(3β,16α)-3,16-Dihydroxypregn-5-en-20-one from the Twigs of Euonymus alatus (Thunb.) Sieb. Exerts Anti-Inflammatory Effects in LPS-Stimulated RAW-264.7 Macrophages




Abstract
1. Introduction
2. Results and Discussion
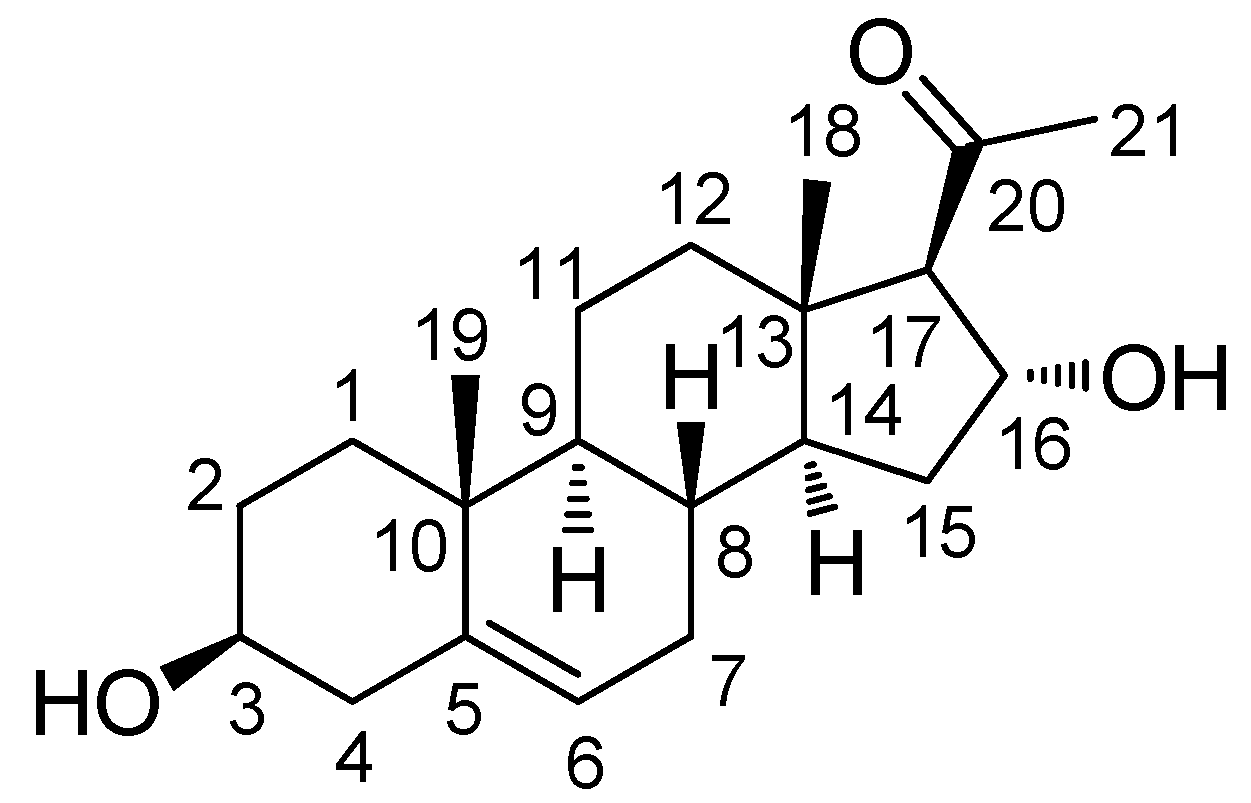
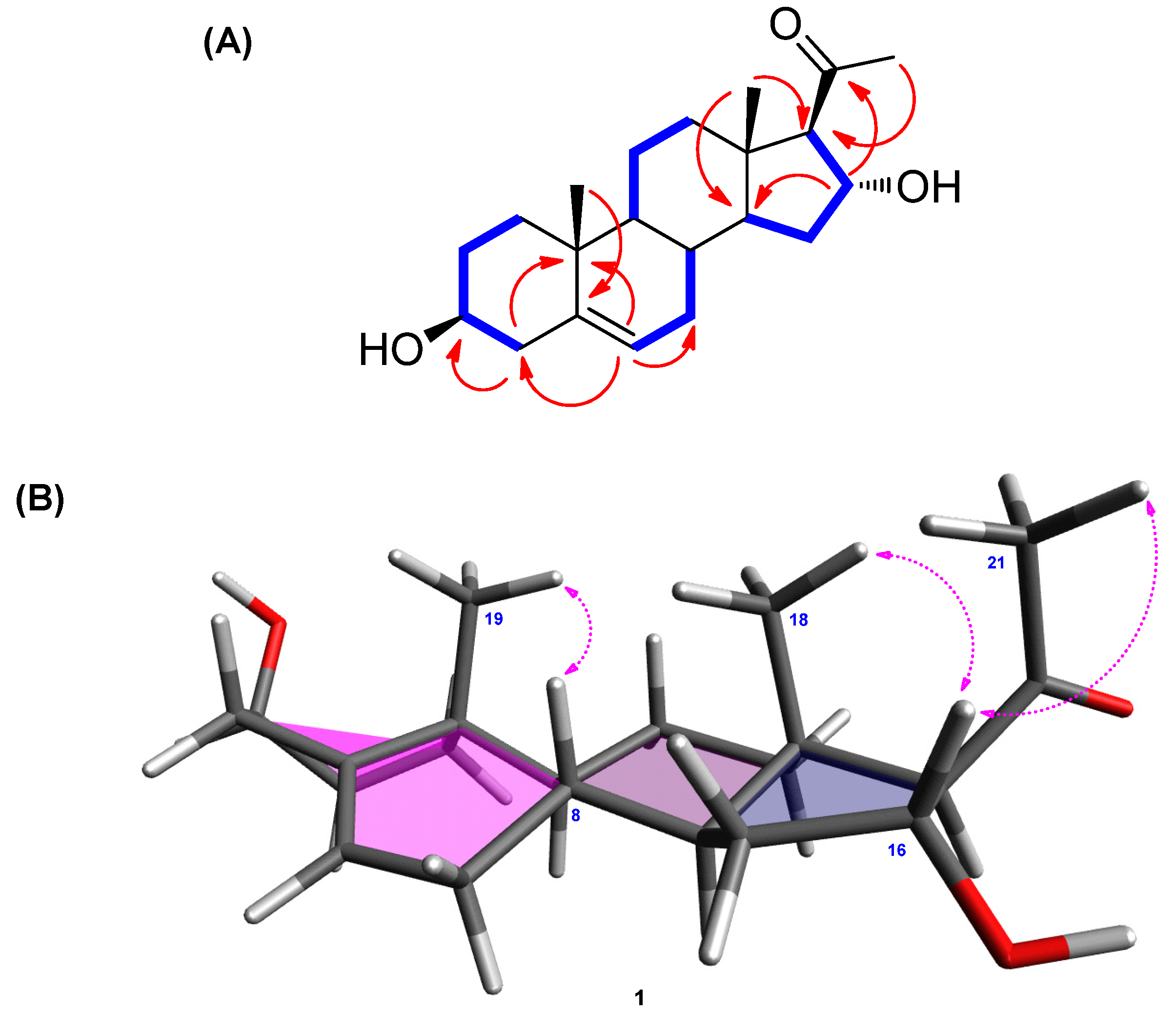
2.1. Isolation and Identification of (3β,16α)-3,16-Dihydroxypregn-5-en-20-one
2.2. Effect of Compound 1 on NO Production
2.3. Compound 1 Downregulates MAPKs (JNK, ERK, and p38) in LPS-Stimulated RAW 264.7 Mouse Macrophages
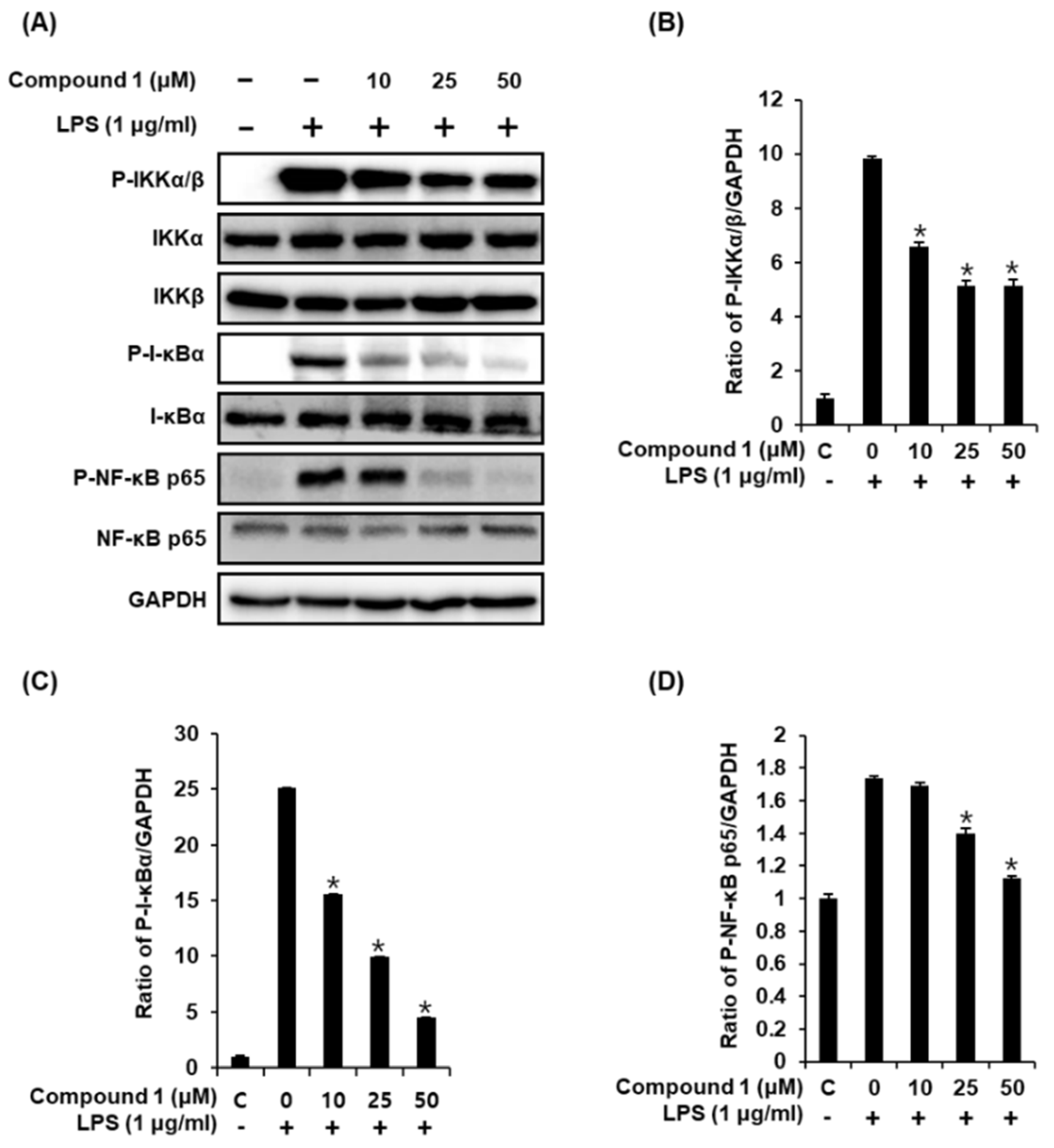
2.4. Compound 1 Downregulates IKKα/β, I-κBα, and NF-κB p65 in LPS-Stimulated RAW 264.7 Mouse Macrophages
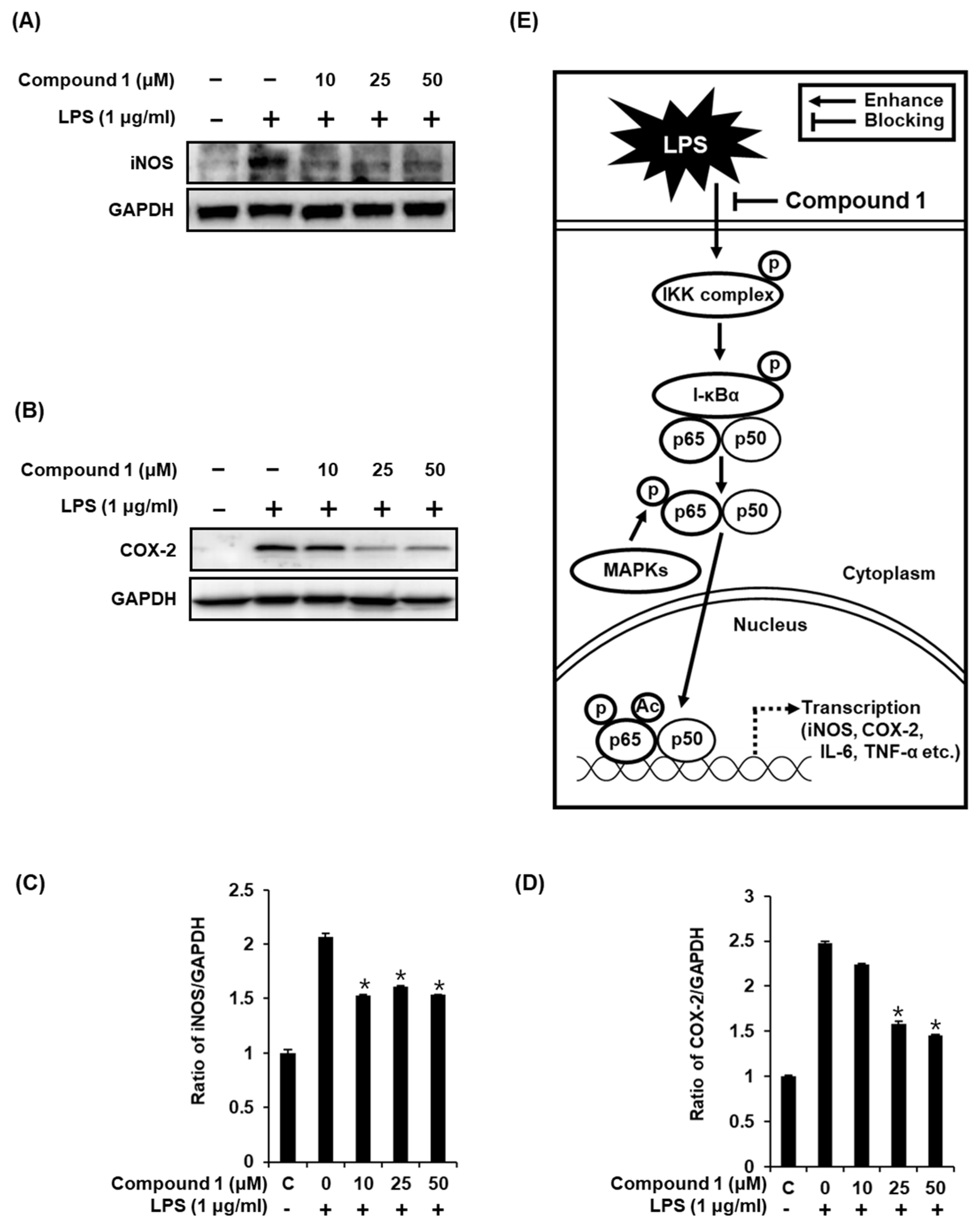
2.5. Compound 1 Downregulates iNOS and COX-2 Expression in LPS-Stimulated RAW 264.7 Mouse Macrophages
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Plant Material
3.3. Extraction and Isolation
(3β,16α)-3,16-Dihydroxypregn-5-en-20-one (1):
3.4. Cell Culture
3.5. Measurement of RAW 264.7 Cell Viability
3.6. Measurement of NO Production in RAW 264.7 Cells
3.7. Western Blot Analysis
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cario, E.; Rosenberg, I.M.; Brandwein, S.L.; Beck, P.L.; Reinecker, H.-C.; Podolsky, D.K. Lipopolysaccharide activates distinct signaling pathways in intestinal epithelial cell lines expressing Toll-like receptors. J. Immunol. 2000, 164, 966–972. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.W.; Lee, K.S.; Kim, C.W. Curcumin attenuates the expression of IL-1ß, IL-6, and TNF-α as well as cyclin e in TNF-α-treated hacat cells; NF-κB and MAPKs as potential upstream targets. Int. J. Mol. Med. 2007, 19, 469–474. [Google Scholar] [PubMed]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed]
