4-O-Methylhonokiol Influences Normal Cardiovascular Development in Medaka Embryo



 , ,
, ,
Abstract
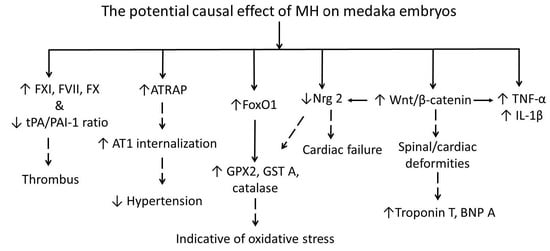
1. Introduction
2. Results
2.1. Toxicity of MH in Medaka Embryo
2.2. Exposure to MH during Early Development Affects Cardiovascular Structure and Function
2.3. MH Is Implicated in Cardiovascular Dysfunction
2.4. MH Possesses the Proinflammatory and Pro-Oxidative Properties
2.5. MH Reduces the Normal Hatching Process of Medaka Embryos
2.6. MH Regulates Wnt/β-Catenin Pathway during Cardiomyogenesis
2.7. MH Prolongs Swimming Duration
3. Discussion
4. Materials and Methods
4.1. Medaka Maintenance and Breeding
4.2. Extraction and Isolation of MH
4.3. MH Treatment
4.4. Determination of MH Purity and MH Absorption in Medaka Embryos
4.5. Microscopy Study
4.6. Locomotion Study
4.7. RNA Extraction
4.8. Reverse Transcription—Quantitative Polymerase Chain Reaction (RT-qPCR)
4.9. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ATRAP | AT II type 1 receptor-associated protein |
| BNP A | Brain natriuretic peptide A |
| CAT | Catalase |
| DKK1 | Dickkopf 1 |
| dpf | Days post fertilization |
| dph | Days post hatching |
| Dvl | Dishevelled |
| eEf1α | Eukaryotic elongation factor 1 alpha |
| ErbB | Erythroblastic leukemia viral oncogene homolog |
| ETB | Endothelin receptor B |
| FoxO1 | Forkhead box O1 |
| Fzd | Frizzled |
| GPX 2 | Glutathione peroxidase 2 |
| GSK-3β | Glycogen synthase kinase 3 beta |
| GST A | Glutathione S-transferase A |
| hpf | Hours post fertilization |
| LRP5 | Low-density lipoprotein receptor-related protein 5 |
| MH | 4-O-methylhonokiol |
| MI | Myocardial infarction |
| Nrg | Neuregulin |
| PAI-1 | Plasminogen activator inhibitor 1 |
| SOD2 | Superoxide dismutase 2 |
| STEMI | ST-segment elevation MI |
| tPA | Tissue plasminogen activator |
| UHPLC/MS | Ultra-high performance liquid chromatography/mass spectrometer |
| u-PA | Urokinase plasminogen activator |
| uPAR | Urokinase plasminogen activator receptor |
Appendix A

Appendix B
References
- Li, N.; Song, Y.; Zhang, W.; Wang, W.; Chen, J.; Wong, A.W.; Roberts, A. Evaluation of the in vitro and in vivo genotoxicity of magnolia bark extract. Regul. Toxicol. Pharmacol. 2007, 49, 154–159. [Google Scholar] [CrossRef]
- Shen, J.L.; Man, K.M.; Huang, P.H.; Chen, W.C.; Chen, D.C.; Cheng, Y.W.; Liu, P.L.; Chou, M.C.; Chen, Y.H. Honokiol and magnolol as multifunctional antioxidative molecules for dermatologic disorders. Molecules 2010, 15, 6452–6465. [Google Scholar] [CrossRef] [PubMed]
- Chicca, A.; Gachet, M.S.; Petrucci, V.; Schuehly, W.; Charles, R.P.; Gertsch, J. 4′-O-methylhonokiol increases levels of 2-arachidonoyl glycerol in mouse brain via selective inhibition of its COX-2-mediated oxygenation. J. Neuroinflamm. 2015, 12, 89. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, F.; Chavez, M.; Garduno-Ramirez, M.L.; Chavez-Avila, V.M.; Mata, M.; Cruz-Sosa, F. Honokiol and magnolol production by in vitro micropropagated plants of Magnolia dealbata, an endangered endemic Mexican species. Nat. Prod. Commun. 2010, 5, 235–240. [Google Scholar] [PubMed]
