
Isorhamnetin Has Potential for the Treatment of Escherichia coli-Induced Sepsis



Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
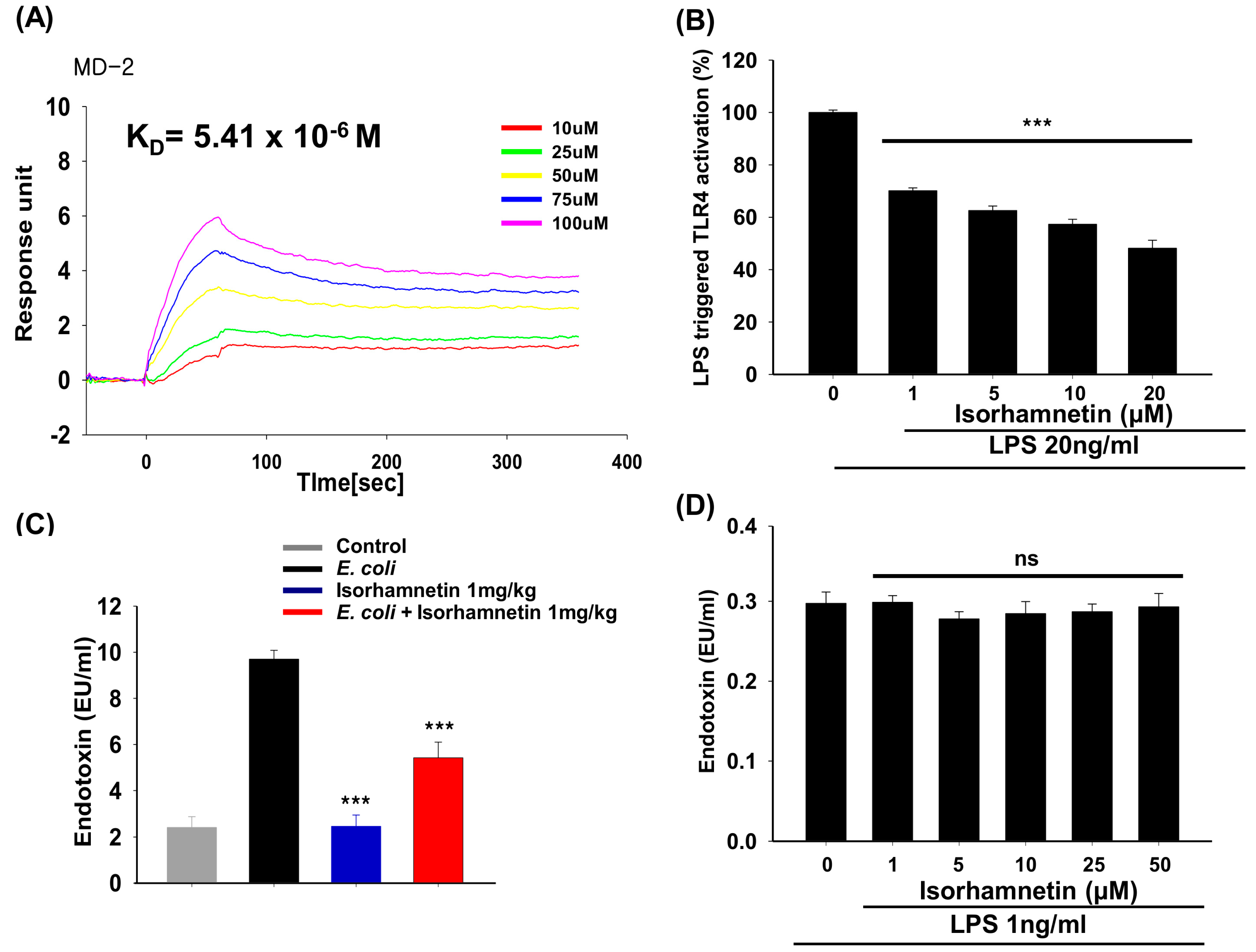{kind=link}
{kind=link}
1. Introduction
2. Results
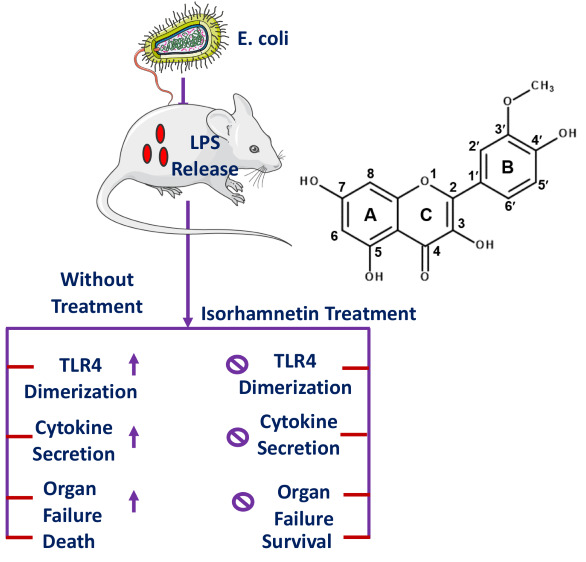
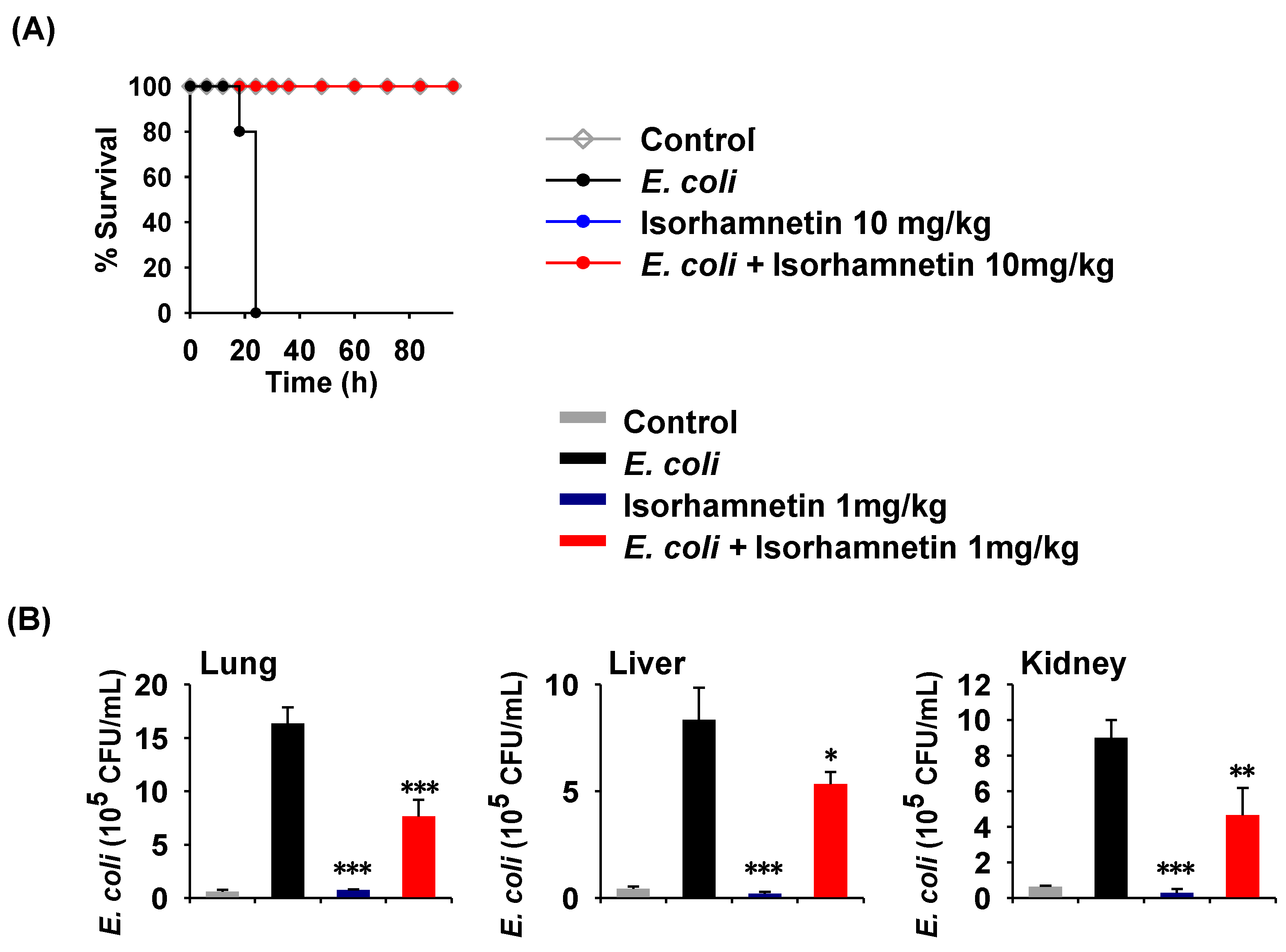
2.1. Isorhamnetin Treatment Protects Mice from E. coli-Induced Sepsis
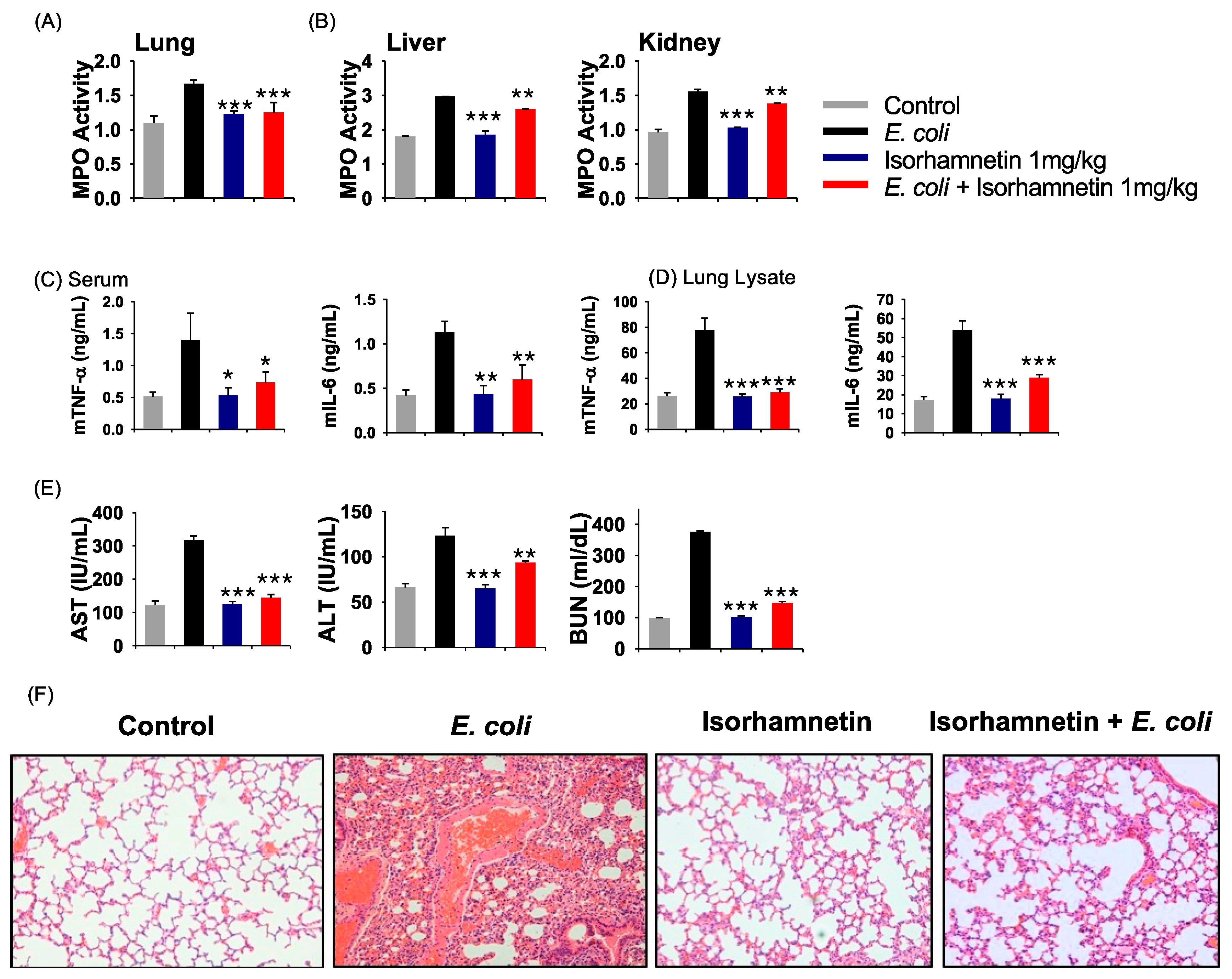
2.2. Isorhamnetin Treatment Protects Mice from E. coli-Induced Inflammation
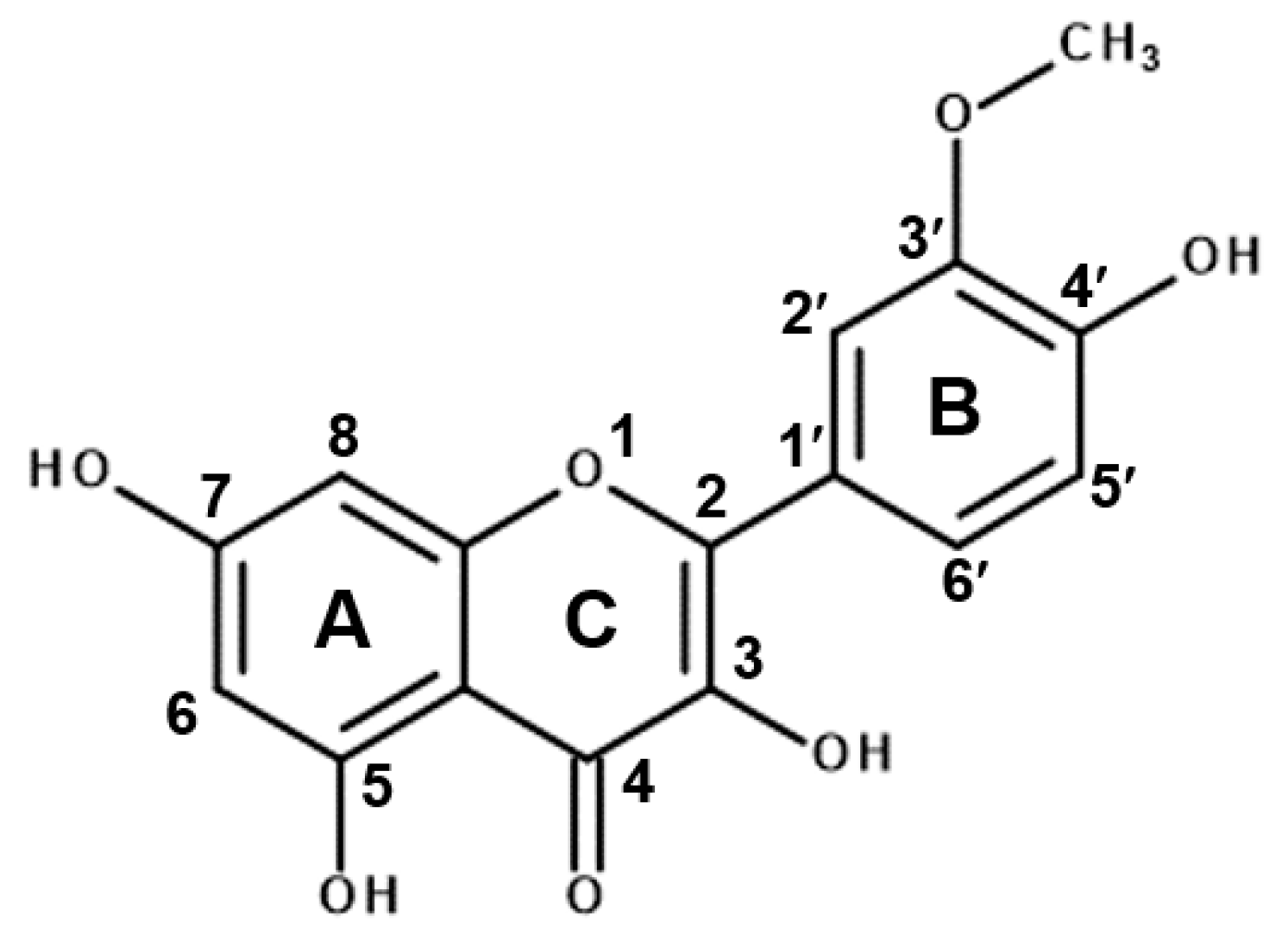
2.3. Interactions between Isorhamnetin and MD-2 Represent the Principal Mechanism Underlying the Anti-Inflammatory Activity
2.4. Molecular Docking
3. Discussion
4. Materials and Methods
4.1. Chemicals and Biological Reagents
4.2. Animals
4.3. SEAP Assay
4.4. Measurement of Binding Affinity by SPR
4.5. Molecular Docking
4.6. Survival test for the sepsis mouse model
4.7. Cytokine Levels in the Serum and Lung Lysates in the Sepsis Mouse Model
4.8. Detection of AST, ALT, and BUN in the mouse serum
4.9. Determination of E. coli Counts in Organ Tissues
4.10. LAL Assay
4.11. Histopathological Examination
4.12. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Tiru, B.; DiNino, E.K.; Orenstein, A.; Mailloux, P.T.; Pesaturo, A.; Gupta, A.; McGee, W.T. The economic and humanistic burden of severe sepsis. PharmacoEconomics 2015, 33, 925–937. [Google Scholar] [CrossRef] [PubMed]
