The Role of Phosphatidylethanolamine Adducts in Modification of the Activity of Membrane Proteins under Oxidative Stress



{kind=link}
{kind=link}
{kind=link}
Abstract
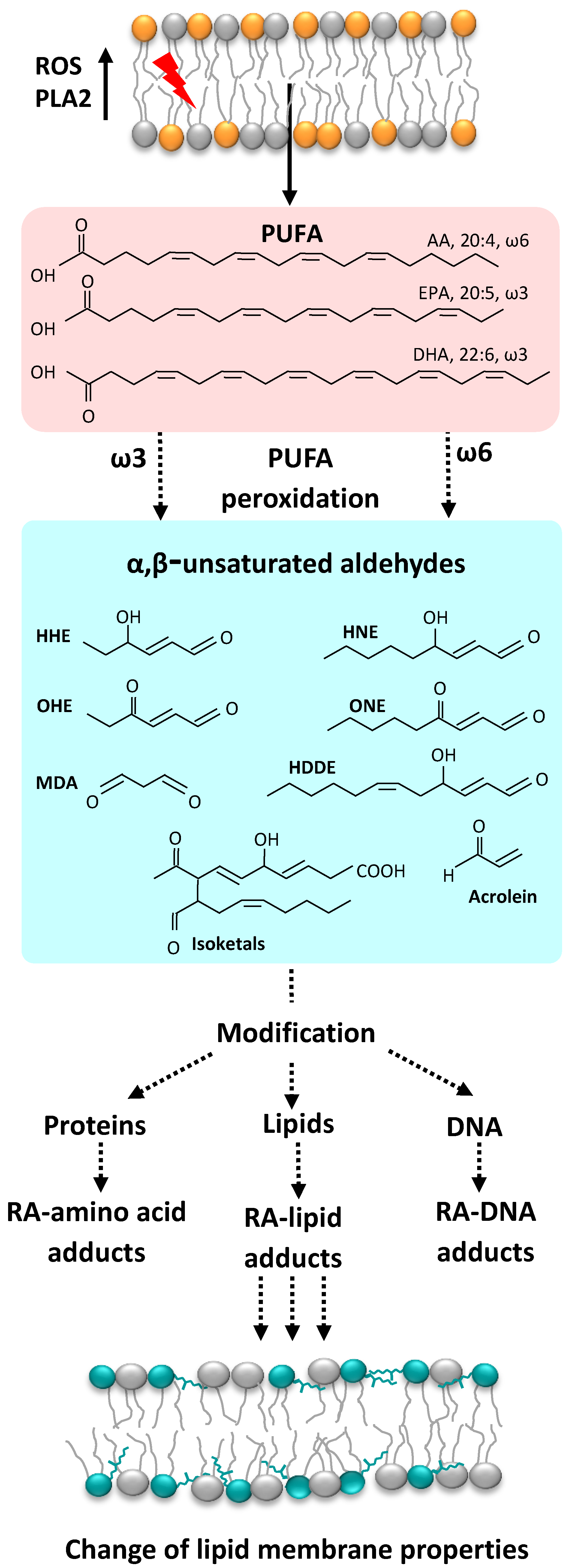
1. Reactive Oxygen Species and Their Derivatives, Reactive Aldehydes
2. ROS Detoxification Systems
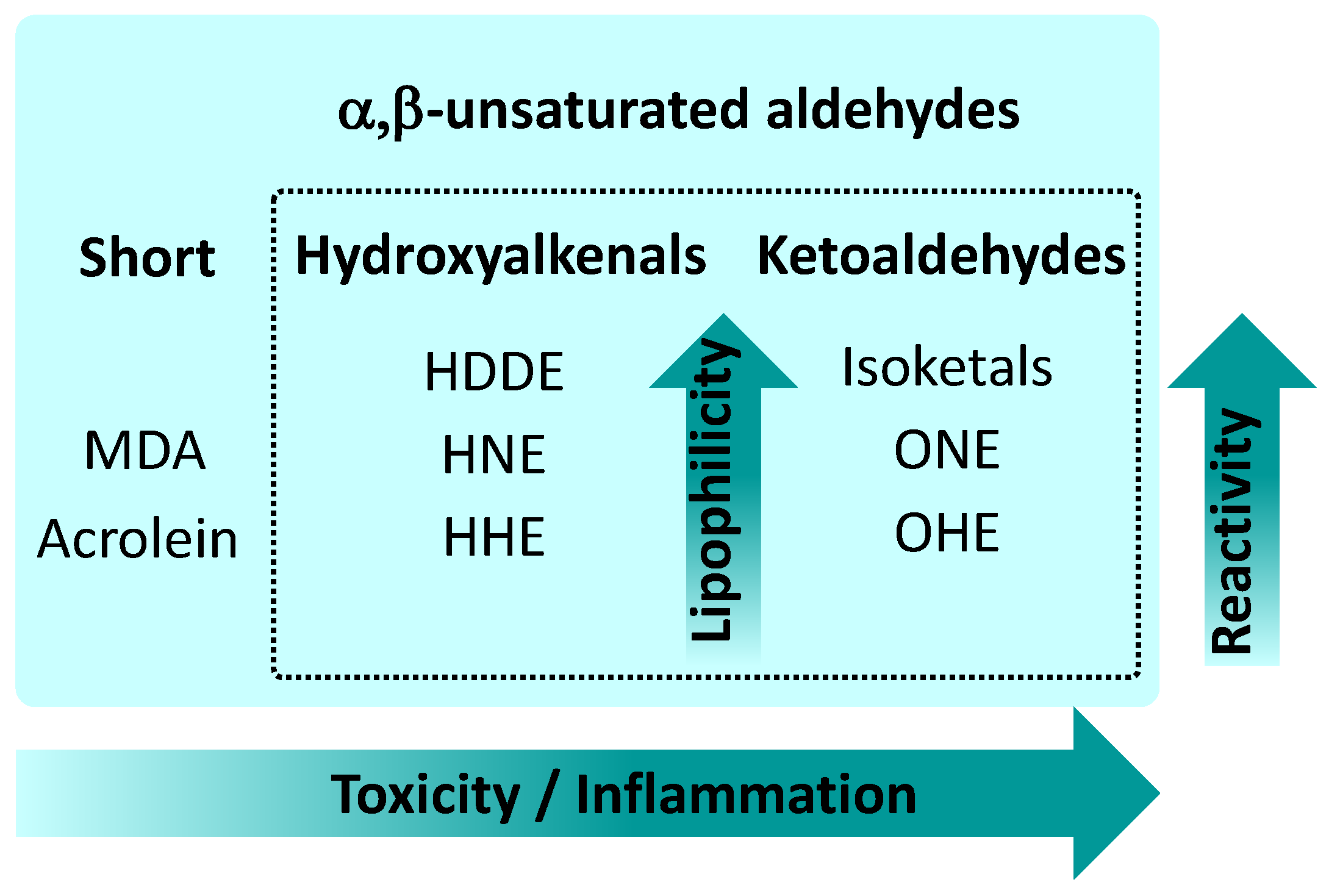
3. Mechanisms of RA Action
4. Phosphatidylethanolamine as a Crucial Target for Reactive Aldehydes
4.1. Phosphatidylethanolamine and Its Physiological Functions
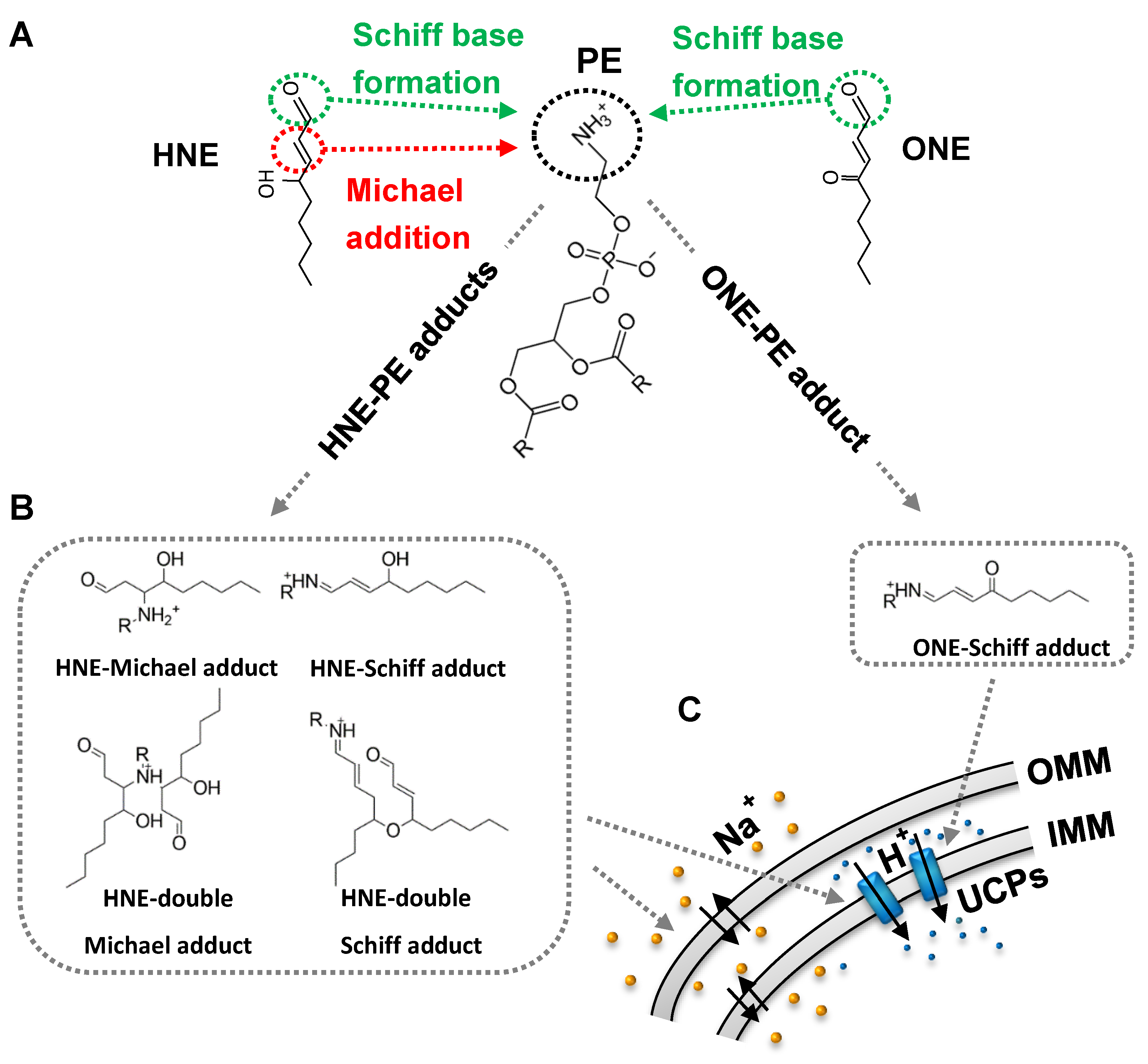
4.2. PE Adducts
4.3. Modification of Membrane Properties by PE and PE Adducts
4.4. Modification of Membrane Transporter Function in the Presence of PE Adducts
5. Conclusions and Outlook
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Skulachev, V.P. Mitochondria-Targeted Antioxidants as Promising Drugs for Treatment of Age-Related Brain Diseases. J. Alzheimers Dis. 2012, 28, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Wong, H.S.; Dighe, P.A.; Mezera, V.; Monternier, P.A.; Brand, M.D. Production of Superoxide and Hydrogen Peroxide from Specific Mitochondrial Sites under Different Bioenergetic Conditions. J. Biol. Chem. 2017, 292, 16804–16809. [Google Scholar] [CrossRef] [PubMed]
- Brand, M.D. Mitochondrial Generation of Superoxide and Hydrogen Peroxide as the Source of Mitochondrial Redox Signaling. Free Radic. Biol. Med. 2016, 100, 14–31. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M. Mitochondrial Superoxide Production in Health and Disease. Biochim. Biophys. Acta 2016, 1857, E6–E7. [Google Scholar] [CrossRef]
- Vinogradov, A.D.; Grivennikova, V.G. Oxidation of Nadh And Ros Production By Respiratory Complex, I. Biochim. Biophys. Acta 2016, 1857, 863–871. [Google Scholar] [CrossRef]
- Naudi, A.; Jove, M.; Ayala, V.; Cabre, R.; Portero-Otin, M.; Pamplona, R. Non-Enzymatic Modification of Aminophospholipids by Carbonyl-Amine Reactions. Int. J. Mol. Sci. 2013, 14, 3285–3313. [Google Scholar] [CrossRef]
- Esterbauer, H.; Schaur, R.J.; Zollner, H. Chemistry and Biochemistry Of 4-Hydroxynonenal, Malonaldehyde and Related Aldehydes. Free Radic. Biol. Med. 1991, 11, 81–128. [Google Scholar] [CrossRef]
- Roede, J.R.; Jones, D.P. Reactive Species and Mitochondrial Dysfunction: Mechanistic Significance of 4-Hydroxynonenal. Environ Mol Mutagen 2010, 51, 380–390. [Google Scholar] [CrossRef]
- Zarkovic, N. 4-Hydroxynonenal as a Bioactive Marker of Pathophysiological Processes. Mol. Asp. Med. 2003, 24, 281–291. [Google Scholar] [CrossRef]
- Kasai, H.; Maekawa, M.; Kawai, K.; Hachisuka, K.; Takahashi, Y.; Nakamura, H.; Sawa, R.; Matsui, S.; Matsuda, T. 4-Oxo-2-Hexenal, A Mutagen Formed By Omega-3 Fat Peroxidation, Causes Dna Adduct Formation In Mouse Organs. Ind Health 2005, 43, 699–701. [Google Scholar] [CrossRef][Green Version]
- Lin, D.; Lee, H.G.; Liu, Q.; Perry, G.; Smith, M.A.; Sayre, L.M. 4-Oxo-2-Nonenal Is Both More Neurotoxic And More Protein Reactive Than 4-Hydroxy-2-Nonenal. Chem. Res. Toxicol. 2005, 18, 1219–1231. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free Radicals And Antioxidants In Normal Physiological Functions And Human Disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Benedetti, A.; Comporti, M.; Fulceri, R.; Esterbauer, H. Cytotoxic Aldehydes Originating from the Peroxidation OF Liver Microsomal Lipids. Identification Of 4,5-Dihydroxydecenal. Biochim. Biophys. Acta 1984, 792, 172–181. [Google Scholar] [CrossRef]
- Uchida, K. 4-Hydroxy-2-Nonenal: A Product and Mediator of Oxidative Stress. Prog. Lipid Res. 2003, 42, 318–343. [Google Scholar] [CrossRef]