- Schuhly, W.; Khan, S.I.; Fischer, N.H. Neolignans from North American Magnolia species with cyclooxygenase 2 inhibitory activity. Inflammopharmacology 2009, 17, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Bernaskova, M.; Schoeffmann, A.; Schuehly, W.; Hufner, A.; Baburin, I.; Hering, S. Nitrogenated honokiol derivatives allosterically modulate GABAA receptors and act as strong partial agonists. Bioorg. Med. Chem. 2015, 23, 6757–6762. [Google Scholar] [CrossRef]
- Lin, C.F.; Hwang, T.L.; Al-Suwayeh, S.A.; Huang, Y.L.; Hung, Y.Y.; Fang, J.Y. Maximizing dermal targeting and minimizing transdermal penetration by magnolol/honokiol methoxylation. Int. J. Pharm. 2013, 445, 153–162. [Google Scholar] [CrossRef]
- Fang, J.Y.; Huang, T.H.; Hung, C.F.; Huang, Y.L.; Aljuffali, I.A.; Liao, W.C.; Lin, C.F. Derivatization of honokiol by integrated acetylation and methylation for improved cutaneous delivery and anti-inflammatory potency. Eur. J. Pharm. Sci. 2018, 114, 189–198. [Google Scholar] [CrossRef]
- Kim, H.S.; Ryu, H.S.; Kim, J.S.; Kim, Y.G.; Lee, H.K.; Jung, J.K.; Kwak, Y.S.; Lee, K.; Seo, S.Y.; Yun, J.; et al. Validation of cyclooxygenase-2 as a direct anti-inflammatory target of 4-O-methylhonokiol in zymosan-induced animal models. Arch. Pharm. Res. 2015, 38, 813–825. [Google Scholar] [CrossRef]
- Chiu, J.H.; Ho, C.T.; Wei, Y.H.; Lui, W.Y.; Hong, C.Y. In vitro and in vivo protective effect of honokiol on rat liver from peroxidative injury. Life Sci. 1997, 61, 1961–1971. [Google Scholar] [CrossRef]
- Huang, L.; Zhang, K.; Guo, Y.; Huang, F.; Yang, K.; Chen, L.; Huang, K.; Zhang, F.; Long, Q.; Yang, Q. Honokiol protects against doxorubicin cardiotoxicity via improving mitochondrial function in mouse hearts. Sci. Rep. 2017, 7, 11989. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Choi, D.Y.; Lee, Y.K.; Lee, Y.M.; Han, S.B.; Kim, Y.H.; Kim, K.H.; Nam, S.Y.; Lee, B.J.; Kang, J.K.; et al. 4-O-methylhonokiol prevents memory impairment in the Tg2576 transgenic mice model of Alzheimer’s disease via regulation of beta-secretase activity. J. Alzheimer’s Dis. 2012, 29, 677–690. [Google Scholar] [CrossRef] [PubMed]
- Bunel, V.; Antoine, M.H.; Stevigny, C.; Nortier, J.; Duez, P. New in vitro insights on a cell death pathway induced by magnolol and honokiol in aristolochic acid tubulotoxicity. Food Chem. Toxicol. 2016, 87, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, M.; Urnezis, P.; Tian, M. Compressed mints and chewing gum containing magnolia bark extract are effective against bacteria responsible for oral malodor. J. Agric. Food Chem. 2007, 55, 9465–9469. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.T.; Chi, C.W. Emerging role of the peroxisome proliferator-activated receptor-gamma in hepatocellular carcinoma. J. Hepatocell. Carcinoma 2014, 1, 127–135. [Google Scholar] [PubMed]
- Rijken, D.C.; Hoylaerts, M.; Collen, D. Fibrinolytic properties of one-chain and two-chain human extrinsic (tissue-type) plasminogen activator. J. Biol. Chem. 1982, 257, 2920–2925. [Google Scholar] [PubMed]