- Fleischmann, C.; Scherag, A.; Adhikari, N.K.; Hartog, C.S.; Tsaganos, T.; Schlattmann, P.; Angus, D.C.; Reinhart, K.; International Forum of Acute Care Trialists. Assessment of global incidence and mortality of hospital-treated sepsis. Current estimates and limitations. Am. J. Respir. Crit. Care Med. 2016, 193, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Hotchkiss, R.S.; Moldawer, L.L.; Opal, S.M.; Reinhart, K.; Turnbull, I.R.; Vincent, J.L. Sepsis and septic shock. Nat. Rev. Dis. Primers 2016, 2, 16045. [Google Scholar] [CrossRef] [PubMed]
- Boyd, J.H.; Russell, J.A.; Fjell, C.D. The meta-genome of sepsis: Host genetics, pathogens and the acute immune response. J. Innate. Immun. 2014, 6, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Ohto, U. Conservation and divergence of ligand recognition and signal transduction mechanisms in toll-like receptors. Chem. Pharm. Bull. 2017, 65, 697–705. [Google Scholar] [CrossRef]
- Bell, J.K.; Mullen, G.E.; Leifer, C.A.; Mazzoni, A.; Davies, D.R.; Segal, D.M. Leucine-rich repeats and pathogen recognition in toll-like receptors. Trends Immunol. 2003, 24, 528–533. [Google Scholar] [CrossRef]
- Lu, Y.C.; Yeh, W.C.; Ohashi, P.S. Lps/tlr4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef]
- O’Neill, L.A.; Bowie, A.G. The family of five: Tir-domain-containing adaptors in toll-like receptor signalling. Nat. Reviews. Immunol. 2007, 7, 353–364. [Google Scholar] [CrossRef]
- Pérez-Cano, F.J.; Massot-Cladera, M.; Rodríguez-Lagunas, M.J.; Castell, M. Flavonoids affect host-microbiota crosstalk through TLR modulation. Antioxidants 2014, 3, 649–670. [Google Scholar] [CrossRef]
- Corradini, E.; Foglia, P.; Giansanti, P.; Gubbiotti, R.; Samperi, R.; Lagana, A. Flavonoids: Chemical properties and analytical methodologies of identification and quantitation in foods and plants. Nat. Prod. Res. 2011, 25, 469–495. [Google Scholar] [CrossRef]