- Sharif, O.; Bolshakov, V.N.; Raines, S.; Newham, P.; Perkins, N.D. Transcriptional profiling of the LPS induced NF-κB response in macrophages. BMC Immunol. 2007, 8, 1. [Google Scholar] [CrossRef]
- Förstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart J. 2011, 33, 829–837. [Google Scholar] [CrossRef]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef]
- Schoeb, T.R.; Jarmi, T.; Hicks, M.J.; Henke, S.; Zarjou, A.; Suzuki, H.; Kramer, P.; Novak, J.; Agarwal, A.; Bullard, D.C. Endothelial nitric oxide synthase inhibits the development of autoimmune-mediated vasculitis in mice. Arthritis Rheum. 2012, 64, 4114–4124. [Google Scholar] [CrossRef]
- Chun, J.; Song, K.; Kim, Y.S. Anti-inflammatory activity of standardized fraction from Inula helenium L. via Suppression of NF-κB pathway in RAW 264.7 cells. Nat. Prod. Sci. 2019, 25, 16–22. [Google Scholar] [CrossRef][Green Version]
- Roy, A.; Park, H.-J.; Jung, H.A.; Choi, J.S. Estragole exhibits anti-inflammatory activity with the regulation of NF-κB and Nrf-2 signaling pathways in LPS-induced RAW 264.7 cells. Nat. Prod. Sci. 2018, 24, 13–20. [Google Scholar] [CrossRef]
- Lee, S.; Lee, D.; Lee, J.C.; Kang, K.S.; Ryoo, R.; Park, H.-J.; Kim, K.H. Bioactivity-guided isolation of anti-inflammatory constituents of the rare mushroom Calvatia nipponica in LPS-stimulated RAW264.7 macrophages. Chem. Biodivers. 2018, 15, e1800203. [Google Scholar] [CrossRef]