- Koster, J.F.; Slee, R.G.; Montfoort, A.; Lang, J.; Esterbauer, H. Comparison of The Inactivation of Microsomal Glucose-6-Phosphatase By In Situ Lipid Peroxidation-Derived 4-Hydroxynonenal And Exogenous 4-Hydroxynonenal. Free Radic. Res. Commun. 1986, 1, 273–287. [Google Scholar] [CrossRef]
- Jovanovic, O.; Pashkovskaya, A.A.; Annibal, A.; Vazdar, M.; Burchardt, N.; Sansone, A.; Gille, L.; Fedorova, M.; Ferreri, C.; Pohl, E.E. The Molecular Mechanism behind Reactive Aldehyde Action on Transmembrane Translocations of Proton and Potassium Ions. Free Radic. Biol. Med. 2015, 89, 1067–1076. [Google Scholar] [CrossRef]
- Droge, W. Free Radicals in the Physiological Control of Cell Function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef]
- Hannesschlaeger, C.; Pohl, P. Membrane Permeabilities of Ascorbic Acid and Ascorbate. Biomolecules 2018, 8, 73. [Google Scholar] [CrossRef]
- Lushchak, V.I. Free Radicals, Reactive Oxygen Species, Oxidative Stress and Its Classification. Chem. Biol. Interact. 2014, 224, 164–175. [Google Scholar] [CrossRef]
- Tikhonova, I.M.; Andreyev, A.Y.; Kaulen, A.D.; Komrakov, A.Y.; Skulachev, V.P. Ion Permeability Induced In Artificial Membranes By The Atp/Adp Antiporter. FEBS Lett. 1994, 337, 231–234. [Google Scholar] [CrossRef]
- Bertholet, A.M.; Chouchani, E.T.; Kazak, L.; Angelin, A.; Fedorenko, A.; Long, J.Z.; Vidoni, S.; Garrity, R.; Cho, J.; Terada, N.; et al. H(+) Transport Is An Integral Function of The Mitochondrial Adp/Atp Carrier. Nature 2019, 571, 515–520. [Google Scholar] [CrossRef]
- Wieckowski, M.R.; Wojtczak, L. Involvement of the Dicarboxylate Carrier in the Protonophoric Action of Long-Chain Fatty Acids In Mitochondria. Biochem. Biophys. Res. Commun. 1997, 232, 414–417. [Google Scholar]
- Samartsev, V.N.; Marchik, E.I.; Shamagulova, L.V. Free Fatty Acids As Inducers And Regulators Of Uncoupling Of Oxidative Phosphorylation In Liver Mitochondria With Participation Of Adp/Atp- And Aspartate/Glutamate-Antiporter. Biochemistry 2011, 76, 217–224. [Google Scholar] [CrossRef]
- Shabalina, I.G.; Jacobsson, A.; Cannon, B.; Nedergaard, J. Native Ucp1 Displays Simple Competitive Kinetics between the Regulators Purine Nucleotides and Fatty Acids. J. Biol. Chem. 2004, 279, 38236–38248. [Google Scholar] [CrossRef]
- Urbankova, E.; Voltchenko, A.; Pohl, P.; Jezek, P.; Pohl, E.E. Transport Kinetics Of Uncoupling Proteins. Analysis of Ucp1 Reconstituted In Planar Lipid Bilayers. J. Biol. Chem. 2003, 278, 32497–32500. [Google Scholar] [CrossRef]
- Macher, G.; Koehler, M.; Rupprecht, A.; Kreiter, J.; Hinterdorfer, P.; Pohl, E.E. Inhibition Of Mitochondrial Ucp1 And Ucp3 By Purine Nucleotides And Phosphate. Biochim. Biophys. Acta Biomembr. 2018, 1860, 664–672. [Google Scholar] [CrossRef]
- Jezek, P.; Orosz, D.E.; Garlid, K.D. Reconstitution of the Uncoupling Protein of Brown Adipose Tissue Mitochondria. Demonstration of Gdp-Sensitive Halide Anion Uniport. J. Biol. Chem. 1990, 265, 19296–19302. [Google Scholar]
- Bouillaud, F.; Ricquier, D.; Gulik-Krzywicki, T.; Gary-Bobo, C.M. The Possible Proton Translocating Activity of the Mitochondrial Uncoupling Protein of Brown Adipose Tissue. Reconstitution Studies in Liposomes. FEBS Lett. 1983, 164, 272–276. [Google Scholar] [CrossRef][Green Version]
- Bienengraeber, M.; Echtay, K.S.; Klingenberg, M. H+ Transport by Uncoupling Protein (Ucp-1) Is Dependent On A Histidine Pair, Absent In Ucp-2 and Ucp-3. Biochemistry 1998, 37, 3–8. [Google Scholar] [CrossRef]
- Cannon, B.; Shabalina, I.G.; Kramarova, T.V.; Petrovic, N.; Nedergaard, J. Uncoupling Proteins: A Role In Protection Against Reactive Oxygen Species-Or Not? Biochim. Biophys. Acta 2006, 1757, 449–458. [Google Scholar] [CrossRef]