- Monici, M.; Baglioni, P.; Mulinacci, N.; Baldi, A.; Vincieri, F.F. A research model on flavonoids as photoprotectors: Studies on the photochemistry of kaempferol and pelargonidin. Acta Hortic. 1994, 381, 340–347. [Google Scholar] [CrossRef]
- Nijveldt, R.J.; van Nood, E.; van Hoorn, D.E.C.; Boelens, P.G.; van Norren, K.; van Leeuwen, P.A.M. Flavonoids: A review of probable mechanisms of action and potential applications. Am. J. Clin. Nutr. 2001, 74, 418–425. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.N.; Shin, S.A.; Choo, G.S.; Kim, H.J.; Park, Y.S.; Kim, B.S.; Kim, S.K.; Cho, S.D.; Nam, J.S.; Choi, C.S.; et al. Antiinflammatory effect of quercetin and galangin in lpsstimulated raw264.7 macrophages and dncbinduced atopic dermatitis animal models. Int. J. Mol. Med. 2018, 41, 888–898. [Google Scholar] [PubMed]
- Cho, Y.H.; Kim, N.H.; Khan, I.; Yu, J.M.; Jung, H.G.; Kim, H.H.; Jang, J.Y.; Kim, H.J.; Kim, D.I.; Kwak, J.H.; et al. Anti-inflammatory potential of quercetin-3-o-beta-d-(“2”-galloyl)-glucopyranoside and quercetin isolated from diospyros kaki calyx via suppression of map signaling molecules in lps-induced raw 264.7 macrophages. J. Food Sci. 2016, 81, C2447–C2456. [Google Scholar] [CrossRef]
- Qi, F.; Sun, J.H.; Yan, J.Q.; Li, C.M.; Lv, X.C. Anti-inflammatory effects of isorhamnetin on lps-stimulated human gingival fibroblasts by activating nrf2 signaling pathway. Microb. Pathog 2018, 120, 37–41. [Google Scholar] [CrossRef]