- Lee, S.; Lee, D.; Park, J.Y.; Seok, S.; Jang, T.S.; Park, H.B.; Shim, S.H.; Kang, K.S.; Kim, K.H. Anti-gastritis effects of Armillariella tabescens (Scop.) Sing. and the identification of its anti-inflammatory metabolites. J. Pharm. Pharmacol. 2018, 70, 404–412. [Google Scholar] [CrossRef] [PubMed]
- Pack, J.H.; Sung, S.H. Medicinal Plants; Shinil Books Co.: Seoul, Korea, 2007; pp. 446–447. [Google Scholar]
- Jeong, S.-Y.; Nguyen, P.-H.; Zhao, B.-T.; Ali, M.Y.; Choi, J.-S.; Min, B.-S.; Woo, M.-H. Chemical constituents of Euonymus alatus (Thunb.) Sieb. and their PTP1B and α-glucosidase inhibitory activities. Phytother. Res. 2015, 29, 1540–1548. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.-H.; Yang, M.H.; Heo, J.-D.; Sung, S.H.; Jeong, E.J. Dihydrobenzofuran neolignans isolated from Euonymus alatus leaves and twigs attenuated inflammatory responses in the activated RAW264.7 macrophage cells. Nat. Prod. Sci. 2016, 22, 53–59. [Google Scholar] [CrossRef]
- Yan, Z.-H.; Han, Z.-Z.; Hu, X.-Q.; Liu, Q.-X.; Zhang, W.-D.; Liu, R.-H.; Li, H.-L. Two new sesquiterpenes from Euonymus alatus. Helv. Chim. Acta 2013, 96, 85–92. [Google Scholar] [CrossRef]
- Lee, S.; Moon, E.; Choi, S.U.; Kim, K.H. Lignans from the twigs of Euonymus alatus (Thunb.) Siebold and their biological evaluation. Chem. Biodivers. 2016, 13, 1391–1396. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.-I.; Lee, S.R.; Kim, K.H. Antioxidant and α-glucosidase inhibitory activities of constituents from Euonymus alatus twigs. Ind. Crops Prod. 2015, 76, 1055–1060. [Google Scholar] [CrossRef]
- Jeong, E.J.; Bae, J.-Y.; Rho, J.-R.; Kim, Y.C.; Ahn, M.-J.; Sung, S.H. Anti-inflammatory triterpenes from Euonymus alatus leaves and twigs on lipopolysaccharide-activated RAW264.7 macrophage cells. Bull. Korean Chem. Soc. 2014, 35, 2945–2949. [Google Scholar] [CrossRef]