- Pohl, E.E.; Rupprecht, A.; Macher, G.; Hilse, K.E. Important Trends In Ucp3 Investigation. Front. Physiol. 2019, 10, 470. [Google Scholar] [CrossRef]
- Vozza, A.; Parisi, G.; De Leonardis, F.; Lasorsa, F.M.; Castegna, A.; Amorese, D.; Marmo, R.; Calcagnile, V.M.; Palmieri, L.; Ricquier, D.; et al. Ucp2 Transports C4 Metabolites Out Of Mitochondria, Regulating Glucose And Glutamine Oxidation. Proc. Natl. Acad. Sci. USA 2014, 111, 960–965. [Google Scholar] [CrossRef]
- Rupprecht, A.; Moldzio, R.; Modl, B.; Pohl, E.E. Glutamine Regulates Mitochondrial Uncoupling Protein 2 To Promote Glutaminolysis In Neuroblastoma Cells. Biochim. Biophys. Acta Bioenerg. 2019, 1860, 391–401. [Google Scholar] [CrossRef]
- Hilse, K.E.; Rupprecht, A.; Egerbacher, M.; Bardakji, S.; Zimmermann, L.; Wulczyn, A.; Pohl, E.E. The Expression Of Uncoupling Protein 3 Coincides With The Fatty Acid Oxidation Type Of Metabolism In Adult Murine Heart. Front. Physiol. 2018, 9, 747. [Google Scholar] [CrossRef]
- Beck, V.; Jaburek, M.; Demina, T.; Rupprecht, A.; Porter, R.K.; Jezek, P.; Pohl, E.E. Polyunsaturated Fatty Acids Activate Human Uncoupling Proteins 1 And 2 In Planar Lipid Bilayers. FASEB J. 2007, 21, 1137–1144. [Google Scholar] [CrossRef]
- Zackova, M.; Jezek, P. Reconstitution of Novel Mitochondrial Uncoupling Proteins Ucp2 and Ucp3. Biosci. Rep. 2002, 22, 33–46. [Google Scholar] [CrossRef]
- Zackova, M.; Skobisova, E.; Urbankova, E.; Jezek, P. Activating Omega-6 Polyunsaturated Fatty Acids And Inhibitory Purine Nucleotides Are High Affinity Ligands For Novel Mitochondrial Uncoupling Proteins Ucp2 And Ucp3. J. Biol. Chem. 2003, 278, 20761–20769. [Google Scholar] [CrossRef]
- Rupprecht, A.; Sittner, D.; Smorodchenko, A.; Hilse, K.E.; Goyn, J.; Moldzio, R.; Seiler, A.E.; Brauer, A.U.; Pohl, E.E. Uncoupling Protein 2 And 4 Expression Pattern During Stem Cell Differentiation Provides New Insight Into Their Putative Function. PLoS ONE 2014, 9, E88474. [Google Scholar] [CrossRef]
- Hilse, K.E.; Kalinovich, A.V.; Rupprecht, A.; Smorodchenko, A.; Zeitz, U.; Staniek, K.; Erben, R.G.; Pohl, E.E. The Expression Of Ucp3 Directly Correlates To Ucp1 Abundance In Brown Adipose Tissue. Biochim. Biophys. Acta 2016, 1857, 72–78. [Google Scholar] [CrossRef]
- Negre-Salvayre, A.; Hirtz, C.; Carrera, G.; Cazenave, R.; Troly, M.; Salvayre, R.; Penicaud, L.; Casteilla, L. A Role for Uncoupling Protein-2 as a Regulator of Mitochondrial Hydrogen Peroxide Generation. Faseb, J. 1997, 11, 809–815. [Google Scholar] [CrossRef]
- Echtay, K.S.; Roussel, D.; St-Pierre, J.; Jekabsons, M.B.; Cadenas, S.; Stuart, J.A.; Harper, J.A.; Roebuck, S.J.; Morrison, A.; Pickering, S.; et al. Superoxide Activates Mitochondrial Uncoupling Proteins. Nature 2002, 415, 96–99. [Google Scholar] [CrossRef]
- Echtay, K.S.; Esteves, T.C.; Pakay, J.L.; Jekabsons, M.B.; Lambert, A.J.; Portero-Otin, M.; Pamplona, R.; Vidal-Puig, A.J.; Wang, S.; Roebuck, S.J.; et al. A Signalling Role for 4-Hydroxy-2-Nonenal in Regulation of Mitochondrial Uncoupling. EMBO J. 2003, 22, 4103–4110. [Google Scholar] [CrossRef]
- Krauss, S.; Zhang, C.Y.; Lowell, B.B. The Mitochondrial Uncoupling-Protein Homologues. Nat. Rev. Mol. Cell Biol. 2005, 6, 248–261. [Google Scholar] [CrossRef]
- Shabalina, I.G.; Petrovic, N.; Kramarova, T.V.; Hoeks, J.; Cannon, B.; Nedergaard, J. Ucp1 And Defense Against Oxidative Stress. 4-Hydroxy-2-Nonenal Effects On Brown Fat Mitochondria Are Uncoupling Protein 1-Independent. J. Biol. Chem. 2006, 281, 13882–13893. [Google Scholar] [CrossRef]
