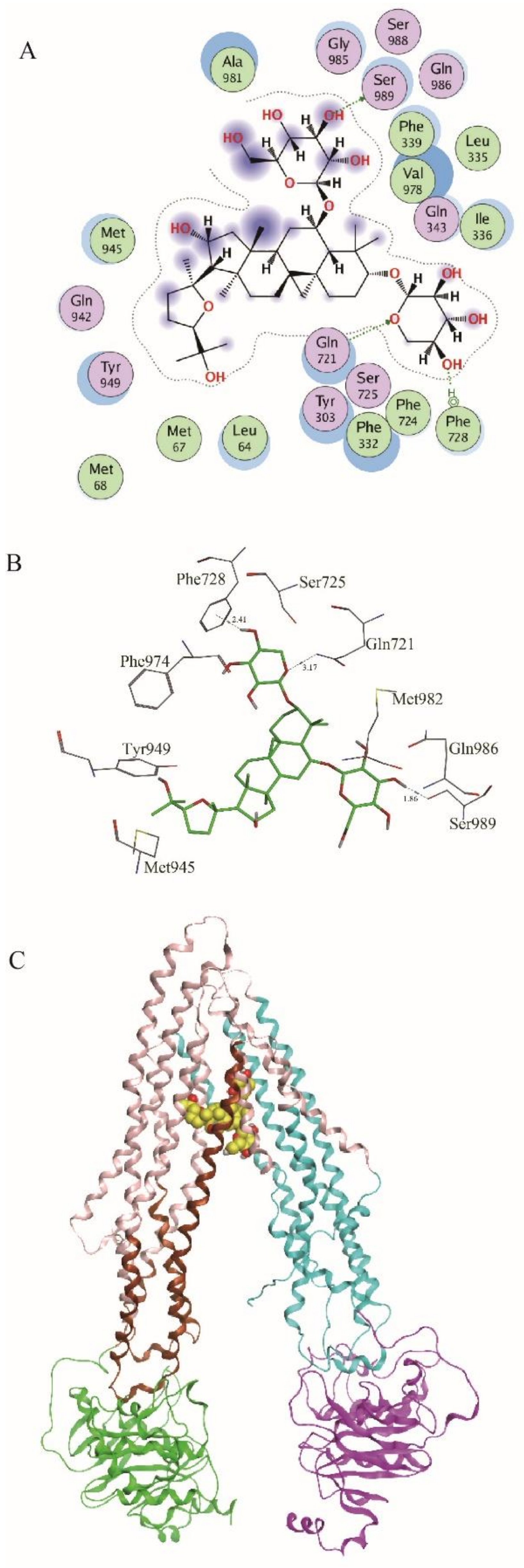
2.3. ASIV Was a Potential Substrate of P-gp
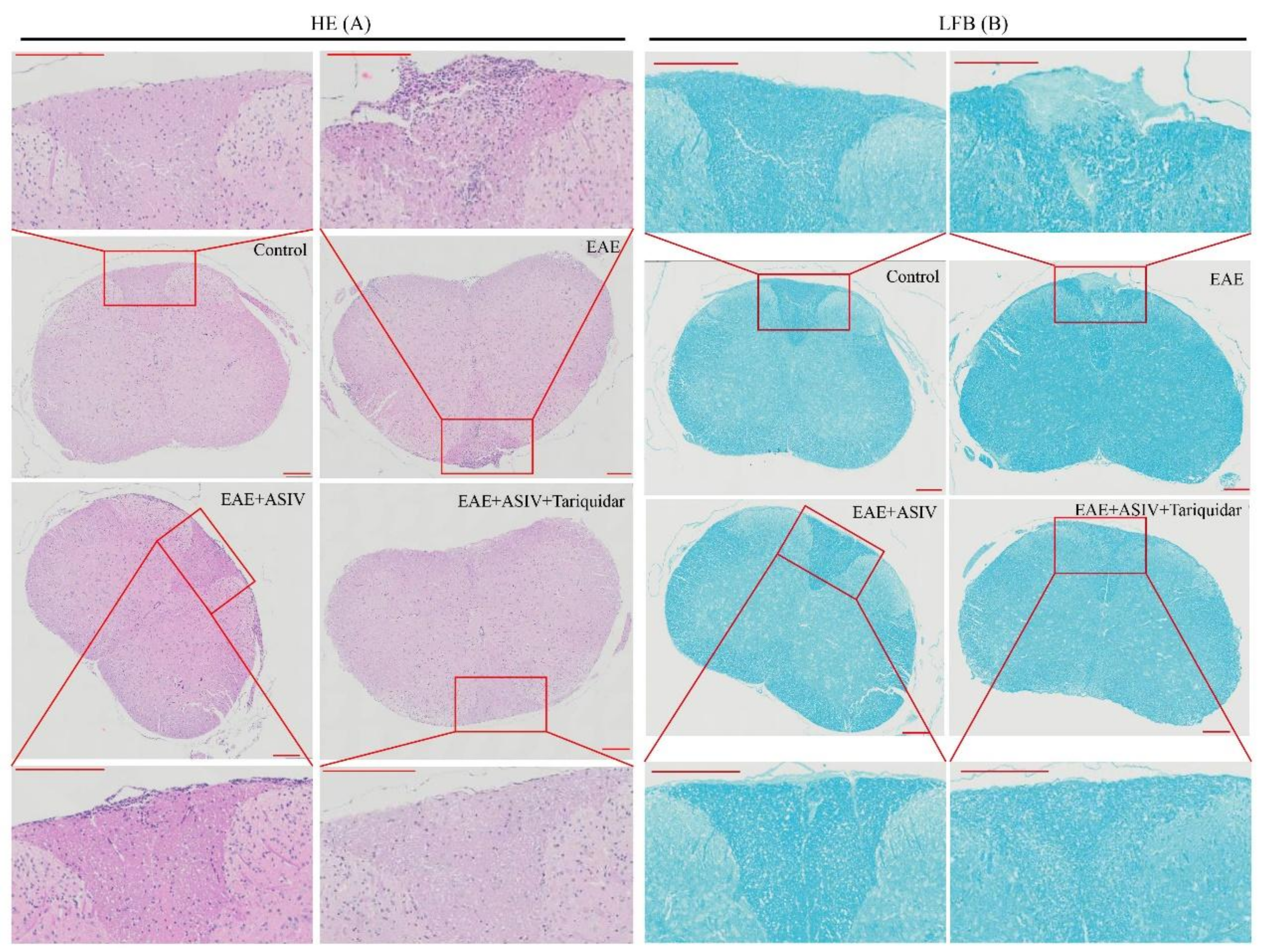
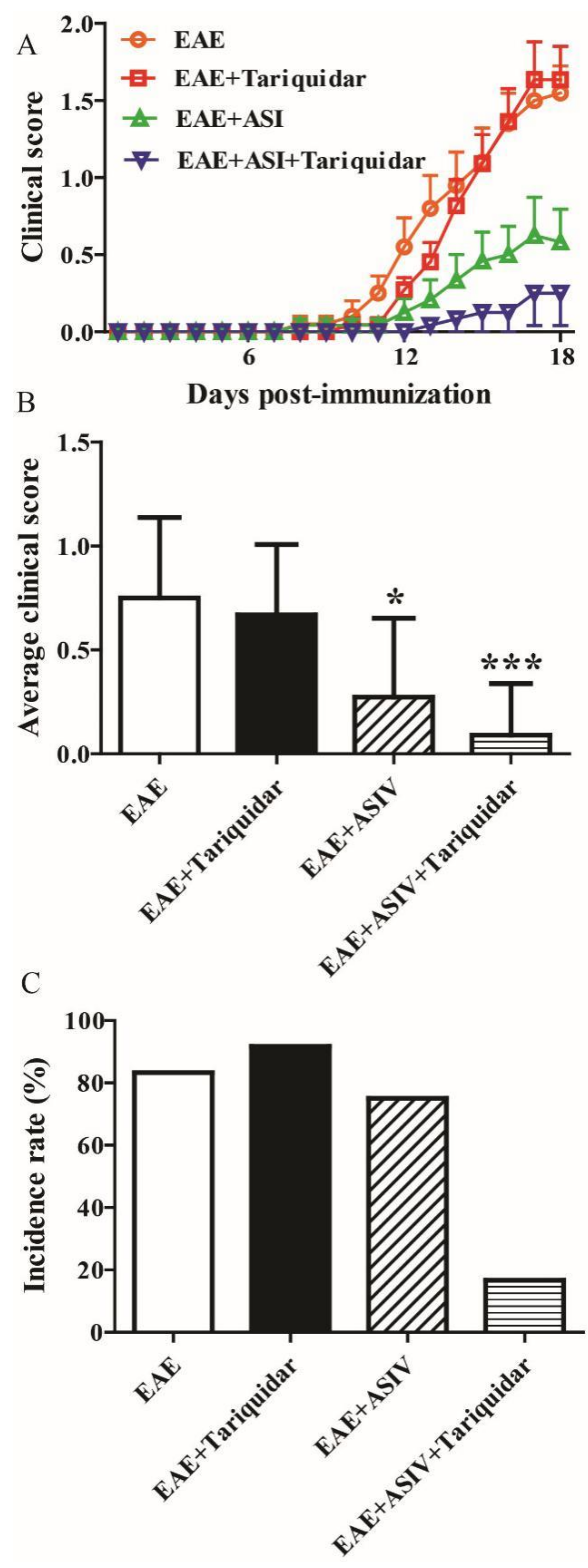
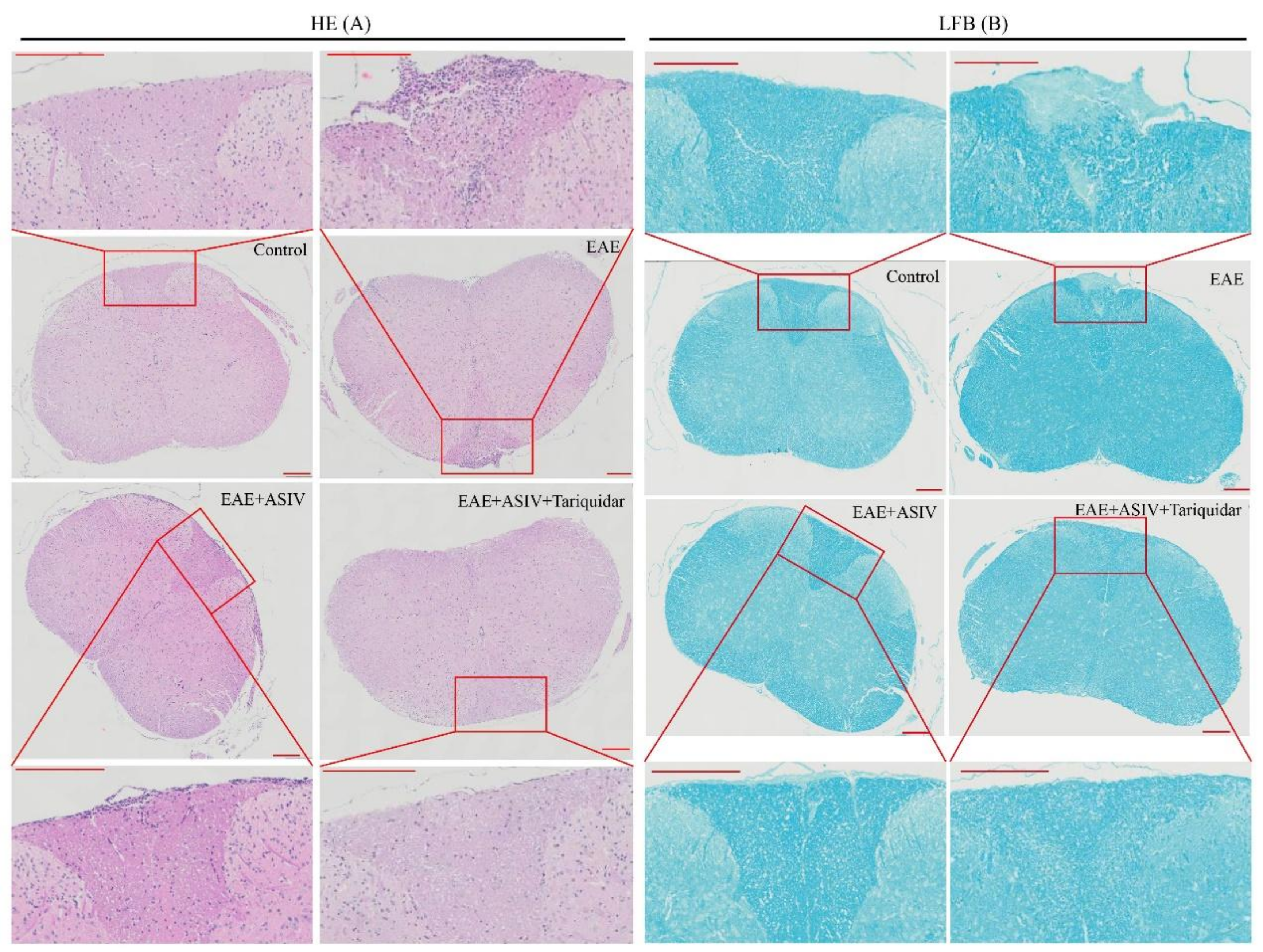
2.4. Tariquidar Potentiated the Efficacy of ASIV on EAE Mice
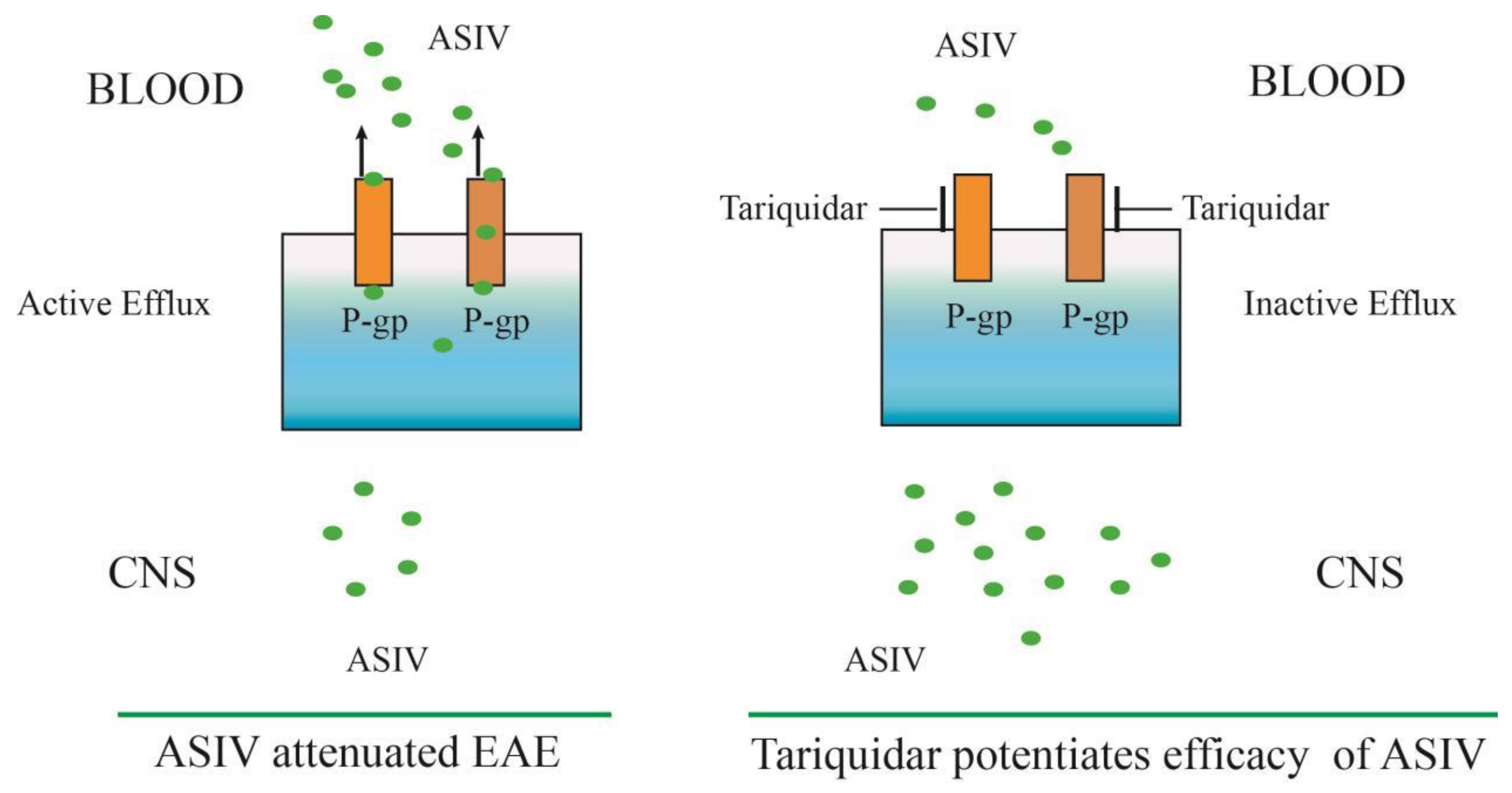
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Animals
4.3. EAE Induction and Treatment
4.4. Histopathology
4.5. Microvessel Isolation
4.6. LC-MS/MS Analysis
4.7. Western Blot Analysis
4.8. Cell Culture
4.9. CCK-8 Assay
4.10. ASIV Net Uptake Assay
4.11. ASIV Transmembrane Absorption
4.12. Molecular Docking Experiment
4.13. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Bartanusz, V.; Jezova, D.; Alajajian, B.; Digicaylioglu, M. The blood-spinal cord barrier: Morphology and clinical implications. Ann. Neurol. 2011, 70, 194–206. [Google Scholar] [CrossRef] [PubMed]
- Reinhold, A.K.; Rittner, H.L. Barrier function in the peripheral and central nervous system—A review. Pflugers Arch. 2017, 469, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Zlokovic, B.V. The blood-brain barrier in health and chronic neurodegenerative disorders. Neuron 2008, 57, 178–201. [Google Scholar] [CrossRef] [PubMed]
- Tenorio, G.; Kulkarni, A.; Kerr, B.J. Resident glial cell activation in response to perispinal inflammation leads to acute changes in nociceptive sensitivity: Implications for the generation of neuropathic pain. Pain 2013, 154, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Aubé, B.; Lévesque, S.A.; Paré, A.; Chamma, É.; Kébir, H.; Gorina, R.; Lécuyer, M.; Alvarez, J.I.; Koninck, Y.D.; Engelhardt, B.; et al. Neutrophils mediate blood-spinal cord barrier disruption in demyelinating neuroinflammatory diseases. J. Immunol. 2014, 193, 2438–2454. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.Y.; Wang, Z.B.; Wang, Y.; Tong, L.C.; Li, Y.; Wei, X.; Luan, P.; Li, L. Increasing the Permeability of the Blood-brain Barrier in Three Different Models in vivo. CNS Neurosci. Ther. 2015, 21, 568–574. [Google Scholar] [CrossRef] [PubMed]
- Abbott, N.J. Blood-brain barrier structure and function and the challenges for CNS drug delivery. J. Inherit. Metab. Dis. 2013, 36, 437–449. [Google Scholar] [CrossRef]
- Kooij, G.; van Horssen, J.; Bandaru, V.V.; Haughey, N.J.; de Vries, H.E. The Role of ATP-Binding Cassette Transporters in Neuro-Inflammation: Relevance for Bioactive Lipids. Front. Pharmacol. 2012, 3, 74. [Google Scholar] [CrossRef]
- Miller, D.S. Regulation of ABC transporters blood-brain barrier: The good, the bad, and the ugly. Adv. Cancer Res. 2015, 125, 43–70. [Google Scholar]
- Chan, G.N.; Evans, R.A.; Banks, D.B.; Mesev, E.V.; Miller, D.S.; Cannon, R.E. Selective induction of P-glycoprotein at the CNS barriers during symptomatic stage of an ALS animal model. Neurosci. Lett. 2017, 639, 103–113. [Google Scholar] [CrossRef]
- Vliet, E.A.V.; van Schaik, R.; Edelbroek, P.M.; Voskuyl, R.A.; Redeker, S.; Aronica, E.; Wadman, W.J.; Gorter, J.A. Region-specific overexpression of P-glycoprotein at the blood-brain barrier affects brain uptake of phenytoin in epileptic rats. J. Pharmacol. Exp. Ther. 2007, 322, 141–147. [Google Scholar] [CrossRef]
- Rees, D.C.; Johnson, E.; Lewinson, O. ABC transporters: The power to change. Nat. Rev. Mol. Cell Biol. 2009, 10, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Higgins, C.F. ABC transporters: From microorganisms to man. Annu. Rev. Cell Biol. 1992, 8, 67–113. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Chang, Z.; Wang, W.; Wang, B. Rapamycin-Mediated mTOR Inhibition Reverses Drug Resistance to Adriamycin in Colon Cancer Cells. Hepatogastroenterology 2015, 62, 880–886. [Google Scholar] [PubMed]