- Lee, S.R.; Park, Y.J.; Han, Y.B.; Lee, J.C.; Lee, S.; Park, H.-J.; Lee, H.-J.; Kim, K.H. Isoamericanoic acid B from Acer tegmentosum as a potential phytoestrogen. Nutrients 2018, 10, 1915. [Google Scholar] [CrossRef]
- Lee, S.R.; Choi, E.; Jeon, S.H.; Zhi, X.Y.; Yu, J.S.; Kim, S.-H.; Lee, J.; Park, K.-M.; Kim, K.H. Tirucallane triterpenoids from the stems and stem bark of Cornus walteri that control adipocyte and osteoblast differentiations. Molecules 2018, 23, 2732. [Google Scholar] [CrossRef]
- Yu, J.S.; Roh, H.S.; Baek, K.H.; Lee, S.; Kim, S.; So, H.M.; Moon, E.; Pang, C.; Jang, T.S.; Kim, K.H. Bioactivity-guided isolation of ginsenosides from Korean Red Ginseng with cytotoxic activity against human lung adenocarcinoma cells. J. Ginseng Res. 2018, 42, 562–570. [Google Scholar] [CrossRef]
- Baek, S.C.; Choi, E.; Eom, H.J.; Jo, M.S.; Kim, S.; So, H.M.; Kim, S.H.; Kang, K.S.; Kim, K.H. LC/MS-based analysis of bioactive compounds from the bark of Betula platyphylla var. japonica and their effects on regulation of adipocyte and osteoblast differentiation. Nat. Prod. Sci. 2018, 24, 235–240. [Google Scholar] [CrossRef]
- So, H.M.; Eom, H.J.; Lee, D.; Kim, S.; Kang, K.S.; Lee, I.K.; Baek, K.H.; Park, J.Y.; Kim, K.H. Bioactivity evaluations of betulin identified from the bark of Betula platyphylla var. japonica for cancer therapy. Arch. Pharm. Res. 2018, 41, 815–822. [Google Scholar] [PubMed]