- Couplan, E.; Mar Gonzalez-Barroso, M.; Alves-Guerra, M.C.; Ricquier, D.; Goubern, M.; Bouillaud, F. No Evidence for A Basal, Retinoic, or Superoxide-Induced Uncoupling Activity of the Uncoupling Protein 2 Present In Spleen or Lung Mitochondria. J. Biol.Chem. 2002, 277, 26268–26275. [Google Scholar] [CrossRef]
- Lombardi, A.; Grasso, P.; Moreno, M.; De Lange, P.; Silvestri, E.; Lanni, A.; Goglia, F. Interrelated Influence Of Superoxides And Free Fatty Acids Over Mitochondrial Uncoupling In Skeletal Muscle. Biochim. Biophys. Acta 2008, 1777, 826–833. [Google Scholar] [CrossRef][Green Version]
- Parker, N.; Vidal-Puig, A.; Brand, M.D. Stimulation of Mitochondrial Proton Conductance by Hydroxynonenal Requires A High Membrane Potential. Biosci. Rep. 2008, 28, 83–88. [Google Scholar] [CrossRef]
- Malingriaux, E.A.; Rupprecht, A.; Gille, L.; Jovanovic, O.; Jezek, P.; Jaburek, M.; Pohl, E.E. Fatty Acids Are Key In 4-Hydroxy-2-Nonenal-Mediated Activation Of Uncoupling Proteins 1 And 2. PLoS ONE 2013, 8, E77786. [Google Scholar] [CrossRef]
- Zarkovic, N.; Cipak, A.; Jaganjac, M.; Borovic, S.; Zarkovic, K. Pathophysiological Relevance of Aldehydic Protein Modifications. J. Proteomics. 2013, 92, 239–247. [Google Scholar] [CrossRef]
- Fritz, K.S.; Petersen, D.R. An Overview of the Chemistry and Biology Of Reactive Aldehydes. Free Radic. Biol. Med. 2013, 59, 85–91. [Google Scholar] [CrossRef]
- Castro, J.P.; Jung, T.; Grune, T.; Siems, W. 4-Hydroxynonenal (Hne) Modified Proteins in Metabolic Diseases. Free Radic. Biol. Med. 2017, 111, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Voulgaridou, G.P.; Anestopoulos, I.; Franco, R.; Panayiotidis, M.I.; Pappa, A. Dna Damage Induced By Endogenous Aldehydes: Current State Of Knowledge. Mutat. Res. 2011, 711, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Gentile, F.; Arcaro, A.; Pizzimenti, S.; Daga, M.; Cetrangolo, G.P.; Dianzani, C.; Lepore, A.; Graf, M.; Ames, P.R.J.; Barrera, G. Dna Damage By Lipid Peroxidation Products: Implications In Cancer, Inflammation And Autoimmunity. AIMS Genet. 2017, 4, 103–137. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Davies, S.S. Bioactive Aldehyde-Modified Phosphatidylethanolamines. Biochimie 2013, 95, 74–78. [Google Scholar] [CrossRef]
- Sousa, B.C.; Pitt, A.R.; Spickett, C.M. Chemistry and Analysis of Hne and Other Prominent Carbonyl-Containing Lipid Oxidation Compounds. Free Radic. Biol. Med. 2017, 111, 294–308. [Google Scholar] [CrossRef] [PubMed]
- Sokolov, V.S.; Batishchev, O.V.; Akimov, S.A.; Galimzyanov, T.R.; Konstantinova, A.N.; Malingriaux, E.; Gorbunova, Y.G.; Knyazev, D.G.; Pohl, P. Residence Time Of Singlet Oxygen In Membranes. Sci. Rep. 2018, 8, 14000. [Google Scholar] [CrossRef]
- Gueraud, F.; Atalay, M.; Bresgen, N.; Cipak, A.; Eckl, P.M.; Huc, L.; Jouanin, I.; Siems, W.; Uchida, K. Chemistry And Biochemistry Of Lipid Peroxidation Products. Free Radic. Res. 2010, 44, 1098–1124. [Google Scholar] [CrossRef]
- Doorn, J.A.; Petersen, D.R. Covalent Modification of Amino Acid Nucleophiles by the Lipid Peroxidation Products 4-Hydroxy-2-Nonenal and 4-Oxo-2-Nonenal. Chem. Res. Toxicol. 2002, 15, 1445–1450. [Google Scholar] [CrossRef]
- Carbone, D.L.; Doorn, J.A.; Kiebler, Z.; Petersen, D.R. Cysteine Modification by Lipid Peroxidation Products Inhibits Protein Disulfide Isomerase. Chem. Res. Toxicol. 2005, 18, 1324–1331. [Google Scholar] [CrossRef]
- Brame, C.J.; Salomon, R.G.; Morrow, J.D.; Roberts, L.J., 2nd. Identification Of Extremely Reactive Gamma-Ketoaldehydes (Isolevuglandins) As Products Of The Isoprostane Pathway And Characterization of Their Lysyl Protein Adducts. J. Biol. Chem. 1999, 274, 13139–13146. [Google Scholar] [CrossRef]