- O’Brien, F.E.; O’Connor, R.M.; Clarke, G.; Dinan, T.G.; Griffin, B.T.; Cryan, J.F. P-glycoprotein inhibition increases the brain distribution and antidepressant-like activity of Escitalopram in rodents. Neuropsychopharmacology 2013, 38, 2209–2219. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Hou, J.; Chen, X.; Liu, G.; Zhang, D.; Sun, H.; Zhang, J. P-glycoprotein mediated efflux limits the transport of the novel anti-Parkinson’s disease candidate drug FLZ across the physiological and PD pathological in vitro BBB models. PLoS ONE 2014, 9, e102442. [Google Scholar] [CrossRef] [PubMed]
- Thuerauf, N.; Fromm, M.F. The role of the transporter P-glycoprotein for disposition and effects of centrally acting drugs and for the pathogenesis of CNS diseases. Eur. Arch. Psychiatry Clin. Neurosci. 2006, 256, 281–286. [Google Scholar] [CrossRef]
- Kawami, M.; Yamada, Y.; Issarachot, O.; Junyaprasert, V.B.; Yumoto, R.; Takano, M. P-gp modulating effect of Azadirachta indica extract in multidrug-resistant cancer cell lines. Pharmazie 2018, 73, 104–109. [Google Scholar]
- Frohman, E.M.; Racke, M.K.; Raine, C.S. Multiple sclerosis--the plaque and its pathogenesis. N. Engl. J. Med. 2006, 354, 942–955. [Google Scholar] [CrossRef]
- Steinman, L.; Zamvil, S.S. How to successfully apply animal studies in experimental allergic encephalomyelitis to research on multiple sclerosis. Ann. Neurol. 2006, 60, 12–21. [Google Scholar] [CrossRef]
- Hirst, C.; Swingler, R.; Compston, D.A.; Ben-Shlomo, Y.; Robertson, N.P. Survival and cause of death in multiple sclerosis: A prospective population-based study. J. Neurol. Neurosurg. Psychiatry 2008, 79, 1016–1021. [Google Scholar] [CrossRef] [PubMed]