- Kobayashi, R.; Wakui, H.; Azushima, K.; Uneda, K.; Haku, S.; Ohki, K.; Haruhara, K.; Kinguchi, S.; Matsuda, M.; Ohsawa, M.; et al. An angiotensin II type 1 receptor binding molecule has a critical role in hypertension in a chronic kidney disease model. Kidney Int. 2017, 91, 1115–1125. [Google Scholar] [CrossRef]
- Wakui, H.; Dejima, T.; Tamura, K.; Uneda, K.; Azuma, K.; Maeda, A.; Ohsawa, M.; Kanaoka, T.; Azushima, K.; Kobayashi, R.; et al. Activation of angiotensin II type 1 receptor-associated protein exerts an inhibitory effect on vascular hypertrophy and oxidative stress in angiotensin II-mediated hypertension. Cardiovasc. Res. 2013, 100, 511–519. [Google Scholar] [CrossRef]
- Ford, B.D.; Liu, Y.; Mann, M.A.; Krauss, R.; Phillips, K.; Gan, L.; Fischbach, G.D. Neuregulin-1 suppresses muscarinic receptor expression and acetylcholine-activated muscarinic K+ channels in cardiac myocytes. Biochem. Biophys. Res. Commun. 2003, 308, 23–28. [Google Scholar] [CrossRef]
- Petrie, T.A.; Strand, N.S.; Yang, C.T.; Rabinowitz, J.S.; Moon, R.T. Macrophages modulate adult zebrafish tail fin regeneration. Development 2014, 141, 2581–2591. [Google Scholar] [CrossRef]
- Potente, M.; Urbich, C.; Sasaki, K.; Hofmann, W.K.; Heeschen, C.; Aicher, A.; Kollipara, R.; DePinho, R.A.; Zeiher, A.M.; Dimmeler, S. Involvement of Foxo transcription factors in angiogenesis and postnatal neovascularization. J. Clin. Investig. 2005, 115, 2382–2392. [Google Scholar] [CrossRef] [PubMed]
- Hagenmaier, H.E. The hatching process in fish embryos. IV. The enzymological properties of a highly purified enzyme (chorionase) from the hatching fluid of the rainbow trout, Salmo gairdneri Rich. Comp. Biochem. Physiol. Part B Comp. Biochem. 1974, 49, 313–324. [Google Scholar] [CrossRef]
- Buikema, J.W.; Mady, A.S.; Mittal, N.V.; Atmanli, A.; Caron, L.; Doevendans, P.A.; Sluijter, J.P.; Domian, I.J. Wnt/beta-catenin signaling directs the regional expansion of first and second heart field-derived ventricular cardiomyocytes. Development 2013, 140, 4165–4176. [Google Scholar] [CrossRef]
- Bajoghli, B.; Aghaallaei, N.; Jung, G.; Czerny, T. Induction of otic structures by canonical Wnt signalling in medaka. Dev. Genes Evol. 2009, 219, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Loh, N.Y.; Neville, M.J.; Marinou, K.; Hardcastle, S.A.; Fielding, B.A.; Duncan, E.L.; McCarthy, M.I.; Tobias, J.H.; Gregson, C.L.; Karpe, F.; et al. LRP5 regulates human body fat distribution by modulating adipose progenitor biology in a dose- and depot-specific fashion. Cell Metab. 2015, 21, 262–273. [Google Scholar] [CrossRef] [PubMed]
- Ackers, I.; Malgor, R. Interrelationship of canonical and non-canonical Wnt signalling pathways in chronic metabolic diseases. Diabetes Vasc. Dis. Res. 2018, 15, 3–13. [Google Scholar] [CrossRef]
- Nguyen, G. Renin, (pro)renin and receptor: An update. Clin. Sci. 2011, 120, 169–178. [Google Scholar] [CrossRef]
- Bisson, J.A.; Mills, B.; Paul Helt, J.C.; Zwaka, T.P.; Cohen, E.D. Wnt5a and Wnt11 inhibit the canonical Wnt pathway and promote cardiac progenitor development via the Caspase-dependent degradation of AKT. Dev. Biol. 2015, 398, 80–96. [Google Scholar] [CrossRef] [PubMed]