- Jaramillo, S.; Lopez, S.; Varela, L.M.; Rodriguez-Arcos, R.; Jimenez, A.; Abia, R.; Guillen, R.; Muriana, F.J.G. The flavonol isorhamnetin exhibits cytotoxic effects on human colon cancer cells. J. Agr. Food Chem. 2010, 58, 10869–10875. [Google Scholar] [CrossRef]
- Choi, Y.H. The cytoprotective effect of isorhamnetin against oxidative stress is mediated by the upregulation of the nrf2-dependent ho-1 expression in c2c12 myoblasts through scavenging reactive oxygen species and erk inactivation. Gen. Physiol. Biophys 2016, 35, 145–154. [Google Scholar] [CrossRef]
- Jnawali, H.N.; Jeon, D.; Jeong, M.C.; Lee, E.; Jin, B.; Ryoo, S.; Yoo, J.; Jung, I.D.; Lee, S.J.; Park, Y.M.; et al. Antituberculosis activity of a naturally occurring flavonoid, isorhamnetin. J. Nat. Prod. 2016, 79, 961–969. [Google Scholar] [CrossRef]
- Antunes-Ricardo, M.; Gutierrez-Uribe, J.A.; Martinez-Vitela, C.; Serna-Saldivar, S.O. Topical anti-inflammatory effects of isorhamnetin glycosides isolated from opuntia ficus-indica. Biomed. Res. Int. 2015, 2015, 847320. [Google Scholar] [CrossRef]
- Akbay, P.; Basaran, A.A.; Undeger, U.; Basaran, N. In vitro immunomodulatory activity of flavonoid glycosides from Urtica dioica L. Phytother. Res. 2003, 17, 34–37. [Google Scholar] [CrossRef]
- Kim, S.Y.; Jin, C.-Y.; Kim, C.H.; Yoo, Y.H.; Choi, S.H.; Kim, G.-Y.; Yoon, H.M.; Park, H.T.; Choi, Y.H. Isorhamnetin alleviates lipopolysaccharide-induced inflammatory responses in bv2 microglia by inactivating nf-κb, blocking the tlr4 pathway and reducing ros generation. Int. J. Mol. Med. 2019, 43, 682–692. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shan, X.O.; Chen, G.Z.; Jiang, L.L.; Wang, Z.; Fang, Q.L.; Liu, X.; Wang, J.Y.; Zhang, Y.L.; Wu, W.C.; et al. Md-2 as the target of a novel small molecule, l6h21, in the attenuation of lps-induced inflammatory response and sepsis. Br. J. Pharmacol. 2015, 172, 4391–4405. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.T.; Chen, L.F.; Wang, Z.; Zhao, C.W.; Chen, G.Z.; Liu, X.; Dai, Y.R.; Cai, Y.P.; Li, C.L.; Zhou, J.M.; et al. Determination of the binding mode for anti-inflammatory natural product xanthohumol with myeloid differentiation protein 2. Drug Des. Dev. 2016, 10, 455–463. [Google Scholar]
- Wang, Z.; Chen, G.Z.; Chen, L.F.; Liu, X.; Fu, W.T.; Zhang, Y.L.; Li, C.L.; Liang, G.; Cai, Y.P. Insights into the binding mode of curcumin to md-2: Studies from molecular docking, molecular dynamics simulations and experimental assessments. Mol. Biosyst 2015, 11, 1933–1938. [Google Scholar] [CrossRef]
- Chen, G.; Zhang, Y.; Liu, X.; Fang, Q.; Wang, Z.; Fu, L.; Liu, Z.; Wang, Y.; Zhao, Y.; Li, X.; et al. Discovery of a new inhibitor of myeloid differentiation 2 from cinnamamide derivatives with anti-inflammatory activity in sepsis and acute lung injury. J. Med. Chem. 2016, 59, 2436–2451. [Google Scholar] [CrossRef]
- Wang, Y.; Shan, X.; Dai, Y.; Jiang, L.; Chen, G.; Zhang, Y.; Wang, Z.; Dong, L.; Wu, J.; Guo, G.; et al. Curcumin analog l48h37 prevents lipopolysaccharide-induced tlr4 signaling pathway activation and sepsis via targeting md2. J. Pharmacol. Exp. Ther. 2015, 353, 539–550. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, J.; Ying, S.; Chen, G.; Wu, B.; Xu, T.; Liu, Z.; Liu, X.; Huang, L.; Shan, X.; et al. Discovery of new md2 inhibitor from chalcone derivatives with anti-inflammatory effects in lps-induced acute lung injury. Sci. Rep. 2016, 6, 25130. [Google Scholar] [CrossRef]