- Bleier, L.; Wittig, I.; Heide, H.; Steger, M.; Brandt, U.; Drose, S. Generator-Specific Targets Of Mitochondrial Reactive Oxygen Species. Free Radic. Biol. Med. 2015, 78, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bogeski, I.; Kappl, R.; Kummerow, C.; Gulaboski, R.; Hoth, M.; Niemeyer, B.A. Redox Regulation of Calcium Ion Channels: Chemical And Physiological Aspects. Cell Calcium 2011, 50, 407–423. [Google Scholar] [CrossRef] [PubMed]
- Van Der Veen, J.N.; Kennelly, J.P.; Wan, S.; Vance, J.E.; Vance, D.E.; Jacobs, R.L. The Critical Role of Phosphatidylcholine and Phosphatidylethanolamine Metabolism in Health and Disease. Biochim. Biophys. Acta Biomembr. 2017, 1859, 1558–1572. [Google Scholar] [CrossRef] [PubMed]
- Vance, J.E.; Tasseva, G. Formation and Function of Phosphatidylserine and Phosphatidylethanolamine in Mammalian Cells. Biochim. Biophys. Acta 2013, 1831, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Devaux, P.F.; Morris, R. Transmembrane Asymmetry and Lateral Domains in Biological Membranes. Traffic 2004, 5, 241–246. [Google Scholar] [CrossRef]
- Epand, R.M.; Fuller, N.; Rand, R.P. Role of the Position of Unsaturation on The Phase Behavior And Intrinsic Curvature Of Phosphatidylethanolamines. Biophys. J. 1996, 71, 1806–1810. [Google Scholar] [CrossRef]
- Van Den Brink-Van Der Laan, E.; Killian, J.A.; De Kruijff, B. Nonbilayer Lipids Affect Peripheral and Integral Membrane Proteins via Changes in the Lateral Pressure Profile. Biochim. Biophys. Acta 2004, 1666, 275–288. [Google Scholar] [CrossRef]
- Cullis, P.R.; De Kruijff, B. Lipid Polymorphism and the Functional Roles of Lipids in Biological Membranes. Biochim. Biophys. Acta 1979, 559, 399–420. [Google Scholar] [CrossRef]
- Verkleij, A.J.; Leunissen-Bijvelt, J.; De Kruijff, B.; Hope, M.; Cullis, P.R. Non-Bilayer Structures in Membrane Fusion. Ciba Found. Symp. 1984, 103, 45–59. [Google Scholar]
- Siegel, D.P.; Epand, R.M. The Mechanism of Lamellar-To-Inverted Hexagonal Phase Transitions in Phosphatidylethanolamine: Implications for Membrane Fusion Mechanisms. Biophys. J. 1997, 73, 3089–3111. [Google Scholar] [CrossRef]
- Martens, C.; Shekhar, M.; Borysik, A.J.; Lau, A.M.; Reading, E.; Tajkhorshid, E.; Booth, P.J.; Politis, A. Direct Protein-Lipid Interactions Shape The Conformational Landscape Of Secondary Transporters. Nat. Commun. 2018, 9, 4151. [Google Scholar] [CrossRef] [PubMed]
- Van Den Brink-Van Der Laan, E.; Chupin, V.; Killian, J.A.; De Kruijff, B. Stability of Kcsa Tetramer Depends On Membrane Lateral Pressure. Biochemistry 2004, 43, 4240–4250. [Google Scholar] [CrossRef] [PubMed]
- Vance, J.E. Phospholipid Synthesis and Transport in Mammalian Cells. Traffic 2015, 16, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Bogdanov, M.; Dowhan, W. Lipid-Assisted Protein Folding. J. Biol. Chem. 1999, 274, 36827–36830. [Google Scholar] [CrossRef]
- Shinzawa-Itoh, K.; Aoyama, H.; Muramoto, K.; Terada, H.; Kurauchi, T.; Tadehara, Y.; Yamasaki, A.; Sugimura, T.; Kurono, S.; Tsujimoto, K.; et al. Structures And Physiological Roles Of 13 Integral Lipids Of Bovine Heart Cytochrome C Oxidase. EMBO J. 2007, 26, 1713–1725. [Google Scholar] [CrossRef]
- Calzada, E.; Avery, E.; Sam, P.N.; Modak, A.; Wang, C.; Mccaffery, J.M.; Han, X.; Alder, N.N.; Claypool, S.M. Phosphatidylethanolamine Made In The Inner Mitochondrial Membrane Is Essential For Yeast Cytochrome Bc1 Complex Function. Nat. Commun. 2019, 10, 1432. [Google Scholar] [CrossRef]
- Ichimura, Y.; Kirisako, T.; Takao, T.; Satomi, Y.; Shimonishi, Y.; Ishihara, N.; Mizushima, N.; Tanida, I.; Kominami, E.; Ohsumi, M.; et al. A Ubiquitin-Like System Mediates Protein Lipidation. Nature 2000, 408, 488–492. [Google Scholar] [CrossRef]
