Identification of a Novel Gene Encoding the Specialized Alanine Decarboxylase in Tea (Camellia sinensis) Plants


Abstract
1. Introduction
2. Results and Discussion
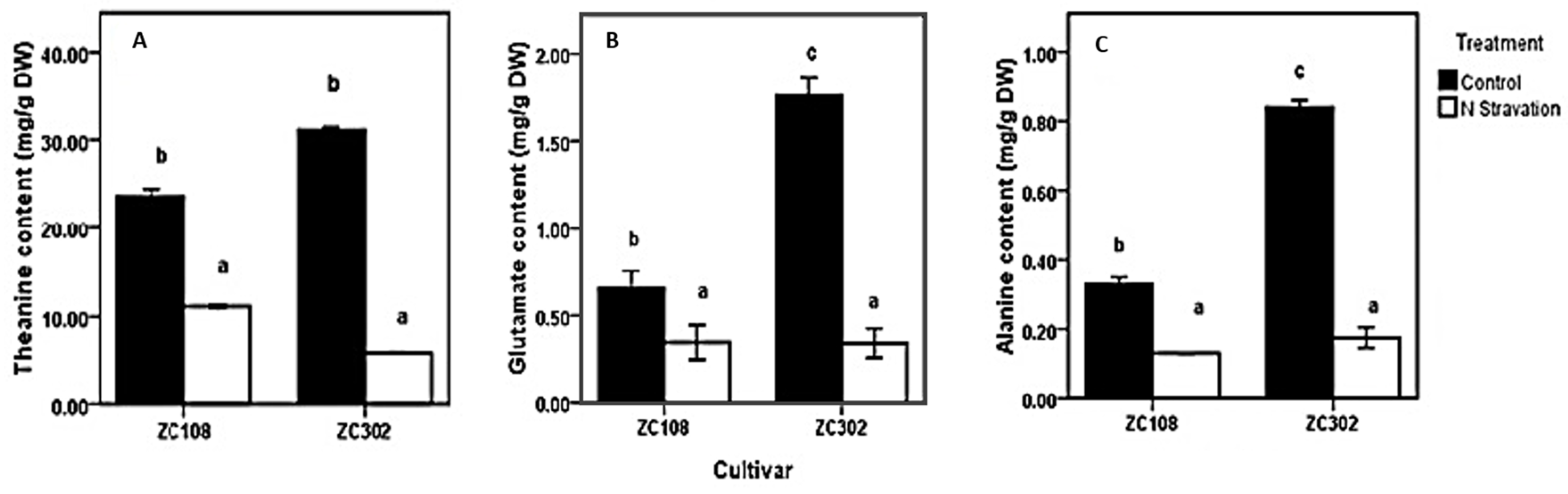
2.1. Effects of Nitrogen Starvation on Content of Free Amino Acids in Tea Roots
2.2. Identification of a Candidate Gene of CsAlaDC
2.3. Cloning of pCsAlaDC from Camellia sinensis
2.4. CsAlaDC Gene Was Expressed at a High Level in the Roots Where Theanine Was Synthesized
3. Materials and Methods
3.1. Plant Materials and Growth Conditions
3.2. Determination of Free Amino-Acid Content
3.3. RNA Extraction
3.4. cDNA Library Preparation and Illumina Sequencing
3.5. De Novo Assembly of Transcriptome and Analysis of Illumina Sequencing Results
3.6. Functional Annotation and Classification
3.7. Identification of Differentially Expressed Genes
3.8. Isolation of pCsAlaDC and Sequence Analysis
3.9. Heterologous Expression and Purification of the Recombinant Protein
3.10. Measurement of Enzymatic Activity
3.11. Relative Quantification by Real-Time PCR
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Chen, L.; Zhou, Z.-X.; Yang, Y.-J. Genetic improvement and breeding of tea plant (Camellia sinensis) in China: From individual selection to hybridization and molecular breeding. Euphytica 2007, 154, 239–248. [Google Scholar] [CrossRef]
- Zhang, H.-B.; Xia, E.-H.; Huang, H.; Jiang, J.-J.; Liu, B.-Y.; Gao, L.-Z. De novo transcriptome assembly of the wild relative of tea tree (Camellia taliensis) and comparative analysis with tea transcriptome identified putative genes associated with tea quality and stress response. BMC Genom. 2015, 16, 298. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Baldermann, S.; Watanabe, N. Recent studies of the volatile compounds in tea. Food Res. Int. 2013, 53, 585–599. [Google Scholar] [CrossRef]
- Liang, Y.R.; Ma, W.Y.; Lu, J.L.; Wu, Y. Comparison of chemical compositions of Ilex latifolia thumb and Camellia sinensis L. Food Chem. 2001, 75, 339–343. [Google Scholar] [CrossRef]
- Mamati, G.E.; Liang, Y.R.; Lu, J.L. Expression of basic genes involved in tea polyphenol synthesis in relation to accumulation of catechins and total tea polyphenols. J. Sci. Food Agric. 2006, 86, 459–464. [Google Scholar] [CrossRef]
- Sakato, Y. The chemical constituents of tea: III. A new amide theanine. Nippon Nogeikagaku Kaishi 1949, 23, 262–267. [Google Scholar] [CrossRef]
- Yamaguchi, S.; Ninomiya, K. Umami and food palatability. J. Nutr. 2000, 130, 921S–926S. [Google Scholar] [CrossRef]
- Oh, K.; Kato, T.; Xu, H.L. Transport of nitrogen assimilation in xylem vessels of green tea plants fed with NH(4)-N and NO(3)-N. Pedosphere 2008, 18, 222–226. [Google Scholar] [CrossRef]