- Ramachandran, L.; Manu, K.A.; Shanmugam, M.K.; Li, F.; Siveen, K.S.; Vali, S.; Kapoor, S.; Abbasi, T.; Surana, R.; Smoot, D.T.; et al. Isorhamnetin inhibits proliferation and invasion and induces apoptosis through the modulation of peroxisome proliferator-activated receptor gamma activation pathway in gastric cancer. J. Biol. Chem. 2012, 287, 38028–38040. [Google Scholar] [CrossRef]
- Pengfei, L.; Tiansheng, D.; Xianglin, H.; Jianguo, W. Antioxidant properties of isolated isorhamnetin from the sea buckthorn marc. Plant. Foods Hum. Nutr 2009, 64, 141–145. [Google Scholar] [CrossRef]
- Yang, B.; Li, X.P.; Ni, Y.F.; Du, H.Y.; Wang, R.; Li, M.J.; Wang, W.C.; Li, M.M.; Wang, X.H.; Li, L.; et al. Protective effect of isorhamnetin on lipopolysaccharide-induced acute lung injury in mice. Inflammation 2016, 39, 129–137. [Google Scholar] [CrossRef]
- Schneider, M. Complementary and alternative medicines during cancer treatments in the united states. Oncologie 2007, 9, 64–65. [Google Scholar] [CrossRef]
- Karak, P. Biological activities of flavonoids: An overview. Int. J. Pharm Sci Res. 2019, 10, 1567–1574. [Google Scholar]
- Li, Y.; Yao, J.Y.; Han, C.Y.; Yang, J.X.; Chaudhry, M.T.; Wang, S.N.; Liu, H.N.; Yin, Y.L. Quercetin, inflammation and immunity. Nutrients 2016, 8, 167. [Google Scholar] [CrossRef] [PubMed]