- Kim, K.H.; Ha, S.K.; Choi, S.U.; Kim, S.Y.; Lee, K.R. Phenolic constituents from the twigs of Euonymus alatus and their cytotoxic and anti-inflammatory activity. Planta Med. 2013, 79, 361–364. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.R.; Eom, H.J.; Lee, S.R.; Choi, S.U.; Kang, K.S.; Lee, K.R.; Kim, K.H. Bioassay-guided isolation of antiproliferative triterpenoids from Euonymus alatus twigs. Nat. Prod. Commun. 2015, 10, 1929–1932. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.E.; Johnson, T.W. First total synthesis of xestobergsterol A and active structural analogues of the xestobergsterols. Tetrahedron 2001, 57, 1449–1481. [Google Scholar] [CrossRef]
- Boonyarattanakalin, S.; Martin, S.E.; Dykstra, S.A.; Peterson, B.R. Synthetic mimics of small mammalian cell surface receptors. J. Am. Chem. Soc. 2004, 126, 16379–16386. [Google Scholar] [CrossRef]
- Shapiro, E.L.; Gentles, M.J. Electrochemical reduction of epoxy ketones. J. Org. Chem. 1981, 46, 5017–5019. [Google Scholar] [CrossRef]
- Salvador, J.A.R.; Leitão, A.J.L.; Melo, M.L.S.; Hanson, J.R. Hydrazine hydrate induced reductive cleavage of α,β-epoxy ketones: An efficient procedure for the preparation of β-hydroxy ketones. Tetrahedron Lett. 2005, 46, 1067–1070. [Google Scholar] [CrossRef]
- Hansen, F.; Kalle, M.; van der Plas, M.; Stromdahl, A.C.; Malmsten, M.; Morgelin, M.; Schmidtchen, A. The thrombin-derived host defense peptide Gky25 inhibits endotoxin-induced responses through interactions with lipopolysaccharide and macrophages/monocytes. Wound Repair Regen. 2015, 23, A11. [Google Scholar] [CrossRef]
- Hambleton, J.; Weinstein, S.L.; Lem, L.; DeFranco, A.L. Activation of c-Jun N-terminal kinase in bacterial lipopolysaccharide-stimulated macrophages. Proc. Natl. Acad. Sci. USA 1996, 93, 2774–2778. [Google Scholar] [CrossRef]