- Furlan, R.; Cuomo, C.; Martino, G. Animal models of multiple sclerosis. Methods Mol. Biol. 2009, 549, 157–173. [Google Scholar] [PubMed]
- Chan, A.; Gold, R.; von Ahsen, N. ATP-binding cassette transporters in inflammatory brain disease. Curr. Pharm. Des. 2011, 17, 2803–2807. [Google Scholar] [CrossRef] [PubMed]
- Kooij, G.; van Horssen, J.; de Lange, E.C.; Reijerkerk, A.; van der Pol, S.M.; van Het Hof, B.; Drexhage, J.; Vennegoor, A.; Killestein, J.; Scheffer, G.; et al. T lymphocytes impair P-glycoprotein function during neuroinflammation. J. Autoimmun. 2010, 34, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Chinese Pharmacopoeia Committee. Chin Pharmacopoeia; People’s Medical Publishing House: Beijing, China, 1963. (In Chinese) [Google Scholar]
- Wu, H.; Gao, Y.; Shi, H.L.; Qin, L.Y.; Huang, F.; Lan, Y.Y.; Zhang, B.B.; Hu, Z.B.; Wu, X.J. Astragaloside IV improves lipid metabolism in obese mice by alleviation of leptin resistance and regulation of thermogenic network. Sci. Rep. 2016, 6, 30190. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Du, M.; Gao, Y.; Liu, H.; Wang, H.; Wu, X.; Wang, Z. Astragaloside IV attenuates experimental autoimmune encephalomyelitis of mice by counteracting oxidative stress at multiple levels. PLoS ONE 2013, 8, e76495. [Google Scholar] [CrossRef]
- Sun, B.; Rui, R.; Pan, H.; Zhang, L.; Wang, X. Effect of Combined Use of Astragaloside IV (AsIV) and Atorvastatin (AV) on Expression of PPAR-γ and Inflammation- Associated Cytokines in Atherosclerosis Rats. Med. Sci. Monit. 2018, 24, 6229–6236. [Google Scholar] [CrossRef]