- Kagan, V.E.; Mao, G.; Qu, F.; Angeli, J.P.; Doll, S.; Croix, C.S.; Dar, H.H.; Liu, B.; Tyurin, V.A.; Ritov, V.B.; et al. Oxidized Arachidonic And Adrenic Pes Navigate Cells To Ferroptosis. Nat. Chem. Biol. 2017, 13, 81–90. [Google Scholar] [CrossRef]
- Deleault, N.R.; Piro, J.R.; Walsh, D.J.; Wang, F.; Ma, J.; Geoghegan, J.C.; Supattapone, S. Isolation Of Phosphatidylethanolamine As A Solitary Cofactor For Prion Formation In The Absence Of Nucleic Acids. Proc. Natl. Acad. Sci. USA 2012, 109, 8546–8551. [Google Scholar] [CrossRef]
- Menon, A.K.; Eppinger, M.; Mayor, S.; Schwarz, R.T. Phosphatidylethanolamine Is the Donor of the Terminal Phosphoethanolamine Group in Trypanosome Glycosylphosphatidylinositols. EMBO J. 1993, 12, 1907–1914. [Google Scholar] [CrossRef]
- Vance, J.E. Historical Perspective: Phosphatidylserine and Phosphatidylethanolamine from the 1800s to the Present. J. Lipid Res. 2018, 59, 923–944. [Google Scholar] [CrossRef] [PubMed]
- Calzada, E.; Onguka, O.; Claypool, S.M. Phosphatidylethanolamine Metabolism in Health And Disease. Int. Rev. Cell Mol. Biol. 2016, 321, 29–88. [Google Scholar] [PubMed]
- Chidley, C.; Trauger, S.A.; Birsoy, K.; O’shea, E.K. The Anticancer Natural Product Ophiobolin A Induces Cytotoxicity By Covalent Modification of Phosphatidylethanolamine. Elife 2016, 5, E14601. [Google Scholar] [CrossRef] [PubMed]
- Guichardant, M.; Taibi-Tronche, P.; Fay, L.B.; Lagarde, M. Covalent Modifications of Aminophospholipids by 4-Hydroxynonenal. Free Radic. Biol. Med. 1998, 25, 1049–1056. [Google Scholar] [CrossRef]
- Bacot, S.; Bernoud-Hubac, N.; Baddas, N.; Chantegrel, B.; Deshayes, C.; Doutheau, A.; Lagarde, M.; Guichardant, M. Covalent Binding Of Hydroxy-Alkenals 4-Hdde, 4-Hhe, And 4-Hne To Ethanolamine Phospholipid Subclasses. J. Lipid Res. 2003, 44, 917–926. [Google Scholar] [CrossRef]
- Guo, L.; Amarnath, V.; Davies, S.S. A Liquid Chromatography-Tandem Mass Spectrometry Method for Measurement of N-Modified Phosphatidylethanolamines. Anal. Biochem. 2010, 405, 236–245. [Google Scholar] [CrossRef][Green Version]
- Annibal, A.; Schubert, K.; Wagner, U.; Hoffmann, R.; Schiller, J.; Fedorova, M. New Covalent Modifications Of Phosphatidylethanolamine By Alkanals: Mass Spectrometry Based Structural Characterization And Biological Effects. J. Mass Spectrom. 2014, 49, 557–569. [Google Scholar] [CrossRef]
- Vazdar, K.; Vojta, D.; Margetic, D.; Vazdar, M. Reaction Mechanism of Covalent Modification of Phosphatidylethanolamine Lipids by Reactive Aldehydes 4-Hydroxy-2-Nonenal And 4-Oxo-2-Nonenal. Chem. Res. Toxicol. 2017, 30, 840–850. [Google Scholar] [CrossRef]
- Jovanovic, O.; Skulj, S.; Pohl, E.E.; Vazdar, M. Covalent Modification of Phosphatidylethanolamine by 4-Hydroxy-2-Nonenal Increases Sodium Permeability across Phospholipid Bilayer Membranes. Free Radic. Biol. Med. 2019, 143, 433–440. [Google Scholar] [CrossRef]
- Annibal, A.; Riemer, T.; Jovanovic, O.; Westphal, D.; Griesser, E.; Pohl, E.E.; Schiller, J.; Hoffmann, R.; Fedorova, M. Structural, Biological And Biophysical Properties of Glycated and Glycoxidized Phosphatidylethanolamines. Free Radic. Biol. Med. 2016, 95, 293–307. [Google Scholar] [CrossRef]
- Guo, L.; Chen, Z.; Amarnath, V.; Davies, S.S. Identification Of Novel Bioactive Aldehyde-Modified Phosphatidylethanolamines Formed By Lipid Peroxidation. Free Radic. Biol.Med. 2012, 53, 1226–1238. [Google Scholar] [CrossRef] [PubMed]
- Stadelmann-Ingrand, S.; Favreliere, S.; Fauconneau, B.; Mauco, G.; Tallineau, C. Plasmalogen Degradation by Oxidative Stress: Production and Disappearance of Specific Fatty Aldehydes and Fatty Alpha-Hydroxyaldehydes. Free Radic. Biol. Med. 2001, 31, 1263–1271. [Google Scholar] [CrossRef]