- Egashira, N.; Hayakawa, K.; Osajima, M.; Mishima, K.; Iwasaki, K.; Oishi, R.; Fujiwara, M. Involvement of GABA(A)-receptors in the neuroprotective effect of theanine on focal cerebral ischemia in mice. J. Pharm. Sci. 2007, 105, 211–214. [Google Scholar] [CrossRef]
- Haskell, C.F.; Kennedy, D.O.; Milne, A.L.; Wesnes, K.A.; Scholey, A.B. The effects of L-theanine, caffeine and their combination on cognition and mood. Biol. Psychol. 2008, 77, 113–122. [Google Scholar] [CrossRef]
- Kakuda, T.; Yanase, H.; Utsunomiya, K.; Nozawa, A.; Unno, T.; Kataoka, K. Protective effect of gamma-glutamylethylamide (theanine) on ischemic delayed neuronal death in gerbils. Neurosci. Lett. 2000, 289, 189–192. [Google Scholar] [CrossRef]
- Liu, Q.; Duan, H.; Luan, J.; Yagasaki, K.; Zhang, G. Effects of theanine on growth of human lung cancer and leukemia cells as well as migration and invasion of human lung cancer cells. Cytotechnology 2009, 59, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.; Gray, M.A.; Oliver, C.; Liley, D.T.; Harrison, B.J.; Bartholomeusz, C.F.; Phan, K.L.; Nathan, P.J. The acute effects of L-theanine in comparison with alprazolam on anticipatory anxiety in humans. Hum. Psychopharm. Clin. 2004, 19, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Rogers, P.J.; Smith, J.E.; Heatherley, S.V.; Pleydell-Pearce, C. Time for tea: Mood, blood pressure and cognitive performance effects of caffeine and theanine administered alone and together. Psychopharmacology 2008, 195, 569. [Google Scholar] [CrossRef] [PubMed]
- Takagi, Y.; Kurihara, S.; Higashi, N.; Morikawa, S.; Kase, T.; Maeda, A.; Arisaka, H.; Shibahara, S.; Akiyama, Y. Combined Administration of L-Cystine and L-Theanine Enhances Immune Functions and Protects against Influenza Virus Infection in Aged Mice. J. Vet. Med. Sci. 2010, 72, 157–165. [Google Scholar] [CrossRef]
- Zheng, G.D.; Sayama, K.; Okubo, T.; Juneja, L.R.; Oguni, I. Anti-obesity effects of three major components of green tea, catechins, caffeine and theanine, in mice. In Vivo 2004, 18, 55–62. [Google Scholar]
- Cartwright, R.; Roberts, E.; Wood, D. Theanine, an amino-acid n-ethyl amide present in tea. J. Sci. Food Agric. 1954, 5, 597–599. [Google Scholar] [CrossRef]
- Finger, A.; Kuhr, S.; Engelhardt, U.H. Chromatography of tea constituents. J. Chromatogr. A 1992, 624, 293–315. [Google Scholar] [CrossRef]
- Konishi, S.; Takahashi, E. Existence and synthesis of L-glutamic acid γ-methylamide in tea plants. Plant Cell Physiol. 1966, 7, 171–175. [Google Scholar] [CrossRef]
- Millin, D.J.; Crispin, D.J.; Swaine, D. Nonvolatile components of black tea and their contribution to the character of the beverage. J. Agric. Food Chem. 1969, 17, 717–722. [Google Scholar] [CrossRef]
- Wickremasinghe, R.L. Tea. In Advances in Food Research; Academic Press: London, UK, 1978; Volume 24, pp. 229–286. [Google Scholar]