- Jiang, P.; Ma, D.; Wang, X.; Wang, Y.; Bi, Y.; Yang, J.; Wang, X.; Li, X. Astragaloside IV Prevents Obesity-Associated Hypertension by Improving Pro-Inflammatory Reaction and Leptin Resistance. Mol. Cells 2018, 41, 244–255. [Google Scholar]
- Chen, Q.; Su, Y.; Ju, Y.; Ma, K.; Li, W.; Li, W. Astragalosides IV protected the renal tubular epithelial cells from free fatty acids-induced injury by reducing oxidative stress and apoptosis. Biomed. Pharmacother. 2018, 108, 679–686. [Google Scholar] [CrossRef]
- Ji, C.; Luo, Y.; Zou, C.; Huang, L.; Tian, R.; Lu, Z. Effect of astragaloside IV on indoxyl sulfate-induced kidney injury in mice via attenuation of oxidative stress. BMC Pharmacol. Toxicol. 2018, 19, 53. [Google Scholar] [CrossRef]
- Huang, F.; Lan, Y.; Qin, L.; Dong, H.; Shi, H.; Wu, H.; Zou, Q.; Hu, Z.; Wu, X. Astragaloside IV Promotes Adult Neurogenesis in Hippocampal Dentate Gyrus of Mouse through CXCL1/CXCR2 Signaling. Molecules 2018, 23, 2178. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.; Chang, H.; Zhang, J.; Guo, G.; Shi, J.; Xu, H. Astragaloside IV Protects Rat Cardiomyocytes from Hypoxia-Induced Injury by Down-Regulation of miR-23a and miR-92a. Cell Physiol. Biochem. 2018, 49, 2240–2253. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Gao, Y.; Tian, N.; Zhu, Z.; Wang, T.; Xu, J.; Wu, B.; Zhang, N. Astragaloside IV represses high glucose-induced mesangial cells activation by enhancing autophagy via SIRT1 deacetylation of NF-κB p65 subunit. Drug Des. Devel. Ther. 2018, 12, 2971–2980. [Google Scholar] [CrossRef] [PubMed]
- Costa, I.M.; Lima, F.O.V.; Fernandes, L.C.B.; Norrara, B.; Neta, F.I.; Alves, R.D.; Cavalcanti, J.R.L.P.; Lucena, E.E.S.; Cavalcante, J.S.; Rego, A.C.M.; et al. Astragaloside IV supplementation promotes a neuroprotective effect in experimental models of neurological disorders: A