- Stadelmann-Ingrand, S.; Pontcharraud, R.; Fauconneau, B. Evidence For The Reactivity Of Fatty Aldehydes Released From Oxidized Plasmalogens With Phosphatidylethanolamine To Form Schiff Base Adducts In Rat Brain Homogenates. Chem. Phys. Lipids 2004, 131, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Chen, Z.; Cox, B.E.; Amarnath, V.; Epand, R.F.; Epand, R.M.; Davies, S.S. Phosphatidylethanolamines Modified By Gamma-Ketoaldehyde (Gammaka) Induce Endoplasmic Reticulum Stress And Endothelial Activation. J. Biol. Chem. 2011, 286, 18170–18180. [Google Scholar] [CrossRef]
- Bernoud-Hubac, N.; Fay, L.B.; Armarnath, V.; Guichardant, M.; Bacot, S.; Davies, S.S.; Roberts II, L.J.; Lagarde, M. Covalent Binding Of Isoketals to Ethanolamine Phospholipids. Free Radic. Biol. Med. 2004, 37, 1604–1611. [Google Scholar] [CrossRef]
- Sullivan, C.B.; Matafonova, E.; Roberts II, L.J.; Amarnath, V.; Davies, S.S. Isoketals Form Cytotoxic Phosphatidylethanolamine Adducts In Cells. J. Lipid Res. 2010, 51, 999–1009. [Google Scholar] [CrossRef]
- Guo, L.; Chen, Z.; Amarnath, V.; Yancey, P.G.; Van Lenten, B.J.; Savage, J.R.; Fazio, S.; Linton, M.F.; Davies, S.S. Isolevuglandin-Type Lipid Aldehydes Induce The Inflammatory Response Of Macrophages By Modifying Phosphatidylethanolamines And Activating The Receptor For Advanced Glycation Endproducts. Antioxid. Redox Signal. 2015, 22, 1633–1645. [Google Scholar] [CrossRef]
- Davies, S.D.; May-Zhang, L.S.; Boutaud, O.; Amarnath, V.; Kirabo, A.; Harrison, D.G. Isolevuglandins as Mediators of Disease and the Development of Dicarbonyl Scavengers as Pharmaceutical Interventions. Pharmacol. Ther 2019, 107418. [Google Scholar] [CrossRef]
- Salomon, R.G.; Bi, W. Isolevuglandin Adducts In Disease. Antioxid. Redox Signal. 2015, 22, 1703–1718. [Google Scholar] [CrossRef]
- May-Zhang, L.S.; Yermalitsky, V.; Huang, J.; Pleasent, T.; Borja, M.S.; Oda, M.N.; Jerome, W.G.; Yancey, P.G.; Linton, M.F.; Davies, S.S. Modification By Isolevuglandins, Highly Reactive Gamma-Ketoaldehydes, Deleteriously Alters High-Density Lipoprotein Structure And Function. J. Biol. Chem. 2018, 293, 9176–9187. [Google Scholar] [CrossRef]
- Zemski Berry, K.A.; Murphy, R.C. Characterization of Acrolein-Glycerophosphoethanolamine Lipid Adducts Using Electrospray Mass Spectrometry. Chem. Res. Toxicol. 2007, 20, 1342–1351. [Google Scholar] [CrossRef] [PubMed]
- Ravandi, A.; Kuksis, A.; Marai, L.; Myher, J.J. Preparation and Characterization of Glucosylated Aminoglycerophospholipids. Lipids 1995, 30, 885–891. [Google Scholar] [CrossRef] [PubMed]
- Fountain, W.C.; Requena, J.R.; Jenkins, A.J.; Lyons, T.J.; Smyth, B.; Baynes, J.W.; Thorpe, S.R. Quantification of N-(Glucitol)Ethanolamine and N-(Carboxymethyl)Serine: Two Products of Nonenzymatic Modification of Aminophospholipids Formed in Vivo. Anal. Biochem. 1999, 272, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Requena, J.R.; Ahmed, M.U.; Fountain, C.W.; Degenhardt, T.P.; Reddy, S.; Perez, C.; Lyons, T.J.; Jenkins, A.J.; Baynes, J.W.; Thorpe, S.R. Carboxymethylethanolamine, A Biomarker Of Phospholipid Modification During The Maillard Reaction In Vivo. J. Biol. Chem. 1997, 272, 17473–17479. [Google Scholar] [CrossRef]
- Lapolla, A.; Fedele, D.; Traldi, P. Glyco-Oxidation in Diabetes and Related Diseases. Clin. Chim. Acta 2005, 357, 236–250. [Google Scholar] [CrossRef]
- Simoes, C.; Silva, A.C.; Domingues, P.; Laranjeira, P.; Paiva, A.; Domingues, M.R. Modified Phosphatidylethanolamines Induce Different Levels Of Cytokine Expression In Monocytes And Dendritic Cells. Chem. Phys. Lipids 2013, 175, 57–64. [Google Scholar] [CrossRef]