- Ueno, S.; Weidinger, G.; Osugi, T.; Kohn, A.D.; Golob, J.L.; Pabon, L.; Reinecke, H.; Moon, R.T.; Murry, C.E. Biphasic role for Wnt/beta-catenin signaling in cardiac specification in zebrafish and embryonic stem cells. Proc. Natl. Acad. Sci. USA 2007, 104, 9685–9690. [Google Scholar] [CrossRef]
- Sklepkiewicz, P.; Shiomi, T.; Kaur, R.; Sun, J.; Kwon, S.; Mercer, B.; Bodine, P.; Schermuly, R.T.; George, I.; Schulze, P.C.; et al. Loss of secreted frizzled-related protein-1 leads to deterioration of cardiac function in mice and plays a role in human cardiomyopathy. Circ. Heart Fail. 2015, 8, 362–372. [Google Scholar] [CrossRef]
- Foulquier, S.; Daskalopoulos, E.P.; Lluri, G.; Hermans, K.C.M.; Deb, A.; Blankesteijn, W.M. WNT Signaling in Cardiac and Vascular Disease. Pharmacol. Rev. 2018, 70, 68–141. [Google Scholar] [CrossRef] [PubMed]
- Askevold, E.T.; Aukrust, P.; Nymo, S.H.; Lunde, I.G.; Kaasboll, O.J.; Aakhus, S.; Florholmen, G.; Ohm, I.K.; Strand, M.E.; Attramadal, H.; et al. The cardiokine secreted Frizzled-related protein 3, a modulator of Wnt signalling, in clinical and experimental heart failure. J. Intern. Med. 2014, 275, 621–630. [Google Scholar] [CrossRef]
- Barandon, L.; Casassus, F.; Leroux, L.; Moreau, C.; Allieres, C.; Lamaziere, J.M.; Dufourcq, P.; Couffinhal, T.; Duplaa, C. Secreted frizzled-related protein-1 improves postinfarction scar formation through a modulation of inflammatory response. Arterioscler. Thromb. Vasc. Biol. 2011, 31, e80–e87. [Google Scholar] [CrossRef] [PubMed]
- Halleskog, C.; Dijksterhuis, J.P.; Kilander, M.B.; Becerril-Ortega, J.; Villaescusa, J.C.; Lindgren, E.; Arenas, E.; Schulte, G. Heterotrimeric G protein-dependent WNT-5A signaling to ERK1/2 mediates distinct aspects of microglia proinflammatory transformation. J. Neuroinflamm. 2012, 9, 111. [Google Scholar] [CrossRef]
- Kasaai, B.; Moffatt, P.; Al-Salmi, L.; Lauzier, D.; Lessard, L.; Hamdy, R.C. Spatial and temporal localization of WNT signaling proteins in a mouse model of distraction osteogenesis. J. Histochem. Cytochem. 2012, 60, 219–228. [Google Scholar] [CrossRef]
- Tortelote, G.G.; Reis, R.R.; de Almeida Mendes, F.; Abreu, J.G. Complexity of the Wnt/betacatenin pathway: Searching for an activation model. Cell Signal. 2017, 40, 30–43. [Google Scholar] [CrossRef] [PubMed]
- Luu, H.H.; Zhang, R.; Haydon, R.C.; Rayburn, E.; Kang, Q.; Si, W.; Park, J.K.; Wang, H.; Peng, Y.; Jiang, W.; et al. Wnt/beta-catenin signaling pathway as a novel cancer drug target. Curr. Cancer Drug Targets 2004, 4, 653–671. [Google Scholar] [CrossRef]
- Dawson, K.; Aflaki, M.; Nattel, S. Role of the Wnt-Frizzled system in cardiac pathophysiology: A rapidly developing, poorly understood area with enormous potential. J. Physiol. 2013, 591, 1409–1432. [Google Scholar] [CrossRef]
- Koch, A.; Waha, A.; Hartmann, W.; Hrychyk, A.; Schuller, U.; Wharton, K.A., Jr.; Fuchs, S.Y.; von Schweinitz, D.; Pietsch, T. Elevated expression of Wnt antagonists is a common event in hepatoblastomas. Clin. Cancer Res. 2005, 11, 4295–4304. [Google Scholar] [CrossRef]