systematic review. Curr. Neuropharmacol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Cui, W.Q.; Wei, Y.; Cui, J.; Qiu, J.; Hu, L.L.; Gong, W.Y.; Dong, J.C.; Liu, B.J. Astragaloside IV inhibits lung cancer progression and metastasis by modulating macrophage polarization through AMPK signaling. J. Exp. Clin. Cancer Res. 2018, 37, 207. [Google Scholar] [CrossRef] [PubMed]
- Song, M.T.; Ruan, J.; Zhang, R.Y.; Deng, J.; Ma, Z.Q.; Ma, S.P. Astragaloside IV ameliorates neuroinflammation-induced depressive-like behaviors in mice via the PPARγ/NF-κB/NLRP3 inflammasome axis. Acta. Pharmacol. Sin. 2018. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.S.; Shi, H.L.; Huang, F.; Peterson, K.E.; Wu, H.; Lan, Y.Y.; Zhang, B.B.; He, Y.X.; Woods, T.; Du, M.; et al. Astragaloside IV inhibits microglia activation via glucocorticoid receptor mediated signaling pathway. Sci. Rep. 2016, 6, 19137. [Google Scholar] [CrossRef]
- Hermann, D.M.; Bassetti, C.L. Implications of ATP-binding cassette transporters for brain pharmacotherapies. Trends Pharmacol. Sci. 2007, 28, 128–134. [Google Scholar] [CrossRef]
- Jablonski, M.R.; Markandaiah, S.S.; Jacob, D.; Meng, N.J.; Li, K.; Gennaro, V.; Lepore, A.C.; Trotti, D.; Pasinelli, P. Inhibiting drug efflux transporters improves efficacy of ALS therapeutics. Ann. Clin. Transl. Neurol. 2014, 1, 996–1005. [Google Scholar] [CrossRef]
- Lee, C.G.; Tang, K.; Cheung, Y.B.; Wong, L.P.; Tan, C.; Shen, H.; Zhao, Y.; Pavanni, R.; Lee, E.J.; Wong, M.C.; et al. MDR1, the blood-brain barrier transporter, is associated with Parkinson’s disease in ethnic Chinese. J. Med. Genet. 2004, 41, e60. [Google Scholar] [CrossRef]