- Van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane Lipids: Where they are and How They Behave. Nat. Rev. Mol. Cell Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef]
- Patel, D.; Witt, S.N. Ethanolamine and Phosphatidylethanolamine: Partners in Health and Disease. Oxid. Med. Cell Longev. 2017, 2017, 4829180. [Google Scholar] [CrossRef]
- Peterson, U.; Mannock, D.A.; Lewis, R.N.; Pohl, P.; Mcelhaney, R.N.; Pohl, E.E. Origin of Membrane Dipole Potential: Contribution of The Phospholipid Fatty Acid Chains. Chem. Phys. Lipids 2002, 117, 19–27. [Google Scholar] [CrossRef]
- Hannesschlaeger, C.; Horner, A.; Pohl, P. Intrinsic Membrane Permeability to Small Molecules. Chem. Rev. 2019, 119, 5922–5953. [Google Scholar] [CrossRef]
- Chekashkina, K.; Jovanovic, O.; Kuzmin, P.; Pohl, E.; Pavel, B. The Changes of Physical Parameters of Lipid Membrane Caused By Lipid Peroxidation-Derived Aldehydes. Biophys. J. 2017, 112, 520a. [Google Scholar] [CrossRef][Green Version]
- Galimzyanov, T.R.; Kuzmin, P.I.; Pohl, P.; Akimov, S.A. Undulations Drive Domain Registration From The Two Membrane Leaflets. Biophys. J. 2017, 112, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Jovanovic, O.; Chekashkina, K.; Bashkirov, P.; Vazdar, M.; Pohl, E.E. Uncoupling Proteins Are Highly Sensitive to the Membrane Lipid Composition. Eur. Biophys. J. Biophys. Lett. 2017, 46, S287. [Google Scholar]
- Pohl, P.; Rokitskaya, T.I.; Pohl, E.E.; Saparov, S.M. Permeation Of Phloretin Across Bilayer Lipid Membranes Monitored By Dipole Potential And Microelectrode Measurements. Biochim. Biophys. Acta 1997, 1323, 163–172. [Google Scholar] [CrossRef]
- Pohl, E.E.; Krylov, A.V.; Block, M.; Pohl, P. Changes of the Membrane Potential Profile Induced By Verapamil and Propranolol. Biochim. Biophys. Acta 1998, 1373, 170–178. [Google Scholar] [CrossRef]
- Jovanovic, O.; Chekashkina, K.; Bashkirov, P.; Skuljc, S.; Vazdar, M.; Pohl, E.E. Lipid Curvature Modulates Function of Mitochondrial Membrane Proteins. Eur. Biophys. J. Biophys. Lett. 2019, 48, S51. [Google Scholar]
- Aimon, S.; Callan-Jones, A.; Berthaud, A.; Pinot, M.; Toombes, G.E.; Bassereau, P. Membrane Shape Modulates Transmembrane Protein Distribution. Dev. Cell 2014, 28, 212–218. [Google Scholar] [CrossRef]
- Phillips, R.; Ursell, T.; Wiggins, P.; Sens, P. Emerging Roles for Lipids in Shaping Membrane-Protein Function. Nature 2009, 459, 379–385. [Google Scholar] [CrossRef]
- Farooqui, A.A.; Horrocks, L.A.; Farooqui, T. Modulation of Inflammation in Brain: A Matter of Fat. J. Neurochem. 2007, 101, 577–599. [Google Scholar] [CrossRef]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pohl, E.E.; Jovanovic, O. The Role of Phosphatidylethanolamine Adducts in Modification of the Activity of Membrane Proteins under Oxidative Stress. Molecules 2019, 24, 4545. https://doi.org/10.3390/molecules24244545
Pohl EE, Jovanovic O. The Role of Phosphatidylethanolamine Adducts in Modification of the Activity of Membrane Proteins under Oxidative Stress. Molecules. 2019; 24(24):4545. https://doi.org/10.3390/molecules24244545
Chicago/Turabian StylePohl, Elena E., and Olga Jovanovic. 2019. "The Role of Phosphatidylethanolamine Adducts in Modification of the Activity of Membrane Proteins under Oxidative Stress" Molecules 24, no. 24: 4545. https://doi.org/10.3390/molecules24244545
APA StylePohl, E. E., & Jovanovic, O. (2019). The Role of Phosphatidylethanolamine Adducts in Modification of the Activity of Membrane Proteins under Oxidative Stress. Molecules, 24(24), 4545. https://doi.org/10.3390/molecules24244545