- Jnawali, H.N.; Lee, E.; Jeong, K.W.; Shin, A.; Heo, Y.S.; Kim, Y. Anti-inflammatory activity of rhamnetin and a model of its binding to c-jun nh2-terminal kinase 1 and p38 mapk. J. Nat. Prod. 2014, 77, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Lee, E.J.; Cho, J.; Gharbi, A.; Han, H.D.; Kang, T.H.; Kim, Y.; Lee, Y.; Park, W.S.; Jung, I.D.; et al. Tamarixetin exhibits anti-inflammatory activity and prevents bacterial sepsis by increasing il-10 production. J. Nat. Prod. 2018, 81, 1435–1443. [Google Scholar] [CrossRef] [PubMed]
- Peri, F.; Calabrese, V. Toll-like receptor 4 (tlr4) modulation by synthetic and natural compounds: An update. J. Med. Chem. 2014, 57, 3612–3622. [Google Scholar] [CrossRef]
- Jayaprakasha, G.K.; Jagan, L.; Rao, M.; Sakariah, K.K. Chemistry and biological activities of c-longa. Trends Food Sci. Tech. 2005, 16, 533–548. [Google Scholar] [CrossRef]
- Choi, Y.H.; Kim, G.Y.; Lee, H.H. Anti-inflammatory effects of cordycepin in lipopolysaccharide-stimulated raw 264.7 macrophages through toll-like receptor 4-mediated suppression of mitogen-activated protein kinases and nf-kappa b signaling pathways. Drug Des. Dev. 2014, 8, 1941–1953. [Google Scholar] [CrossRef] [PubMed]
- Gradisar, H.; Keber, M.M.; Pristovsek, P.; Jerala, R. Md-2 as the target of curcumin in the inhibition of response to lps. J. Leukoc. Biol. 2007, 82, 968–974. [Google Scholar] [CrossRef]
- Kang, J.Y.; Lee, J.O. Structural biology of the toll-like receptor family. Annu. Rev. Biochem. 2011, 80, 917–941. [Google Scholar] [CrossRef]