- Park, K.-I.; Kang, S.-R.; Park, H.-S.; Lee, D.H.; Nagappan, A.; Kim, J.A.; Shin, S.C.; Kim, E.H.; Lee, W.S.; Chung, H.-J. Regulation of proinflammatory mediators via NF-κB and p38 MAPK-dependent mechanisms in RAW 264.7 macrophages by polyphenol components isolated from Korea Lonicera japonica Thunb. Evid. Based Complement. Alternat. Med. 2012, 2012, 828521. [Google Scholar] [CrossRef]
- Kaminska, B. MAPK signaling pathways as molecular targets for anti-inflammatory therapy—from molecular mechanisms to therapeutic benefits. Biochim. Biophys. Acta 2005, 1754, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.; Lee, M.-Y. Anti-inflammatory activity of Populus deltoides leaf extract via modulating NF-κB and p38/JNK pathways. Int. J. Mol. Sci. 2018, 19, 3746. [Google Scholar] [CrossRef] [PubMed]
- Caamaño, J.; Hunter, C.A. NF-κB family of transcription factors: Central regulators of innate and adaptive immune functions. Clin. Microbiol. Rev. 2002, 15, 414–429. [Google Scholar] [CrossRef] [PubMed]
- Nahrevanian, H. Immune effector mechanisms of the nitric oxide pathway in malaria: Cytotoxicity versus cytoprotection. Braz. J. Infect. Dis. 2006, 10, 283–292. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | 1 | 1 [29] | ||
|---|---|---|---|---|
| δHa | δCa | δH | δC | |
| 1 | 1.11 dd (13.5, 3.5), 1.88 dt (13.5, 3.5) | 37.0 | ||
| 2 | 1.48 m, 1.80 m | 30.8 | ||
| 3 | 3.40 m | 70.9 | 3.48 m | 73.7 |
| 4 | 2.24 m | 41.5 | ||
| 5 | 140.9 | 141.1 | ||
| 6 | 5.35 m | 120.7 | 5.34 d (4.0) | 121.3 |
| 7 | 1.64 m, 1.96 m | 31.3 | ||
| 8 | 2.20 m | 31.3 | ||
| 9 | 1.07 m | 50.1 | ||
| 10 | 36.3 | |||