- Feldheim, W.; Yongvanit, P.; Cummings, P.H. Investigation of the presence and significance of theanine in the tea plant. J. Sci. Food Agric. 1986, 37, 527–534. [Google Scholar] [CrossRef]
- Deng, W.-W.; Ogita, S.; Ashihara, H. Biosynthesis of theanine (γ-ethylamino-l-glutamic acid) in seedlings of Camellia sinensis. Phytochem. Lett. 2008, 1, 115–119. [Google Scholar] [CrossRef]
- Deng, W.-W.; Ogita, S.; Ashihara, H. Ethylamine Content and Theanine Biosynthesis in Different Organs of Camellia sinensis Seedlings. Z. Nat. C 2009, 64, 387–390. [Google Scholar] [CrossRef]
- Sasaoka, K.; Kito, M. Synthesis of Theanine by Tea Seedling Homogenate. Agric. Biol. Chem. 1964, 28, 313–317. [Google Scholar] [CrossRef]
- Takeo, T. L-Alanine as a precursor of ethylamine in Camellia sinensis. Phytochemistry 1974, 13, 1401–1406. [Google Scholar] [CrossRef]
- Ruan, J.; Ma, L.; Yang, Y. Magnesium nutrition on accumulation and transport of amino acids in tea plants. J. Sci. Food Agric. 2012, 92, 1375–1383. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Yan, J.; Cui, J.; Mao, S.; Li, M.; Liao, X.; Tong, H. Dynamic changes in amino acids, catechins, caffeine and gallic acid in green tea during withering. J. Food Compos. Anal. 2018, 66, 98–108. [Google Scholar] [CrossRef]
- Tsushida, T.; Takeo, T. An enzyme hydrolyzing L-theanine in tea leaves. Agric. Biol. Chem. 1985, 49, 2913–2917. [Google Scholar] [CrossRef][Green Version]
- Kito, M.; Kokura, H.; Izaki, J.; Sasaoka, K. Theanine, a precursor of the phloroglucinol nucleus of catechins in tea plants. Phytochemistry 1968, 7, 599–603. [Google Scholar] [CrossRef]
- Ashihara, H. Occurrence, Biosynthesis and Metabolism of Theanine (gamma-Glutamyl-L-ethylamide) in Plants: A Comprehensive Review. Nat. Prod. Commun. 2015, 10, 803–810. [Google Scholar]
- Lea, P.J.; Black, R.D.; Chen, F.L.; Hecht, U. Enzymes of ammonia assimilation. In Meth Plant Biochemistry; Academic Press: London, UK, 1990; Volume 3, pp. 257–276. [Google Scholar]
- Miflin, B.J.; Lea, P.J. The pathway of nitrogen assimilation in plants. Phytochemistry 1976, 15, 873–885. [Google Scholar] [CrossRef]
- Crocomo, O.; Fowden, L. Amino acid decarboxylases of higher plants: The formation of ethylamine. Phytochemistry 1970, 9, 537–540. [Google Scholar] [CrossRef]
- Smith, T. The occurrence, metabolism and functions of amines in plants. Biol. Rev. 1971, 46, 201–241. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Fu, X.; Wang, X.; Liao, Y.; Zeng, L.; Dong, F.; Yang, Z. Studies on the Biochemical Formation Pathway of the Amino Acid l-Theanine in Tea (Camellia sinensis) and Other Plants. J. Agric. Food Chem. 2017, 65, 7210–7216. [Google Scholar] [CrossRef] [PubMed]
- Sasaoka, K.; Kito, M.; Inagaki, H. Studies on the Biosynthesis of Theanine in Tea Seedlings Synthesis of Theanine by the Homogenate of Tea Seedlings. Agric. Biol. Chem. 1963, 27, 467–468. [Google Scholar] [CrossRef]