- Molteni, M.; Bosi, A.; Rossetti, C. Natural products with toll-like receptor 4 antagonist activity. Int. J. Inflamm 2018, 2859135. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Durai, P.; Jeon, D.; Jung, I.D.; Lee, S.J.; Park, Y.M.; Kim, Y. Phloretin as a potent natural TLR2/1 inhibitor suppresses tlr2-induced inflammation. Nutrients 2018, 10, 868. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Jacob, B.; Jang, M.; Kwak, C.; Lee, Y.; Son, K.; Lee, S.; Jung, I.D.; Jeong, M.S.; Kwon, S.H.; et al. Development of a novel short 12-meric papiliocin-derived peptide that is effective against gram-negative sepsis. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Isorhamnetin can be purchased from Sigma-Aldrich (St. Louis, MO, USA). |





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chauhan, A.K.; Kim, J.; Lee, Y.; Balasubramanian, P.K.; Kim, Y. Isorhamnetin Has Potential for the Treatment of Escherichia coli-Induced Sepsis. Molecules 2019, 24, 3984. https://doi.org/10.3390/molecules24213984
Chauhan AK, Kim J, Lee Y, Balasubramanian PK, Kim Y. Isorhamnetin Has Potential for the Treatment of Escherichia coli-Induced Sepsis. Molecules. 2019; 24(21):3984. https://doi.org/10.3390/molecules24213984
Chicago/Turabian StyleChauhan, Anil Kumar, Jieun Kim, Yeongjoon Lee, Pavithra K. Balasubramanian, and Yangmee Kim. 2019. "Isorhamnetin Has Potential for the Treatment of Escherichia coli-Induced Sepsis" Molecules 24, no. 21: 3984. https://doi.org/10.3390/molecules24213984
APA StyleChauhan, A. K., Kim, J., Lee, Y., Balasubramanian, P. K., & Kim, Y. (2019). Isorhamnetin Has Potential for the Treatment of Escherichia coli-Induced Sepsis. Molecules, 24(21), 3984. https://doi.org/10.3390/molecules24213984