| 11 | 1.50 m, 1.69 m | 20.4 | ||
| 12 | 1.57 m b, 2.01 m | 38.5 | ||
| 13 | 44.7 | |||
| 14 | 1.56 m b | 54.3 | ||
| 15 | 1.54 m b, 1.72 d (8.5) | 35.1 | ||
| 16 | 4.72 m | 71.4 | 4.77 m | 71.3 |
| 17 | 2.55 d (6.5) | 73.2 | 2.57 d (6.5) | 71.7 |
| 18 | 0.64 s | 13.3 | 0.64 s | |
| 19 | 1.02 s | 18.4 | 1.01 s | |
| 20 | 209.3 | 210.2 | ||
| 21 | 2.16 s | 30.7 | 2.19 s | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.; Lee, D.; Baek, S.C.; Jo, M.S.; Kang, K.S.; Kim, K.H. (3β,16α)-3,16-Dihydroxypregn-5-en-20-one from the Twigs of Euonymus alatus (Thunb.) Sieb. Exerts Anti-Inflammatory Effects in LPS-Stimulated RAW-264.7 Macrophages. Molecules 2019, 24, 3848. https://doi.org/10.3390/molecules24213848
Lee S, Lee D, Baek SC, Jo MS, Kang KS, Kim KH. (3β,16α)-3,16-Dihydroxypregn-5-en-20-one from the Twigs of Euonymus alatus (Thunb.) Sieb. Exerts Anti-Inflammatory Effects in LPS-Stimulated RAW-264.7 Macrophages. Molecules. 2019; 24(21):3848. https://doi.org/10.3390/molecules24213848
Chicago/Turabian StyleLee, Seulah, Dahae Lee, Su Cheol Baek, Mun Seok Jo, Ki Sung Kang, and Ki Hyun Kim. 2019. "(3β,16α)-3,16-Dihydroxypregn-5-en-20-one from the Twigs of Euonymus alatus (Thunb.) Sieb. Exerts Anti-Inflammatory Effects in LPS-Stimulated RAW-264.7 Macrophages" Molecules 24, no. 21: 3848. https://doi.org/10.3390/molecules24213848
APA StyleLee, S., Lee, D., Baek, S. C., Jo, M. S., Kang, K. S., & Kim, K. H. (2019). (3β,16α)-3,16-Dihydroxypregn-5-en-20-one from the Twigs of Euonymus alatus (Thunb.) Sieb. Exerts Anti-Inflammatory Effects in LPS-Stimulated RAW-264.7 Macrophages. Molecules, 24(21), 3848. https://doi.org/10.3390/molecules24213848