- Droździk, M.; Białecka, M.; Myśliwiec, K.; Honczarenko, K.; Stankiewicz, J.; Sych, Z. Polymorphism in the P-glycoprotein drug transporter MDR1 gene: A possible link between environmental and genetic factors in Parkinson’s disease. Pharmacogenetics 2003, 13, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Kortekaas, R.; Leenders, K.L.; van Oostrom, J.C.; Vaalburg, W.; Bart, J.; Willemsen, A.T.; Hendrikse, N.H. Blood-brain barrier dysfunction in parkinsonian midbrain in vivo. Ann. Neurol. 2005, 57, 176–179. [Google Scholar] [CrossRef] [PubMed]
- Kooij, G.; Backer, R.; Koning, J.J.; Reijerkerk, A.; van Horssen, J.; van der Pol, S.M.; Drexhage, J.; Schinkel, A.; Dijkstra, C.D.; den Haan, J.M.; et al. P-glycoprotein acts as an immunomodulator during neuroinflammation. PLoS ONE 2009, 4, e8212. [Google Scholar] [CrossRef] [PubMed]
- Löscher, W.; Potschka, H. Role of drug efflux transporters in the brain for drug disposition and treatment of brain diseases. Prog. Neurobiol. 76, 22–76. [CrossRef] [PubMed]
- Löscher, W.; Potschka, H. Drug resistance in brain diseases and the role of drug efflux transporters. Nat. Rev. Neurosci. 2005, 6, 591–602. [Google Scholar] [CrossRef] [PubMed]
- Lou, Y.; Guo, Z.; Zhu, Y.; Zhang, G.; Wang, Y.; Qi, X.; Lu, L.; Liu, Z.; Wu, J. Astragali radix and its main bioactive compounds activate the Nrf2-mediated signaling pathway to induce P-glycoprotein and breast cancer resistance protein. J. Ethnopharmacol. 2018, 228, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Peiris, M.; Monteith, G.R.; Roberts-Thomson, S.J.; Cabot, P.J. A model of experimental autoimmune encephalomyelitis (EAE) in C57BL/6 mice for the characterisation of intervention therapies. J. Neurosci. Methods 2007, 163, 245–254. [Google Scholar] [CrossRef]