- Takeo, T. L-alanine decarboxylase in Camellia sinensis. Phytochemistry 1978, 17, 313–314. [Google Scholar] [CrossRef]
- Hsieh, P.H.; Kan, C.C.; Wu, H.Y.; Yang, H.C.; Hsieh, M.H. Early molecular events associated with nitrogen deficiency in rice seedling roots. Sci. Rep. 2018, 8, 12207. [Google Scholar] [CrossRef]
- Kant, S.; Bi, Y.M.; Weretilnyk, E.; Barak, S.; Rothstein, S.J. The Arabidopsis halophytic relative Thellungiella halophila tolerates nitrogen-limiting conditions by maintaining growth, nitrogen uptake, and assimilation. Plant Physiol. 2008, 147, 1168–1180. [Google Scholar] [CrossRef]
- Lemaitre, T.; Gaufichon, L.; Boutet-Mercey, S.; Christ, A.; Masclaux-Daubresse, C. Enzymatic and metabolic diagnostic of nitrogen deficiency in Arabidopsis thaliana Wassileskija accession. Plant Cell Physiol. 2008, 49, 1056–1065. [Google Scholar] [CrossRef]
- Ruan, L.; Wang, L.; Wei, K.; Cheng, H.; Li, H.; Shao, S.; Wu, L. Comparative analysis of nitrogen spatial heterogeneity responses in low nitrogen susceptible and tolerant tea plants (Camellia sinensis). Sci. Hortic. 2019, 246, 182–189. [Google Scholar] [CrossRef]
- Ruan, J.; Gerendás, J.; Härdter, R.; Sattelmacher, B. Effect of root zone pH and form and concentration of nitrogen on accumulation of quality-related components in green tea. J. Sci. Food Agric. 2007, 87, 1505–1516. [Google Scholar] [CrossRef]
- Kumar, R. Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes. Front. Plant Sci. 2016, 7, 1268. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Thornton, J.M.; Todd, A.E.; Milburn, D.; Borkakoti, N.; Orengo, C.A. From structure to function: Approaches and limitations. Nat. Struct. Biol. 2000, 7, 991–994. [Google Scholar] [CrossRef] [PubMed]
- Rontein, D.; Rhodes, D.; Hanson, A.D. Evidence from Engineering that Decarboxylation of Free Serine is the Major Source of Ethanolamine Moieties in Plants. Plant Cell Physiol. 2003, 44, 1185–1191. [Google Scholar] [CrossRef] [PubMed]
- Torrens-Spence, M.P.; Liu, P.Y.; Ding, H.Z.; Harich, K.; Gillaspy, G.; Li, J.Y. Biochemical evaluation of the decarboxylation and decarboxylation-deamination activities of plant aromatic amino acid decarboxylases. J. Biol. Chem. 2013, 288, 2376–2387. [Google Scholar] [CrossRef] [PubMed]
- Bunsupa, S.; Hanada, K.; Maruyama, A.; Aoyagi, K.; Komatsu, K.; Ueno, H.; Yamashita, M.; Sasaki, R.; Oikawa, A.; Saito, K.; et al. Molecular Evolution and Functional Characterization of a Bifunctional Decarboxylase Involved in Lycopodium Alkaloid Biosynthesis. Plant Physiol. 2016, 171, 2432–2444. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.-W.; Ogita, S.; Ashihara, H. Distribution and biosynthesis of theanine in Theaceae plants. Plant Physiol. Biochem. 2010, 48, 70–72. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Deant, M.A.; Brunet, F.; Long, M. Evolving protein functional diversity in new genes of Drosophila. Proc. Natl. Acad. Sci. USA 2004, 101, 16246–16250. [Google Scholar] [CrossRef]