- Grainger, D.J.; Kemp, P.R.; Liu, A.C.; Lawn, R.M.; Metcalfe, J.C. Activation of transforming growth factor-beta is inhibited in transgenic apolipoprotein(a) mice. Nature 1994, 370, 460–462. [Google Scholar] [CrossRef]
- Drera, B.; Ritelli, M.; Zoppi, N.; Wischmeijer, A.; Gnoli, M.; Fattori, R.; Calzavara-Pinton, P.G.; Barlati, S.; Colombi, M. Loeys-Dietz syndrome type I and type II: Clinical findings and novel mutations in two Italian patients. Orphanet J. Rare Dis. 2009, 4, 24. [Google Scholar] [CrossRef] [PubMed]
- Senthilkumar, K.; Arunkumar, R.; Elumalai, P.; Sharmila, G.; Gunadharini, D.N.; Banudevi, S.; Krishnamoorthy, G.; Benson, C.S.; Arunakaran, J. Quercetin inhibits invasion, migration and signalling molecules involved in cell survival and proliferation of prostate cancer cell line (PC-3). Cell Biochem. Funct. 2011, 29, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Hua, J.; Xu, Y.; He, Y.; Jiang, X.; Ye, W.; Pan, Z. Wnt4/beta-catenin signaling pathway modulates balloon-injured carotid artery restenosis via disheveled-1. Int. J. Clin. Exp. Pathol. 2014, 7, 8421–8431. [Google Scholar]
- Shou, J.; Ali-Osman, F.; Multani, A.S.; Pathak, S.; Fedi, P.; Srivenugopal, K.S. Human Dkk-1, a gene encoding a Wnt antagonist, responds to DNA damage and its overexpression sensitizes brain tumor cells to apoptosis following alkylation damage of DNA. Oncogene 2002, 21, 878–889. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Gong, W.; Li, X.; Wan, R.; Mo, F.; Zhang, Z.; Huang, P.; Hu, Z.; Lai, Z.; Lu, X.; et al. Recent Progress of Wnt Pathway Inhibitor Dickkopf-1 in Liver Cancer. J. Nanosci. Nanotechnol. 2018, 18, 5192–5206. [Google Scholar] [CrossRef] [PubMed]
- Combes, A.N.; Bowles, J.; Feng, C.W.; Chiu, H.S.; Khoo, P.L.; Jackson, A.; Little, M.H.; Tam, P.P.; Koopman, P. Expression and functional analysis of Dkk1 during early gonadal development. Sex. Dev. 2011, 5, 124–130. [Google Scholar] [CrossRef]
- Foley, A.C.; Mercola, M. Heart induction by Wnt antagonists depends on the homeodomain transcription factor Hex. Genes Dev. 2005, 19, 387–396. [Google Scholar] [CrossRef]
- Peghaire, C.; Bats, M.L.; Sewduth, R.; Jeanningros, S.; Jaspard, B.; Couffinhal, T.; Duplaa, C.; Dufourcq, P. Fzd7 (Frizzled-7) Expressed by Endothelial Cells Controls Blood Vessel Formation Through Wnt/beta-Catenin Canonical Signaling. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 2369–2380. [Google Scholar] [CrossRef]
- Lattanzio, S.; Santilli, F.; Liani, R.; Vazzana, N.; Ueland, T.; Di Fulvio, P.; Formoso, G.; Consoli, A.; Aukrust, P.; Davi, G. Circulating dickkopf-1 in diabetes mellitus: Association with platelet activation and effects of improved metabolic control and low-dose aspirin. J. Am. Heart Assoc. 2014, 3, e001000. [Google Scholar] [CrossRef]
- Chen, L.; Wu, Q.; Guo, F.; Xia, B.; Zuo, J. Expression of Dishevelled-1 in wound healing after acute myocardial infarction: Possible involvement in myofibroblast proliferation and migration. J. Cell. Mol. Med. 2004, 8, 257–264. [Google Scholar] [CrossRef]