- Yu, C.; Kastin, A.J.; Tu, H.; Waters, S.; Pan, W. TNF activates P-glycoprotein in cerebral microvascular endothelial cells. Cell Physiol. Biochem. 2007, 20, 853–858. [Google Scholar] [CrossRef]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef]
- Molecular Operating Environment (MOE), 2016.08; Chemical Computing Group Inc.: Montreal, QC, Canada, 2016.
- Bolton, E.E.; Wang, Y.; Thiessen, P.A.; Bryant, S.H. PubChem: Integrated platform of small molecules and biological activities. Annu. Rep. Comput. Chem. 2008, 4, 217–241. [Google Scholar]
Sample Availability: Samples of the compounds are not available from the authors. |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Liu, M.; Yang, L.; Huang, F.; Lan, Y.; Li, H.; Wu, H.; Zhang, B.; Shi, H.; Wu, X. P-glycoprotein Inhibitor Tariquidar Potentiates Efficacy of Astragaloside IV in Experimental Autoimmune Encephalomyelitis Mice. Molecules 2019, 24, 561. https://doi.org/10.3390/molecules24030561
Zhang W, Liu M, Yang L, Huang F, Lan Y, Li H, Wu H, Zhang B, Shi H, Wu X. P-glycoprotein Inhibitor Tariquidar Potentiates Efficacy of Astragaloside IV in Experimental Autoimmune Encephalomyelitis Mice. Molecules. 2019; 24(3):561. https://doi.org/10.3390/molecules24030561
Chicago/Turabian StyleZhang, Wei, Mei Liu, Liu Yang, Fei Huang, Yunyi Lan, Hongli Li, Hui Wu, Beibei Zhang, Hailian Shi, and Xiaojun Wu. 2019. "P-glycoprotein Inhibitor Tariquidar Potentiates Efficacy of Astragaloside IV in Experimental Autoimmune Encephalomyelitis Mice" Molecules 24, no. 3: 561. https://doi.org/10.3390/molecules24030561
APA StyleZhang, W., Liu, M., Yang, L., Huang, F., Lan, Y., Li, H., Wu, H., Zhang, B., Shi, H., & Wu, X. (2019). P-glycoprotein Inhibitor Tariquidar Potentiates Efficacy of Astragaloside IV in Experimental Autoimmune Encephalomyelitis Mice. Molecules, 24(3), 561. https://doi.org/10.3390/molecules24030561