- Salazar, C.; Armenta, J.M.; Cortes, D.F.; Shulaev, V. Combination of an AccQ.Tag-ultra performance liquid chromatographic method with tandem mass spectrometry for the analysis of amino acids. Methods Mol. Biol. 2012, 828, 13–28. [Google Scholar] [CrossRef]
- Armenta, J.M.; Cortes, D.F.; Pisciotta, J.M.; Shuman, J.L.; Blakeslee, K.; Rasoloson, D.; Ogunbiyi, O.; Sullivan, D.J.; Shulaev, V. Sensitive and Rapid Method for Amino Acid Quantitation in Malaria Biological Samples Using AccQ•Tag Ultra Performance Liquid Chromatography-Electrospray Ionization-MS/MS with Multiple Reaction Monitoring. Anal. Chem. 2010, 82, 548–558. [Google Scholar] [CrossRef]
- Jiang, X.; Li, W.; Yang, P.; Shi, X.; Guo, J.; Wang, X.; Zhang, Y.; Peng, H. Preliminary report on introduction experiment of new tea cultivars in Jiangxi province. J. South Agric. 2015, 46, 1856–1861. [Google Scholar]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644. [Google Scholar] [CrossRef] [PubMed]
- Pertea, G.; Huang, X.; Liang, F.; Antonescu, V.; Sultana, R.; Karamycheva, S.; Lee, Y.; White, J.; Cheung, F.; Parvizi, B. TIGR Gene Indices clustering tools (TGICL): A software system for fast clustering of large EST datasets. Bioinformatics 2003, 19, 651–652. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Zhongcha 302 | Zhongcha 108 | |
|---|---|---|
| Unique gene detected | 91,650 | 98,406 |
| Significantly changed genes | 7985 | 17,824 |
| Upregulated | 3288 | 15,785 |
| Downregulated | 4697 | 2039 |
| Shared genes (upregulated) | 742 | |
| Shared genes (downregulated) | 128 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, P.; Wei, K.; Wang, L.; Zhang, F.; Ruan, L.; Li, H.; Wu, L.; Cheng, H. Identification of a Novel Gene Encoding the Specialized Alanine Decarboxylase in Tea (Camellia sinensis) Plants. Molecules 2019, 24, 540. https://doi.org/10.3390/molecules24030540
Bai P, Wei K, Wang L, Zhang F, Ruan L, Li H, Wu L, Cheng H. Identification of a Novel Gene Encoding the Specialized Alanine Decarboxylase in Tea (Camellia sinensis) Plants. Molecules. 2019; 24(3):540. https://doi.org/10.3390/molecules24030540
Chicago/Turabian StyleBai, Peixian, Kang Wei, Liyuan Wang, Fen Zhang, Li Ruan, Hailin Li, Liyun Wu, and Hao Cheng. 2019. "Identification of a Novel Gene Encoding the Specialized Alanine Decarboxylase in Tea (Camellia sinensis) Plants" Molecules 24, no. 3: 540. https://doi.org/10.3390/molecules24030540
APA StyleBai, P., Wei, K., Wang, L., Zhang, F., Ruan, L., Li, H., Wu, L., & Cheng, H. (2019). Identification of a Novel Gene Encoding the Specialized Alanine Decarboxylase in Tea (Camellia sinensis) Plants. Molecules, 24(3), 540. https://doi.org/10.3390/molecules24030540