- Ueland, T.; Otterdal, K.; Lekva, T.; Halvorsen, B.; Gabrielsen, A.; Sandberg, W.J.; Paulsson-Berne, G.; Pedersen, T.M.; Folkersen, L.; Gullestad, L.; et al. Dickkopf-1 enhances inflammatory interaction between platelets and endothelial cells and shows increased expression in atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 1228–1234. [Google Scholar] [CrossRef] [PubMed]
- Babuin, L.; Jaffe, A.S. Troponin: The biomarker of choice for the detection of cardiac injury. Can. Med. Assoc. J. 2005, 173, 1191–1202. [Google Scholar] [CrossRef] [PubMed]
- Gaggin, H.K.; Januzzi, J.L., Jr. Natriuretic peptides in heart failure and acute coronary syndrome. Clin. Lab. Med. 2014, 34, 43–58. [Google Scholar] [CrossRef]
- Kohchi, C.; Noguchi, K.; Tanabe, Y.; Mizuno, D.; Soma, G. Constitutive expression of TNF-alpha and -beta genes in mouse embryo: Roles of cytokines as regulator and effector on development. Int. J. Biochem. 1994, 26, 111–119. [Google Scholar] [PubMed]
- Dick, S.A.; Epelman, S. Chronic Heart Failure and Inflammation: What Do We Really Know? Circ. Res. 2016, 119, 159–176. [Google Scholar] [CrossRef] [PubMed]
- Iwamatsu, T. Stages of normal development in the medaka Oryzias latipes. Mech. Dev. 2004, 121, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Fujita, M.; Isogai, S.; Kudo, A. Vascular anatomy of the developing medaka, Oryzias latipes: A complementary fish model for cardiovascular research on vertebrates. Dev. Dyn. 2006, 235, 734–746. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Doncel, M.; Okihiro, M.S.; Torija, C.F.; Tarazona, J.V.; Hinton, D.E. An artificial fertilization method with the Japanese medaka: Implications in early life stage bioassays and solvent toxicity. Ecotoxicol. Environ. Saf. 2008, 69, 95–103. [Google Scholar] [CrossRef]
- Kais, B.; Schneider, K.E.; Keiter, S.; Henn, K.; Ackermann, C.; Braunbeck, T. DMSO modifies the permeability of the zebrafish (Danio rerio) chorion-implications for the fish embryo test (FET). Aquat. Toxicol. 2013, 140–141, 229–238. [Google Scholar] [CrossRef]
- McCurley, A.T.; Callard, G.V. Characterization of housekeeping genes in zebrafish: Male-female differences and effects of tissue type, developmental stage and chemical treatment. BMC Mol. Biol. 2008, 9, 102. [Google Scholar] [CrossRef]
- Li, M.Y.; Tang, Y.H.; Liu, X.; Lu, H.Y.; Shi, X.Y. Sensitive determination of 4-O-methylhonokiol in rabbit plasma by high performance liquid chromatography and application to its pharmacokinetic investigation. J. Pharm. Anal. 2011, 1, 108–112. [Google Scholar] [CrossRef]
- Poivre, M.; Duez, P. Biological activity and toxicity of the Chinese herb Magnolia officinalis Rehder & E. Wilson (Houpo) and its constituents. J. Zhejiang Univ. Sci. B 2017, 18, 194–214. [Google Scholar] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sense (5′-3′) | Antisense (3′-5′) | Product Size (bp) | Reference |
|---|---|---|---|---|
| eEf1α | GGAGGCCAGCGACAAGAT | GCGAGAAGGTGGCAGGAT | 115 | NM_001104662.1 |
| FX | TGTCAAAGCCCTGTGTGAAT | AGAAATGTTCACAGCCACCA | 147 | XM_020710825.1 |
| FXI | GAAGGATAATGCAGACCAGTGTC | GATGACACCCTTCAAGTAGCATC | 127 | XM_004074394.4 |
| FVII | GTTCTGTCGGATAGGTGGATTT | CCTCCAGGTCATGTTTACCTAC | 97 | XM_004066449.3 |
| PAI-1 | ATGCCGAGGTTTTCTCTGAAC | GTTGAACATGTCTCCCAGTCC | 78 | XM_020711407.1 |
| uPA | ACTGTGTTTCTGGGAAAGAGTG | GGATGATCATTTTCTCCACGGT | 82 | XM_004077409.4 |
| tPA | CAGCCCCGATCCAAGC | CCCTTCCATCGCAGCC | 185 | XM_011478844.3 |
| ATRAP | CATGTGGGGGAACTTCAGC | GCCCACCAGAAACATGAGG | 91 | XM_011475678.2 |
| ETB | CTGATCTTTGTGGTGGGCAT | CCCATTCCTCATGCACTTGT | 78 | NM_001104844.1 |
| IL-1β | CTGTTTCTGGAGGAGGTGG | AGAAGAGGAAGCGCACATT | 79 | XM_011478737.2 |
| TNF-α | AACCGAAGAGTCTGAGAGGG | AGCTGAAGAAGAGTACCGCT | 105 | XM_004074335.3 |
| Catalase | TGCTAGCAGTTGATTGTCTGT | CACAGATCCACTGAAACAGGA | 100 | XM_004069460.2 |
| GPX2 | TCAACGGAGTAAACACGCAT | GATCCTGCATGAGAGAGCTG | 90 | XM_004082594.3 |
| GST A | CTGAAGGAGAGCGGCAC | CAGGAACGAGCCAGAGC | 107 | XM_020710769.1 |
| SOD2 | AAATGTGCGTCCTGACTATGT | TTTTGGCTATCTGAAGACGCT | 83 | XM_004083471.3 |
| Wnt1 | CCAGAAAACCCAGCTCACAA | TTGTGGGAGCAGAAGTTTGG | 80 | XM_020704658.1 |
| FZd2 | CACATGACCCCAGACTTCAC | AGAAACCAGAAGTGATGCCG | 76 | XM_020705151.1 |
| LRP5 | GAAGGCCCGAGCAGTTCA | AAGACATGGCTCCGTCGT | 101 | XM_011472833.2 |
| Dvl | TGCTGAAACAAAGCCCAAAGT | ACCTCAAGGATCTGAGTGAGC | 87 | XM_011490628.3 |
| β-catenin | CACAGAACTCCTACACAGCC | AGGCGCTTCTTGTAGTCTTG | 102 | XM_004077778.3 |
| DKK1 | GTGACACATGCCTGAGATCG | CACAGGCTTACAGATGCGAG | 83 | XM_020709512.1 |
| GSK3β | AGCTGCAGATTATGAGGAAGTTG | TAGACGGTCTCTGGAACATAGTC | 130 | XM_023950884.1 |
| ErbB3 | GAGGTTGAGAAGGATGGCGT | CTACCTGGACTTCCTGTGCC | 86 | XM_011474665.3 |
| NRG-2 | CTCGTCACTGTGGGGGA | CTCGTCAGTGGGGTCCA | 93 | XM_020708867.2 |
| BNP A | GAGCTCTGTTGATGAGGAGG | CAGTCCTGGCTCATCTTCTC | 88 | NM_001104685.2 |
| Troponin T | GAGCTCTGTTGATGAGGAGG | GCTGATCCGGTTTCTGAGT | 362 | XM_004068713.4 |
| FoxO1 | GCCCATGCCAGTTCTGAGTA | ATCCTCCGTGTTGGTGGATG | 102 | XM_011485361.2 |
| TGF-β2 | GTTACTCCGACCTGAGGAAGATAG | TGACACCCAATCTTTAACGGTTTC | 127 | XM_004073149.3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singha, S.K.; Muhammad, I.; Ibrahim, M.A.; Wang, M.; Ashpole, N.M.; Shariat-Madar, Z. 4-O-Methylhonokiol Influences Normal Cardiovascular Development in Medaka Embryo. Molecules 2019, 24, 475. https://doi.org/10.3390/molecules24030475
Singha SK, Muhammad I, Ibrahim MA, Wang M, Ashpole NM, Shariat-Madar Z. 4-O-Methylhonokiol Influences Normal Cardiovascular Development in Medaka Embryo. Molecules. 2019; 24(3):475. https://doi.org/10.3390/molecules24030475
Chicago/Turabian StyleSingha, Santu K., Ilias Muhammad, Mohamed Ali Ibrahim, Mei Wang, Nicole M. Ashpole, and Zia Shariat-Madar. 2019. "4-O-Methylhonokiol Influences Normal Cardiovascular Development in Medaka Embryo" Molecules 24, no. 3: 475. https://doi.org/10.3390/molecules24030475
APA StyleSingha, S. K., Muhammad, I., Ibrahim, M. A., Wang, M., Ashpole, N. M., & Shariat-Madar, Z. (2019). 4-O-Methylhonokiol Influences Normal Cardiovascular Development in Medaka Embryo. Molecules, 24(3), 475. https://doi.org/10.3390/molecules24030475